Происхождение мозга Савельев Сергей Вячеславович
§ 37. Ассоциативный центр мозга рептилий
Рассмотрев общий план строения нервной системы, следует отдельно остановиться на новых принципах организации и работы мозга, впервые реализованных у рептилий. Нервная система архаичных амниот стала логическим развитием строения удачной амфибийной конструкции. Однако мозг амфибий практически выполнял функцию сложного рефлекторного аппарата, а его интеллектуальные возможности остались невостребованными. Эволюция амфибий решалась мышцами, зубами, линейными размерами и масштабами размножения.
Из всех млекопитающих у человека самый развитый и развитый мозг. Однако у всех позвоночных мозг состоит из 3 основных структурных единиц: полушарий головного мозга, мозгового ствола и мозжечка. Сетчатое образование является основной областью обмена информацией, являющейся источником мозговой энергии. В этой сети нервных волокон каждый нейрон может быть связан с другими сшитыми форматами, постоянно получающими сообщения от органов чувств, испускающими, в свою очередь, электрическую активность через кору головного мозга.
Он поддерживает сознание. Если сшитая полоса уменьшает их активность, человек может засыпать или страдать от галлюцинаций, видеть, слышать и иметь нереальные ощущения. Результатом сенсорной депривации, когда человек заперт в темной, тихой и пустой комнате в течение нескольких часов, является кажущееся настроение, транс или кома.
Шло элементарное освоение пищевых ресурсов, где для развития сложного поведения не было ни места, ни биологической необходимости. Со следами этого периода эволюции позвоночных мы сталкиваемся, пытаясь выработать условные рефлексы у различных представителей современных амфибий. Крайне низкая обучаемость и отсутствие долговременной памяти для накопления индивидуального опыта показывают, что сложных поведенческих задач перед древними амфибиями никогда в их эволюции не стояло.
Особенности развития органов чувств и признаки сложного поведения рептилий базируются на особенностях структурной организации головного мозга. Мозг рептилий отличается от мозга амфибий как в количественном, так и в качественном отношении. До появления амниот стратегии поведения или реакция на конкретный раздражитель выбирались по принципу доминантности (см. рис. III-6, е). Этот принцип состоит в том, что выраженного крупного ассоциативного центра мозга у многих первичноводных позвоночных или амфибий нет (см. рис. III-6, e). Выбор формы поведения происходит на основании сравнения активностей примерно равноценных отделов мозга, обслуживающих различные органы чувств. Решающее значение играет уровень возбуждения мозговых аналитических центров одного из анализаторов. Представительство органа чувств, достигшее в мозге наибольшего возбуждения, и становится основной областью для принятия решения. После выбора одной из инстинктивных реакций происходит её поведенческая реализация. Этот процесс осуществляется под контролем того же простого сравнения доминантностей. Если в процессе осуществления реакции возникает новое раздражение, которое изменяет соотношение возбуждений органов чувств, то поведенческая реализация инстинктивного процесса останавливается. Каждая конкретная ситуация отличается от предыдущей, но задействуется тот же набор органов чувств. Если наибольшее возбуждение достигается в той же сенсорной системе, то поведение сохраняется, а если в другой, то изменяется. Поскольку абсолютно идентичные условия в естественной жизни практически не встречаются, поведение даже самых примитивных анамний будет бесконечно разнообразно. Следовательно, поведение каждой особи будет индивидуальным с довольно высокой динамической адаптивностью.
Первые признаки ассоциативного центра появились ещё в мозге амфибий. У них таким центром мог стать средний или промежуточный мозг. Для этого были все основания. В промежуточном мозге находятся нейроэндокринные центры, контролирующие половое поведение, миграции и энергетический баланс организма анамний. Через активизацию центров промежуточного мозга запускаются инстинктивные программы поведения, которые контролируют работу других отделов мозга. Казалось бы, промежуточный мозг мог стать аналитическим центром поведения анамний, а затем и амниот. Однако в этом случае система реализации поведенческих реакций состояла бы не только в работе нервной системы. Каждый раз любое поведенческое событие приводило бы к стимуляции работы нейроэндокринных центров. Гормональная регуляция поведения реализуется долго, а неврологическая - быстро. При быстрой смене форм поведения наступал бы конфликт между инертными гормональными и динамичными нейральными программами поведения. У насекомых этот конфликт был решён в пользу нейрогормональных центров и чисто инстинктивного поведения.
С первичноводными позвоночными и амфибиями сложилась довольно нестабильная ситуация. С одной стороны, роль гормонально-инстинктивной регуляции поведения у амфибий очень велика и явно доминирует при выборе стратегий поведения. С другой стороны, нейроморфологический субстрат развит вполне достаточно для небольшой индивидуализации поведения при реализации этих стратегий. Возникла оригинальная система гормонально-доминантного выбора форм поведения из стандартного инстинктивного набора. У амфибий стратегия поведения определяется нейрогормональным состоянием особи. При реализации выбранной формы поведения она адаптируется к конкретным условиям при помощи сравнения доминантностей, которое было описано выше. В такой схеме контроля поведения анамний не остаётся места для ассоциативного центра. Он мог бы понадобиться только тогда, когда возникла бы потребность в быстрой адаптивной индивидуализации поведения. Такая ситуация может сложиться только в том случае, когда последовательная реализация инстинктивных форм поведения будет прямо зависеть от постоянно меняющейся ситуации.
В таких нестабильных условиях окружающей среды оказались архаичные рептилии. По-видимому, требования к быстрой индивидуализации поведения и памяти возросли, а реализация гормональнодоминантного принципа выбора из стандартного инстинктивного набора форм поведения стала неэффективной. Возник совершенно новый тип принятия решений, который сохранился в мозге современных рептилий, принадлежащих к отдалённым систематическим группам. Их всех объединяет одно принципиально новое качество мозга - выраженный ассоциативный центр (см. рис III-7).
Основной ассоциативный центр рептилий сформировался в крыше среднего мозга (см. рис. III-5, в; III-6; III-7, б). Он возник на базе нескольких органов чувств, которые имели представительство в этом отделе головного мозга. Основную часть крыши среднего мозга занимает представительство зрительной системы. Зрительные нервы после прохождения хиазмы перекрещиваются и поднимаются к крыше среднего мозга. Аксоны ганглиозных клеток сетчатки оканчиваются на нейронах крыши среднего мозга, которые организованы в стратифицированные структуры (см. рис. III-5, в; III-7, б). Существует чёткая топографическая связь между определённым участком сетчатки и зоной крыши среднего мозга. При этом соблюдаются форма изображения и взаимное расположение его элементов. Довольно долго переднюю часть крыши среднего мозга считали исключительно мозговым центром зрительного анализатора. Однако функционально-морфологические исследования показали, что это далеко не так.
Наряду с представительством зрительной системы в крышу среднего мозга приходят информация о соматической (кожной) чувствительности, двигательном анализаторе, вестибулярные и слуховые сигналы (см. рис. III-6, д). Слуховой анализатор у рептилий значительно увеличивает своё представительство в этом центре. В результате у многих рептилий в задней части крыши среднего мозга появляются малозаметные парные выпячивания - задние или нижние бугорки. Крыша среднего мозга становится не однородным анатомическим образованием, как у анамний, а четверохолмием. В ней сосредоточивается представительство практически всех основных дистантных и контактных анализаторов. Даже обонятельная система имеет своё представительство в крыше среднего мозга. За исключением обонятельной системы, практически все сенсорные проекции в крышу среднего мозга рептилий носят топологический характер. Это означает, что информация от каждого конкретного участка тела представлена в строго определённом участке крыши среднего мозга. Сохраняется принцип карты тела, которая точечно переносится в мозг.
Таким образом, в крыше среднего мозга рептилий сосредоточена разнообразная информация о состоянии собственного организма и окружающего мира, которая объединена по топологическому принципу. Рассмотрим, что происходит в крыше среднего мозга, если передняя правая конечность рептилии просто стоит на необычной поверхности. При оценке такой ситуации в крыше среднего мозга осуществляется сравнительный анализ соматической, сенсомоторной, слуховой и зрительной информации. Это легко сделать, поскольку все сигналы концентрируются в одном центре, а зачастую и друг над другом, как в слоистом пироге. Средний мозг проводит комплексный анализ многих факторов одного явления, что позволяет выбрать наиболее адекватную реакцию. Для этого идеально подходит стратифицированная структура крыши среднего мозга.
Предельно упрощая реальную ситуацию, можно сказать, что в крыше среднего мозга представительство различных сенсорных систем расположено на условных «этажах», организованных в горизонтальной плоскости. Каждый этаж занимает своеобразная карта. Она может быть сиюминутной информационной картой рецепторных сигналов от поверхности тела, изображением на сетчатке или акустическим полем. Все эти карты ориентированы на своих «этажах» так, что отражают примерно одно и тоже направление в пространстве. Слуховой сигнал от передней правой конечности лежит под её зрительной картой и над соматическим сигналом от кожи стопы. Специализированные «этажи» соотнесены между собой при помощи вертикальных связей, которые позволяют быстро оценить конкретную ситуацию и принять адекватное решение. Эта схема работы крыши среднего мозга позволяет понять рефлекторное быстродействие мозга рептилий. По-видимому, именно это быстродействие стало основной причиной эволюционного успеха архаичных рептилий.
Появление у рептилий совершенного рефлекторного центра принятия решений привело к нескольким важным последствиям. С одной стороны, возможность быстрого выбора решений бессмысленна, если общий уровень метаболизма будет оставаться на прежнем уровне. Следовательно, развитие среднего мозга сопровождалось повышением обмена. С другой стороны, увеличение размеров крыши среднего мозга создало необходимый клеточный субстрат для развития памяти. Индивидуальный опыт животного стал базой для сравнения событий, разнесённых во времени. Переоценить это событие трудно. Впервые возникли реальные основы для индивидуализации поведения, построенной на сравнении различных событий. Надо отметить, что эти нейробиологические преимущества мозга рептилий обычно даже не рассматриваются при реконструировании ранней эволюции амниот (Carroll, 1982).
Архаичные рептилии вслед за развитием центра рефлексологического анализа получили материальный субстрат для запоминания различных событии. Им стало доступно воспроизведение индивидуального опыта, что служит основой обучения. С самообучаемыми архаичными рептилиями амфибии уже не могли конкурировать. Гормонально-инстинктивные принципы поведения амфибий, рыб и беспозвоночных сделали их кормом для рептилий с развитым рефлекторно-ассоциативным средним мозгом.
Все перечисленные преимущества строения головного мозга рептилий не могли возникнуть сами собой. Для столь глубокой качественной перестройки мозга должны были сложиться крайне жесткие и неординарные условия. Архаичные рептилии должны были оказаться в своеобразной среде с очень высокими требованиями к аналитическим свойствам мозга и индивидуальной памяти.
Из книги Находки остатков верхнеюрских рептилий в Савельевском сланцевом руднике автора Из книги Находки остатков верхнеюрских рептилий в Савельевском сланцевом руднике автора Журавлев Константин ИвановичНаходки остатков рептилий В 1931 г., в первые же месяцы разработок сланца, когда добыча велась еще открытыми карьерами, было получено несколько находок остатков ихтиозавров. В числе этих находок серия из 13 хвостовых позвонков, найденных в пласте горючего сланца в основании
автора§ 36. Общий план строения нервной системы рептилий При масштабном освоении суши мозг рептилий изменился. Однако его морфологические перестройки, на первый взгляд, не были столь принципиальными, как у амфибий. Поверхностный анатомический анализ показывает, что древний
Из книги Происхождение мозга автора Савельев Сергей Вячеславович§ 38. Условия возникновения мозга рептилий Ассоциативный мозговой центр не может возникнуть случайно. Энергетические затраты на содержание ассоциативного центра мозга и цена смены стратегий поведения всегда очень высоки (см. главу I). Должна быть причина для подобных
Из книги Происхождение мозга автора Савельев Сергей Вячеславович§ 40. Адаптивная радиация архаичных рептилий Небольшие хищные и всеядные рептилии, эволюционировавшие в агрессивной среде растительных завалов, не были самыми крупными животными того времени. В конце карбона размер их тела обычно не превышал нескольких десятков
Из книги Биологическая химия автора Лелевич Владимир ВалерьяновичАктивный центр фермента Участок молекулы фермента, который специфически взаимодействует с субстратом, называется активным центром. Активный центр – это уникальная комбинация аминокислотных остатков в молекуле фермента, обеспечивающая непосредственное
автораГде находится центр масс системы Земля – Луна? Центр масс системы Земля – Луна, так называемый барицентр, находится на расстоянии 4672 километра от центра Земли по направлению к Луне, то есть на глубине приблизительно 1700 километров под поверхностью Земли. Строго говоря, по
Из книги Новейшая книга фактов. Том 1. Астрономия и астрофизика. География и другие науки о Земле. Биология и медицина автора Кондрашов Анатолий ПавловичЧто такое центр удовольствия и где он расположен в организме? Одной из частей головного мозга является гипоталамус, являющийся отделом промежуточного мозга и расположенный под зрительными буграми (таламусом). Гипоталамус, в котором расположены центры вегетативной
автора Кондрашов Анатолий Павлович Из книги Новейшая книга фактов. Том 1 [Астрономия и астрофизика. География и другие науки о Земле. Биология и медицина] автора Кондрашов Анатолий Павлович Из книги Фармацевтическая и продовольственная мафия автора Броуэр ЛуиЦентр фармакологического надзора «Париж, 4 февраля 1977 г. НЦФН, созданный в январе 1974 г., 2 февраля собрался на Генеральную ассамблею вместе с делегатами Национальных хартий врачей и фармацевтов, к которым присоединилась группа центров по борьбе с интоксикацией и
Из книги В дебрях времени автора Чижевский Герман МихайловичКлассификация и краткая характеристика основных групп амфибий, рептилий, птиц и млекопитающих КЛАСС АМФИБИИ, ИЛИ ЗЕМНОВОДНЫЕПодкласс апсидоспондилыНадотряд ЛабиринтодонтыГруппа амфибий, вымершая не позже триасового периода и достигшая расцвета в конце палеозойской
автора Кандель Эрик Ричард Из книги В поисках памяти [Возникновение новой науки о человеческой психике] автора Кандель Эрик Ричард Из книги Психопаты. Достоверный рассказ о людях без жалости, без совести, без раскаяния автора Кил Кент А.Мендотский реабилитационный центр для несовершеннолетних В начале 1990-х США захлестнула настоящая эпидемия подросткового насилия. Число преступлений, совершаемых несовершеннолетними, почти удвоилось между 1980 и 1993 годами. Казалось, ничто не может остановить этот
Из книги Читая между строк ДНК [Второй код нашей жизни, или Книга, которую нужно прочитать всем] автора Шпорк ПетерИз Берлина в центр будущей революции Он не обижается, когда его принимают за студента или докторанта. А это происходит с 32-летним генетиком постоянно. Александр Майсснер - серо-голубые глаза, темно-русые небрежно причесанные волосы, трехдневная щетина - не только молод,
Имя материала: Основы зоологии и зоогеографии
Общая характеристика
Благодаря появлению шеи у рептилий голова подвижно сочленена с туловищем и может совершать движения не только, как у амфибий, в вертикальном направлении, но и в горизонтальном.
Туловище удлиненное (только у черепах широкое), довольно гибкое; хвост у подавляющего большинства видов длинный, способный к разнообразным движениям; ноги сильно развиты (у некоторых видов ящериц и у всех змей ноги редуцированы в связи с переходом к другому активному способу передвижения). Величина тела в среднем значительно больше, чем у амфибий, а некоторые виды достигают огромных размеров.
Покровы. Кожа покрыта роговыми прочными и в то же время легкими чешуями эктодермального происхождения, которые защищают тело не только от механических повреждений, но и от высыхания. Ороговевший слой кожи периодически сбрасывается во время линьки. У черепах и крокодилов в мезенхимном слое кожи развиваются окостенения, у остальных рептилий они отсутствуют. Черепахи хорошо защищены от врагов панцирями. Кожа современных рептилий в отличие от земноводных сухая, лишенная желез.
Окраска наружных покровов разнообразна и у многих видов имеет защитное значение, так как благодаря ей животные малозаметны на фоне окружающей среды.
Нервная система. В связи с более подвижным и сложным образом жизни нервная система рептилий развита лучше, чем у амфибий (рис. 95).
Рис. 95. Головной мозг ящерицы, вид сбоку:
1 - полушарие переднего мозга; 2 - обонятельная доля; 3 - промежуточный мозг; 4 - гипофиз; 5 - средний мозг; 6 - перекрест зрительных нервов (хиазма); 7 - мозжечок; 8 - спинной мозг
Передний мозг значительно больших размеров, чем У последних (в особенности за счет увеличения полосатых тел), на поверхности полушарий появляются нервные клетки - зачаток коры головного мозга. Поэтому у рептилий быстрее устанавлива-ются новые связи с внешней средой, чем у их предков. Возрастает роль этого отдела головного мозга как координатора жизнедеятельности всего организма. Усложняется строение промежуточного и среднего мозга, усиливается их связь с передним мозгом и другими отделами центральной нервной системы. Мозжечок в связи с усложнением способов передвижения и необходимостью сохранять равновесие тела развит заметно лучше, чем у земноводных. Увеличился и продолговатый мозг, который образует характерный для рептилий вертикальный изгиб. Спинной мозг длинный; строение его усложнилось. Это выразилось в более резком отделении наружного слоя (белого, т. е. проводящих путей) от внутреннего (серого, т. е. нервных клеток) слоя.
Из органов чувств у рептилий (за исключением змей и некоторых других видов) лучше, чем у амфибий, развиты глаза: они более подвижны. Приспособление их к видению на разных расстояниях (у многих видов на довольно больших) обеспечивается не только, как у рыб и амфибий, перемещением хрусталика, но и, как у высших позвоночных, путем изменения его кривизны. Кроме век имеется мигательная перепонка. У ряда видов есть теменной орган, связанный с промежуточным мозгом и способный к восприятию световых раздражении. Некоторые прогрессивные изменения произошли в органе слуха: немного увеличился нижний мешочек; слуховая косточка лучше передает колебания барабанной перепонки во внутреннее ухо; кроме овального окошечка появилось второе окошечко - круглое, что увеличивает подвижность экзолимфы, окружающей лабиринт; барабанная перепонка расположена не на поверхности головы, а в небольшом углублении. Органы обоняния развиты довольно хорошо, внутренняя поверхность их увеличилась. Органы вкуса у большинства видов развиты слабо. В осязании большую роль играет язык.
Скелет. Он почти полностью окостенел благодаря замене грубой волокнистой костной ткани, характерной для земноводных, тонкой волокнистой, его прочность возросла, опорная и защитная функции улучшились (рис. 96).
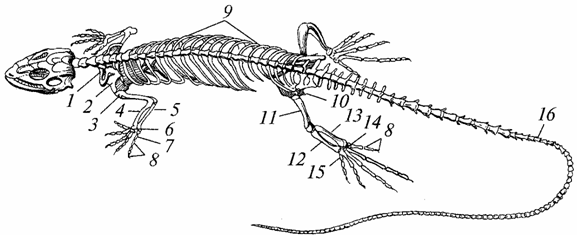
Рис. 96. Скелет ящерицы:
7 - ключица; 2 - лопатка; 3 - плечо; 4 - лучевая кость; 5 - локтевая кость; б - запястье; 7 - пястье; 8 - фаланги пальцев; 9 - ребра; 10 - таз; 11 - бедро; 12 - большая берцовая кость; 13 - малая берцовая кость; 14 - предплюсна; 15 - плюсна; 16 - хвостовые позвонки
Череп увеличился; вместимость мозгового отдела стала большей, чем у амфибий. Боковая поверхность черепа у древних рептилий была сплошной, позднее в ней появились височные ямы - одна или две, что уменьшило его массу и увеличило поверхность для прикрепления жевательных мышц. Нижние челюсти, как и у амфибий, подвешены к мозговому отделу черепа при помощи квадратной кости и тоже состоят из нескольких костей (зубной, сочленовной и др.).
Тело первого позвонка (атланта) приросло в виде зубца к телу второго позвонка (эпистрофея). Кольцевидный атлант получил возможность вращаться вокруг зубца эпистрофея, благодаря чему стали возможны и повороты головы, а это имело большое значение для добывания пищи (в основном других животных) и для защиты от врагов. Атлант соединен с затылочным отделом черепа с помощью одного мыщелка, что также способствует увеличению подвижности головы. Позвоночный столб разделяется на пять отделов: шейный, грудной, поясничный, крестцовый и хвостовой. Позвонки грудного отдела соединяются с грудиной хорошо развитыми ребрами, вследствие чего образуется грудная клетка, отсутствующая у амфибий. От поясничных позвонков отходят укороченные ребра, не доходящие до грудины. Крестцовых позвонков два (у амфибий один). В хвостовом отделе (за исключением черепах) позвонков много.
Кости ног лучше развиты, и они более прочные, чем у земноводных. Между костями запястья и предплюсны имеются подвижные суставы. Поэтому предплечье и проксимальный ряд запястья, а также голень и проксимальный ряд предплюсны функционируют как целостные образования. Особо нужно отметить усиление поясов конечностей: плечевого - благодаря его соединениям с грудиной и грудным отделом позвоночника, тазового - вследствие того, что он опирается на два крестцовых позвонка и обе его половины образуют два сращения - седалищное и лонное.
Мышечная система. Она подверглась дифференциации, развились новые группы мышц, от которых зависят повороты головы, движения возникшей у рептилий шеи, расширение и сужение грудной клетки и др. Значительно увеличилась мускулатура ног, развились сгибатели и разгибатели пальцев. Масса и сила всей системы возросли.
Пищеварительная система. Она более дифференцирована, чем у земноводных (рис. 97). Зубы в основном сосредоточены на челюстях, количество их велико, но они однотипны и служат преимущественно для захвата добычи. Слюнные железы лучше развиты, чем у амфибий (у ряда видов выделения этих желез ядовиты). У рептилий начинается процесс образования твердого нёба, которое особенно развито у крокодилов. Благодаря образованию твердого нёба ротовая полость отделяется от носоглоточной, что облегчает как дыхание, так и поглощение пищи. Хорошо развиты (особенно у хищников, поедающих позвоночных животных) глотка, пищевод и желудок. Печень (с желчным пузырем) и поджелудочная железа более совершенны, чем у земноводных. Тонкая кишка значительно удлинена, толстая кишка короткая и заканчивается клоакой. Современные пресмыкающиеся питаются преимущественно животными, число растительноядных видов незначительно.
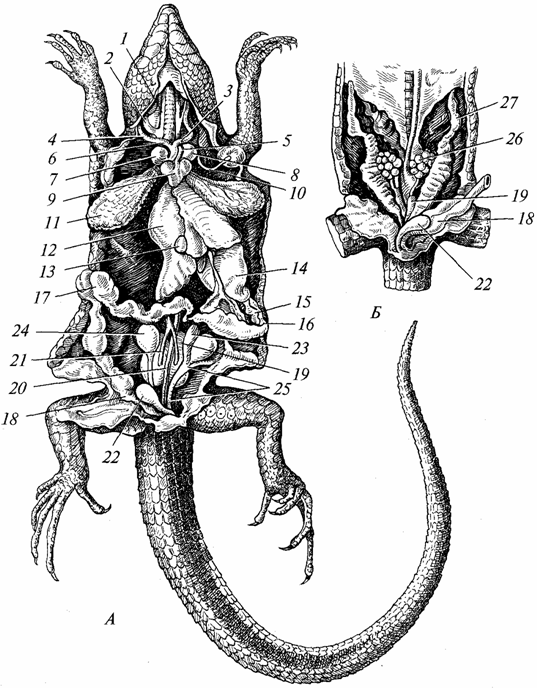
Рис. 97. Внутренние органы ящерицы; А - самец; Б - самка:
/ - внешняя и 2 - внутренняя яремные вены; 3 - левая и 4 - правая сонные артерии; 5 - левая и 6 - правая дуги аорты; 7 - правое и 8 - левое предсердия; 9 - желудочек сердца; 10 - левая подключичная артерия- 11 -легкое; 12 - печень; 13 - желчный пузырь; 14 - желудок; /5 - поджелудочная железа; 16 - двенадцатиперстная кишка; 17 - толстая кишка- 18 - прямая кишка; 19 - почка; 20 - спинная аорта; 21 - выносящие вены почек; 22 - мочевой пузырь; 23 - придаток семенника; 24 - семенник; 25 - семяпровод; 26 - яичник; 27 - яйцепровод
Дыхательная система. Органами дыхания у рептилий служат легкие (см. рис. 97), так как их кожа непригодна для газообмена; слизистая оболочка ротовой полости, играющая значительную роль в дыхании земноводных, используется для выполнения этой функции немногими видами пресмыкающихся. По этой причине и особенно в связи с возросшей активностью рептилий по сравнению с амфибиями строение легких и дыхательных путей усложнилось. Легкие представляют собой органы большей величины и глубже расположены в грудной полости, чем у земноводных. Внутренняя поверхность их значительно возросла благодаря развитию на ней множества перекладин с более густой сетью кровеносных сосудов. Воздух поступает в легкие, пройдя через хоаны, глотку, гортань, длинную трахею, просвет которой поддерживается хрящевыми кольцами. Во время прохождения через трахею воздух очищается от пыли, температура его приближается к температуре тела. У амфибий трахея была в зачаточном состоянии. Задний конец трахеи разделяется на два бронха, в стенках которых тоже имеются хрящевые кольца. Осуществление вдоха и выдоха происходит более эффективно, чем у амфибий, благодаря развитию у рептилий грудной клетки, объем которой увеличивается при вдохе и уменьшается при выдохе. Такой же механизм сохранился у высших позвоночных - птиц и млекопитающих.
Кровеносная система. Сердце, как и у амфибий, трехкамерное, но в желудочке снизу появляется зачаток перегородки. У крокодилов она уже полная, и сердце у них становится четырехкамерным (два предсердия и два желудочка). Следовательно, у пресмыкающихся усиливается разделение венозного и артериального токов крови, но полной изоляции обоих токов еще нет даже у крокодилов, как это видно из нижеследующего описания (рис. 98).
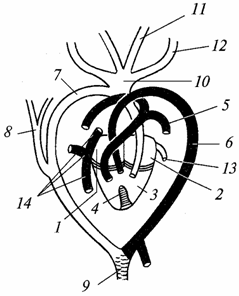
Рис. 98. Сердце и основные сосуды ящерицы (светлые - сосуды с артериальной кровью, штриховка - со смешанной кровью; черные - с венозной):
1 - правое предсердие; 2 - левое предсердие; 3 - желудочек; 4 - неполная перегородка, разделяющая желудочек на левую и правую половины; 5 - легочная артерия; 6 - левая дуга аорты; 7 - правая дуга аорты; 8 - подключичная артерия; 9 - спинная аорта; 10 - общий ствол сонных артерий; 11 - левая внутренняя сонная артерия, 12 - левая наружная сонная артерия; 13 - легочная вена; 14 - задняя полая вена и передние полые (яремные) вены
В левое предсердие поступает артериальная кровь - она идет из легких. В правое предсердие поступает венозная кровь - она собирается из всех частей тела. Далее кровь из обоих предсердий поступает в желудочек: в правую его половину - венозная, в левую - артериальная. Благодаря наличию перегородки, хотя и неполной, кровь в желудочке меньше смешивается, чем у земноводных. Артериального конуса нет, и главные артериальные стволы отходят непосредственно от сердца.
От правой стороны желудочка отходят легочные артерии (бывшая четвертая пара жаберных артерий), несущие в легкие венозную кровь. Бывшая третья пара жаберных сосудов атрофировалась. Вторая пара сосудов состоит из двух дуг аорты: правая дуга начинается от левой части желудочка и несет артериальную кровь, левая дуга начинается от правой части желудочка (но ближе к левой его части) и несет смешанную кровь. От правой дуги аорты отходят сонные артерии (бывшая первая пара жаберных сосудов), идущие к голове. Они снабжают мозг, особенно чувствительный к недостатку кислорода, артериальной кровью. Обе дуги аорты соединяются в аорту, которая разветвляется на множество артерий. Таким образом, организм пресмыкающегося в основном снабжается смешанной кровью, но она насыщена большим количеством кислорода, чем у земноводных, что способствует более интенсивному обмену веществ.
Выделительная система. Она сильно изменилась в связи с усилением интенсивности обмена веществ рептилий, что привело к увеличению количества продуктов диссимиляции, и полным приспособлением их к наземному образу жизни, вследствие чего возникла необходимость более экономно использовать поступающую в организм воду. Поэтому у рептилий развились более сложные органы выделения, названные тазовыми или вторичными почками {метаяефрос). Все канальцы этих почек не имеют воронок и начинаются боуменовыми капсулами, окружающими мальпигие-вы тельца с увеличенным количеством капилляров. Канальцы тазовых почек очень длинные и густо оплетены капиллярами. В боу-меновых капсулах из просочившихся туда из мальпигиевых телец растворенных в воде продуктов диссимиляции и некоторых органических веществ (например, глюкозы) образуется первичная моча. Последняя во время прохождения через длинные почечные канальцы отдает обратно в кровь через оплетающие их стенки капилляры значительную часть воды и другие вещества, необходимые для организма. Образовавшаяся после этого вторичная моча выходит не через вольфовы каналы, а через заново развившиеся мочеточники в клоаку и накапливается в мочевом пузыре. Процессы образования первичной и вторичной мочи совершаются и в туловищных почках, но с возвратом меньшего количества воды в кровь. Первичноводные же позвоночные, в особенности земноводные, кожа которых голая, часто страдают от избытка воды, а не от ее недостатка. Следует также отметить, что у рептилий конечным продуктом разложения белков является мочевая кислота, на выведение которой из организма с мочой затрачивается мало воды. У земноводных и у ряда других первичноводных позвоночных разложение белков приводит к образованию мочевины, на удаление которой воды требуется много.
У эмбрионов рептилий есть зачаток головной почки и развитая туловищная почка, которая функционирует у взрослых особей (у ряда видов ящериц) до наступления половой зрелости. Эти факты подтверждают преемственность трех видов почек в течение эволюции позвоночных.
Размножение. В состав мужской половой системы рептилий входят два семенника округлой формы, лежащие по бокам поясничного отдела позвоночника, два придатка семенников, образовавшиеся из передних участков туловищных почек, два семяпровода (вольфовы каналы) и совокупительные органы, развившиеся из стенки клоаки. Женская половая система состоит из двух яичников, сильно разрастающихся в период размножения, и двух яйцеводов (мюллеровых каналов), впадающих в клоаку. Оплодотворение только внутреннее. Семя вводится самцами в клоаку самки при помощи совокупительных органов. Созревшие яйцеклетки, содержащие много желтка, попадают в воронки яйцеводов. Соединение женских и мужских гамет происходит в верхней части яйцеводов, куда проникают после полового акта сперматозоиды. Зиготы, продвигаясь по яйцеводам, в средней части последних окружаются белковой оболочкой, а в задней покрываются скорлупой.
У некоторых видов ящериц и змей наблюдается партеногенетическое размножение самок. Самцов у таких видов, как правило, нет.
Развитие. У рептилий развитие прямое; личинок у рептилий нет. Оплодотворенные яйца пресмыкающиеся обычно откладывают в почву, а некоторые виды - под гниющие растительные остатки. У ряда видов развитие зигот совершается в половых путях самки и сразу после выхода развившихся яиц наружу из них вылупляются сформированные животные, т. е. для таких видов характерно яйцеживорождение. У немногих видов наблюдается настоящее живорождение: сосуды желточного мешка тесно соединяются с сосудами яйцевода, и эмбрион получает значительную часть питательных веществ из организма матери. Яйцеживорождение чаще наблюдается у видов, распространенных в северных и горных районах. Очевидно, яйцеживорождение является защитой развивающихся зародышей от охлаждения и других неблагоприятных условий. Нет ни одного вида рептилий, у которого яйца развиваются в воде. Даже у рептилий, проводящих большую часть жизни в воде (все крокодилы, многие черепахи, некоторые змеи), откладка яиц и их развитие совершаются только на суше. Морские змеи, никогда не покидающие воду, относятся к живородящим.
Яйца и зародыши рептилий обладают рядом приспособлений для развития на суше. Во-первых, яйца покрыты скорлупой и другими оболочками, которые защищают их от высыхания и повреждений. Во-вторых, у зародышей развиваются особые зародышевые оболочки (серозная, амнион и аллантоис), которые имеют большое значение для обеспечения эмбрионального развития животных на суше. Серозная оболочка и амнион развиваются из одной и той же двойной складки тела зародыша, происходящей из эктодермы и мезодермы. Эта складка появляется с двух противоположных сторон в виде зачатков, которые растут навстречу друг другу и смыкаются над зародышем. Наружная часть складки превращается в серозу (ее наружный слой эктодермальный, внутренний - мезодермальный), а внутренняя - в амнион (его наружный слой мезодермальный, внутренний - эктодермальный). Полость, образуемая амнионом, наполнена водянистой жидкостью, благодаря чему развитие происходит во влажной среде, и зародыш защищен от высыхания и последствий различных сотрясений, особенно опасных в условиях воздушной среды.
Происхождение. Первые рептилии, возникшие во второй половине каменноугольного периода, обитали во влажных биотопах суши и, вероятно, размножались и развивались в воде. К этому времени суша уже была заселена низшими наземными растениями, различными беспозвоночными, начиналось завоевание воздуха насекомыми. Рептилии, которые могли удаляться на большие расстояния, чем амфибии, от водоемов, имели в изобилии пищу и не встречали серьезных врагов. В конце каменноугольного периода и неоднократно в течение пермского периода климат становился более засушливым, что вызывало гибель многих амфибий и, наоборот, содействовало прогрессивному развитию и более широкому распространению на суше рептилий. Спустя некоторое время они перешли к размножению и развитию вне водоемов, сначала в более влажных наземных биотопах, а потом в более сухих местах. В течение мезозойской эры они распространились по всему земному шару и возникло много различных групп. Наряду с хищниками "было много растительноядных видов, появились рептилии, приспособленные к полету, и рептилии, обитающие в морях. Величина их тела была различна, но большинство были крупными животными, а некоторые имели гигантские размеры - до 20 м и более в длину. Климат на протяжении почти всей мезозойской эры был теплым, очень благоприятным для этих животных, не имеющих достаточно высокой постоянной температуры тела и теряющих свою активность при охлаждении окружающей среды.
В конце мезозойской эры на земном шаре происходили мощные горообразовательные процессы, сопровождающиеся резким изменением условий существования (температуры, влажности и др.). Это повлекло за собой массовое вымирание мезозойских рептилий, не выдержавших борьбу за существование в таких тяжелых для них абиотических условиях среды, осложненную соревнованием с млекопитающими и птицами. Тогда же возникли и стали многочисленными в течение кайнозойской эры новые группы пресмыкающихся - ящерицы и произошедшие от них змеи. Из старых же групп сохранились лишь один очень примитивный вид первоящеров, небольшое количество крокодилов и черепах. Современные рептилии распространены преимущественно в теплых странах, особенно в субтропических и тропических, в странах с умеренным климатом их мало, на далеком севере они отсутствуют.