Липиды 200401
ХИМИЯ И ОБМЕН ЛИПИДОВ.
ЛИПИДЫ - органические вещества, характерные для живых организмов, нерастворимые в воде, но растворимые в органических растворителях и друг в друге.
Группы липидов отличаются по степени гидрофобности. Фосфолипиды и гликолипиды являются полярными липидами.
Холестерин занимает промежуточное положение между полярными и абсолютно гидрофобными липидами.
Абсолютно гидрофобными являются триглицериды и эфиры холестерина.
Большинство липидов (кроме стеринов и некоторых минорных липидов) содержат высшие жирные кислоты (ВЖК).
В состав мембран входят только ФОСФОЛИПИДЫ (ФЛ), ГЛИКОЛИПИДЫ (ГЛ) и ХОЛЕСТЕРИН (ХС).
ЖИРНЫЕ КИСЛОТЫ, ХАРАКТЕРНЫЕ ДЛЯ ОРГАНИЗМА ЧЕЛОВЕКА
Кислота называется жирной, если число углеродных атомов в ее молекуле больше четырех. Преобладают длинноцепочечные жирные кислоты (число атомов углерода 16 и выше).Количество углеродных атомов и двойных связей обозначается двойным индексом. Например: С18:1 (9-10). В данном случае 18 – число атомов углерода и 1 – количество двойных связей. В скобках указывается местоположение двойных связей (по номерам углеродных атомов).
Студенты должны знать количество углеродных атомов и двойных связей в молекулах следующих жирных кислот:
С16:0 - пальмитиновая,
С18:0 - стеариновая,
С18:1 – олеиновая (9:10),
С18:2 – линолевая (9-10,12-13),
С18:3 - линоленовая (9-10, 12-13, 15-16),
С20:4 - арахидоновая (5-6, 8-9, 12-13, 15-16).
Жирные кислоты, имеющие двойную связь в положении 3, обладают очень выраженным антиатеросклеротическим действием (линоленовая). Особенно много 3-содержащих жирных кислот находится в рыбьем жире и других морепродуктах. В организме человека такие жирные кислоты не синтезируются.
Жирные кислоты, входящие в состав организма человека, имеют общие черты строения:
Чётное число атомов углерода.
Линейная (неразветвлённая) углеродная цепь.
Полиненасыщенные жирные кислоты имеют ТОЛЬКО ИЗОЛИРОВАННЫЕ двойные связи (между соседними двойными связями не меньше двух одинарных).
Двойные связи имеют только цис-конфигурацию.
По количеству двойных связей жирные кислоты можно разделить на НАСЫЩЕННЫЕ (нет двойных связей), МОНОНЕНАСЫЩЕННЫЕ (есть одна двойная связь) и ПОЛИНЕНАСЫЩЕННЫЕ (две или более двойных связей).
Свободные жирные кислоты встречаются в очень небольшом количестве. В основном они находятся в составе других липидов. При этом они связаны с другими компонентами липидов сложноэфирной связью (эстерифицированы).
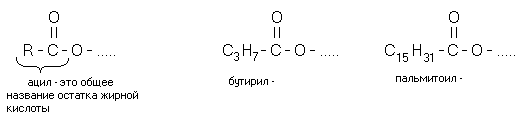
ФОРМЫ СУЩЕСТВОВАНИЯ ЛИПИДОВ В ОРГАНИЗМЕ ЧЕЛОВЕКА
Жировые включения, которых больше всего в адипоцитах. Это форма существования триглицеридов.
Биомембраны. Они не содержат триглицеридов, зато в них присутствуют фосфолипиды, гликолипиды и холестерин.
Липопротеины. Могут включать в себя липиды всех классов.
Липопротеины
Липопротеины - это сферические частицы, в которых можно выделить гидрофобную сердцевину, состоящую из триглицеридов (ТРГ) и эфиров холестерина (ЭХС) и амфифильную оболочку, в составе которой – фосфолипиды, гликолипиды и белки.
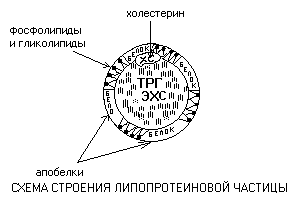
Белки оболочки называются апобелками. Холестерин (ХС) обычно занимает промежуточное положение между оболочкой и сердцевиной. Компоненты частицы связаны слабыми типами связей и находятся в состоянии постоянной диффузии – способны перемещаться друг относительно друга.
Основная роль липопротеинов – транспорт липидов, поэтому обнаружить их можно в биологических жидкостях.
При изучении липидов плазмы крови оказалось, что их можно разделить на группы, так как они отличаются друг от друга по соотношению компонентов. У разных липопротеинов наблюдается различное соотношение липидов и белка в составе частицы, поэтому различна и плотность.
Липопротеины разделяют по плотности методом ультрацентрифугирования, при этом они не осаждаются, а всплывают (флотируют). Мерой всплывания является константа флотации, обозначаемая S f (сведберг флотации). В соответствии с этим показателем различают следующие группы липопротеинов:

Липопротеины можно разделить и методом электрофореза. При классическом щелочном электрофорезе разные липопротеины ведут себя по-разному. При помещении липопротеинов в электрическое поле хиломикроны остаются на старте. ЛОНП и ЛПП можно обнаружить во фракции пре-глобулинов, ЛНП - во фракции -глобулинов, а ЛВП - -глобулинов:
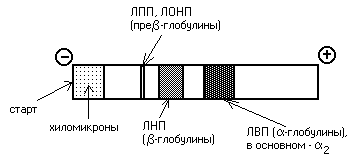
Определение липопротеинового спектра плазмы крови применяется в медицине для диагностики атеросклероза.
Все эти липопротеины отличаются по своей функции.
1. Хиломикроны (ХМ) - образуются в клетках кишечника, их функция: перенос экзогенного жира из кишечника в ткани (в основном - в жировую ткань), а также - транспорт экзогенного холестерина из кишечника в печень.
2. Липопротеины Очень Низкой Плотности (ЛОНП) - образуются в печени, их роль: транспорт эндогенного жира, синтезированного в печени из углеводов, в жировую ткань.
3. Липопротеины Низкой Плотности (ЛНП) - образуются в кровеносном русле из ЛОНП через стадию образования Липопротеинов Промежуточной Плотности (ЛПП). Их роль: транспорт эндогенного холестерина в ткани.
4. Липопротеины Высокой Плотности (ЛВП) - образуются в печени, основная роль - транспорт холестерина из тканей в печень, то есть удаление холестерина из тканей, а дальше холестерин выводится с желчью.
При определении содержания в крови липопротеинов различной плотности их обычно разделяют методом электрофореза. При этом ХМ остаются на старте, ЛОНП оказываются во фракции пре-глобулинов, ЛНП и ЛПП находят во фракции -глобулинов, а ЛВП - 2 -глобулинов. Если в крови повышено содержание -глобулинов (ЛНП) - это означает, что холестерин откладывается в тканях (развивается атеросклероз).
РОЛЬ АПОБЕЛКОВ В СОСТАВЕ ЛИПОПРОТЕИНОВ
Белковые компоненты липопротеинов особенно важны. Они называются "аполипопротеинами " и обозначаются латинскими буквами (апоА, апоВ, апоС и так до апоG).
ФУНКЦИИ АПОБЕЛКОВ .
1. Апобелки выполняют функцию эмульгаторов, потому что являются амфифильными веществами.
2. Некоторые из аполипопротеинов являются регуляторами активности ферментов липидного обмена.
3. Могут обладать собственной ферментативной активностью.
Могут выступать в качестве лигандов клеточных рецепторов для липопротеинов.
Многие апобелки осуществляют транспорт липидов из одного липопротеина в другой.
ОБМЕН ЖИРА (ТРИАЦИЛГЛИЦЕРИНА)
Химическое название жиров - ацилглицерины, то есть жиры. Это сложные эфиры глицерина и высших жирных кислот. "Ацил-" - это означает "остаток жирных кислот" (не путать с "ацетил-" - остатком уксусной кислоты). В зависимости от количества ацильных радикалов жиры разделяются на моно-, ди- и триглицериды. Если в составе молекулы 2 радикала жирных кислот, то жир называется ДИАЦИЛГЛИЦЕРИНОМ. Если в составе молекулы 1 радикал жирных кислот, то жир называется МОНОАЦИЛГЛИЦЕРИНОМ.
В организме человека и животных преобладают ТРИАЦИЛГЛИЦЕРИНЫ (содержат три радикала жирных кислот).
Свойства жира определяются составом жирных кислот.
Функции жира:
1. ЭНЕРГЕТИЧЕСКАЯ.
В отличие от углеводов жиры составляют энергетический резерв организма. Преимущество жира в качестве энергетического резерва заключается в том, что жиры являются более восстановленными веществами по сравнению с углеводами (в молекулах углеводов при каждом углеродном атоме есть кислород – группы “–CHOH-“; у жира имеются длинные углеводородные радикалы, в которых преобладают группы “-CH 2 -“ - в них нет кислорода). От жира можно отнять больше водорода, который затем проходит по цепи митохондриального окисления с образованием АТФ.
Калорийность углеводов и белков: ~ 4 ккал/грамм.
Калорийность жира: ~ 9 ккал/грамм.
Преимуществом жира как энергетического резерва, в отличие от углеводов, является гидрофобность – он не связан с водой. Это обеспечивает компактность жировых запасов - они хранятся в безводной форме, занимая малый объем.
В среднем, у человека запас чистых триацилглицеринов составляет примерно 13 кг. Этих запасов могло бы хватить на 40 дней голодания в условиях умеренной физической нагрузки. Для сравнения: общие запасы гликогена в организме – примерно 400 гр.; при голодании этого количества не хватает даже на одни сутки.
2.ЗАЩИТНАЯ.
Жировая ткань:
а) защищает органы от механических повреждений.
б) участвует в терморегуляции.
Образование запасов жира в организме человека и некоторых животных рассматривается как приспособление к нерегулярному питанию и к обитанию в холодной среде. Особенно большой запас жира у животных, впадающих в длительную спячку (медведи, сурки) и приспособленных к обитанию в условиях холода (моржи, тюлени). У плода жир практически отсутствует, и появляется только перед рождением.
ИСТОЧНИКИ ЖИРА В ОРГАНИЗМЕ:
1. Пищевой жир (экзо генный)
2. Эндо генный жир, синтезируется в печени из углеводов.
Липиды нерастворимы в воде, поэтому они транспортируются в крови в комплексе с белками. Жирные кислоты в крови ассоциированы с альбумином, другие липиды транспортируются в составе липопротеинов (ЛП).
Один из факторов, способствующих развитию атеросклеротического процесса, - нарушение обмена ЛП. Этот факт на протяжении последних 25 лет послужил причиной широких исследований структуры ЛП, их синтеза, метаболических превращений и интернализации ЛП-частиц клетками. В многочисленных работах представлены данные о свойствах и составе ЛП-частиц разных классов, а также об особенностях биосинтеза и катаболизма липидных и белковых компонентов.
На рис. 1.5 представлены структурные формулы основных липидов плазмы крови.
Свободные, или неэтерифицированные, жирные кислоты - основной поставщик энергии в организме. В крови человека присутствуют также насыщенные, мононенасыщенные и полине- насыщенные жирные кислоты (табл. 1.7).
Таблица 1.7. Жирные кислоты, присутствующие в плазме Жирные кислоты Название Длина
цепи Источник Миристиновая С 14:0 Кокосовое масло Насыщенные Пальмитиновая С 16:0 Животный жир Стеариновая С 18:0 Животный жир Мононенасыщенные Пальмитолеиновая С 16:1 со 7 Жир Олеиновая С 18:1 со9 Сливочное масло Полиненасыщенные Линоленовая С 18:2 соб Растительное
масло (полиеновые) Арахидоновая С 20:4 со8 Растительное
масло Эйкозапентаеновая С 20:5 соЗ Рыбий жир Примечание. В сокращенной формуле указаны количество атомов углерода и число двойных связей. Ближайшая к метильному концу двойная связь обозначена символом со.
ТГ - это эфиры глицерина и жирных кислот; они так же, как и жирные кислоты, служат важным источником энергии.
Фосфолипиды - эфиры глицерина или сфингозина с жирными кислотами и фосфорной кислотой. В состав фосфолипидов входят также азотсодержащие соединения (холин, этаноламин, серин) или другие гидрофильные группы, соединенные эфирной связью с фосфатом. Фосфолипиды - компоненты всех клеточных мембран и ЛП.
СН3(СН2)„СОСГ Жирные кислоты (п> 14)
Триглицериды ®
О СН2 - О - С - (СН,)„ -сн3II I
СН, -(СН2)„ - с - О-сн о
II
сн, - о - с - к.
Фосфолипиды О
СН, - О - С - (СН,)„ -сн,
I
СН, ~(СН2)„ - с - о-сн о
сн2 - О - Р - о -1
I
о
Рис. 1.5. Структурные формулы липидов.
Холестерин - стероид, на основе которого образуются стероидные гормоны и желчные кислоты. Около 2/3 холестерина в крови представлено в форме эфиров холестерина и жирных кислот, около 1/3 - в форме свободного холестерина. Свободный холестерин входит в состав всех клеточных мембран; в составе ЛП имеется как свободный, так и этерифицированный холестерин.
В клинической практике для выявления нарушений липидного обмена определяют уровень холестерина и ТГ в сыворотке крови.
ЛИПОПРОТЕИНЫ
Частицы ЛП - макромолекулярные комплексы, внутренняя часть которых содержит нейтральные липиды (ТГ и эфиры холестерина), а поверхностный слой состоит из фосфолипидов, неэте- рифицированного холестерина и специфических липидтранспорт- ных белков, называемых аполипопротеинами (апоЛП).
ЛП классифицируют на основании подвижности их в электрическом поле (при проведении ЭФ) или гидратированной плотности в условиях усиленной гравитации при препаративном ультрацентрифугировании (флотация или седиментация). При проведении ЭФ ЛП подразделяют на фракции, одна из которых остается на старте (хиломикроны), другие мигрируют к зонам глобулинов - Р-ЛП, пре-р-ЛП, а-ЛП. По величине гидратированной плотности ЛП принято разделять на 5 классов: хиломикроны (ХМ), ЛП очень низкой плотности (ЛПОНП), ЛП промежуточной плотности (ЛППП), ЛП низкой плотности (ЛПНП), ЛП высокой плотности (ЛПВП). По электрофоретической подвижности ЛПОНП соответствуют пре-р-ЛП, ЛПНП - Р-ЛП, ЛПВП - а-ЛП, а ХМ остаются на старте (рис. 1.6).
АпоЛП способствуют формированию мицелл ЛП в эндо- плазматическом ретикулуме гепатоцитов, служат лигандами для специфических рецепторов на поверхности плазматической мембраны клеток и кофакторами (активаторами и ингибиторами) процессов липолиза и метаболизма ЛП в сосудистом русле (табл. 1.8).
В зависимости от роли апоЛП в организации первичных частиц ЛП и их последующих превращениях апопротеины услов
но можно разделить на два класса. К одному из них следует отнести апоЛП, которые формируют мицеллярную структуру ЛП-комплексов и служат ядром ЛП-частиц. Особенностью таких апопротеинов является то, что белки не покидают ЛП-частицу, в формировании которой они участвуют. В эту группу апоЛП входят апоВ (апоВ-100 и апоВ-48) и апоА (А-1 и А-Н), ответственные за осуществление афферентного и эфферентного транспорта липидов. АпоВ - основной структурный белок богатых триглицеридами ЛП-частиц; он не покидает мицеллярный
Таблица 1.8. Свойства и функции аполипопротеидов ДпоЛП Входит в состав Место
синтеза Мол.
масса,
кД Функции С* в
плаз-ме, мг/л А-1 ЛПВП,
хм Тонкая
кишка,
печень 28,3 Активация ЛХАТ, транспорт липидов, лиганд для ЛПВП-рецептора 100-1600 А-Н лпвп Тонкая
кишка,
печень 17 Структурный белок, лиганд для ЛПВП-рецептора 300-500 А-1У хм,
лпвп,
лпонп Тонкая
кишка 46 Активация ЛХАТ, лиганд для ЛПВП-рецептора 150 В-100 лпнп,
лппп,
лпонп Печень 549 Перенос липидов из печени, связывание с В-, Е-рецептором 500-900 В-48 хм Тонкая
кишка 265 Перенос липидов из тонкой кишки лпонп,
лпвп Печень 6,5 Активатор ЛХАТ лпонп,
лпвп Печень 8,8 Активатор ЛПЛ 30-80 С-Ш хм,
лпонп.
лпвп Печень 8,9 Ингибитор активности ЛПЛ 8-150 Б ЛПВПз 20 Активатор ЛХАТ ~100 Е лпонп,
хм,
лпвп Печень 36,5 Связывание с В- и Е-рецепторами 30-50 (а) ЛП(а) Печень 270-1000 Неизвестны комплекс в процессе последовательных метаболических превра-щений ЛПОНП в ЛППП и далее в ЛПНП, накопление которых в сосудистой стенке служит патогенетическим звеном атероскле-ротического процесса. Апопротеины А-1 и А-Н - основные белки ЛПВП. Основой предположения, что апоВ и апоА формируют разные по составу и функции классы ЛП, послужил факт, что апоВ и апоА не присутствуют вместе в длительно циркулирующих ЛП-частицах. Генетические нарушения синтеза этих апо- протеинов - причина нарушений афферентного и эфферентного транспорта липидов.
К другому классу можно отнести апоЛП, их основная роль - регуляция метаболизма ЛП в сосудистом русле и процесса интернализации их клетками. Эти апопротеины содержатся в ЛП в значительно меньших количествах и в процессе взаимопревращения ЛП-частиц в кровеносном русле перемещаются между ЛП разных классов в виде белково-липидных комплексов. Метаболически активные апоЛП находятся в сосудистом русле дольше, чем частицы ЛП, в состав которых они были включены в момент синтеза. Основные представители группы метаболически активных апоЛП - апоЕ (с изоформами Е2, ЕЗ, Е4) и апоС (С-1, С-Н, С-Ш).
АпоЛП имеют большое значение для метаболизма ЛП, играя роль лигандов ЛП-частиц, взаимодействующих с клеточными ре-цепторами для специфических ЛП. АпоВ-100 и апоЕ взаимодей-ствуют с рецептором ЛПНП, начиная абсорбционный эндоцитоз, сопровождающийся катаболизмом ЛПНП. АпоЕ взаимодействует с рецептором остаточных компонентов ХМ, играющего важную роль в удалении этих компонентов печенью. Предполагают, что апоА-1 взаимодействует с рецептором к ЛПВП и облегчает удаление холестерина из периферических клеток для обратного транспорта в печень.
В плазме содержатся 4 основных класса ЛП: ХМ, ЛПОНП, ЛПНП и ЛПВП и несколько других ЛП, представленных в сравнительно низких концентрациях: ЛППП и ЛП(а). В табл. 1.9 представлены некоторые свойства ЛП.
ХМ синтезируются в кишечнике и служат для переноса экзогенных липидов. Основной белок насцентных (первичных) ХМ - апоВ-48. Вскоре после секреции ХМ энтероцитами в их состав в сосудистом русле включаются 2 дополнительных апопротеина, содержащихся в ЛПВП, - апоС-Н и апоЕ. АпоС-Н активирует гепаринзависимую липопротеинлипазу - фермент, фиксирован- Свойства ХМ лпонп лппп ЛПНП ЛПВП Плотность, г/мл подвижность Остаются на старте Пре-р Флотирующие
Р Р а Место образования Тонкая кишка Печень Катаболизм
ЛПОНП Катаболизм ЛПОНП через ЛППП Печень, тонкая кишка, катаболизм ХМ и ЛПОНП Основная функция Транспорт экзогенных ТГ Транспорт эндогенных ТГ Предшественник ЛПНП Транспорт
холестерина Обратный
транспорт
холестерина Состав: ТГ 90% 65% 20% 5% 5% холестерин 5% 15% 25% 50% 20% фосфолипиды 4% 10% 35% 25% 25% белок 1% 10% 20% 20% 50% апопротеины А, В-48, С, Е В-100, С, Е В-100, Е В-100 А, С, Е Таблица 1.9. Характеристика липопротеидов
1.5. Липиды и липопротеины -ф- 143
ный на эндотелии капилляров. Липопротеинлипаза гидролизует ТГ хиломикронов с образованием свободных жирных кислот. В процессе гидролиза ЛП частицы теряют около 96% своей массы, в основном за счет ТГ, а также апоЛП А и С. В результате этого ЛП-частица преобразуется в меньший по размерам, обогащенный холестерином остаток ХМ, в котором апоВ и апоЕ служат основными апоЛП. Образованные остаточные компоненты ХМ имеют гидратированную плотность ЛПОНП и ЛППП, и в норме из сыворотки крови их элиминируют гепатоциты посредством специфических рецепторов (апоЕ-рецепторы). Поэтому у здоровых лиц промежуточные формы ЛП имеют короткий период жизни и в физиологических условиях находятся в сыворотке крови в низкой концентрации. Однако при некоторых формах ГЛП происходит накопление ЛППП в сыворотке крови. В результате этих метаболических превращений транзиторная ГЛП, имеющая место в течение нескольких часов после приема пищи, устраняется, после чего утилизация экзогенных липидов идет в печени. Поступившие в печень липиды по жирнокислотному составу соответствуют экзогенным липидам. Секретируемые печенью в кровоток ЛП-частицы имеют жирнокислотный состав, свойственный организму человека.
ЛПОНП образуются в печени, секретируются гепатоцита- ми в кровоток и транспортируют эндогенные ТГ. Насцентные ЛПОНП имеют в составе только апоВ-100. В кровотоке в состав ЛПОНП также включаются апоЛП из ЛПВП - апоС-П и апоЕ. Исследования метаболизма показали, что радиоактивность, первоначально присутствующая во фракции ЛПОНП, быстро оказывается в ЛП-частицах с гидратированной плотностью ЛПНП. У здоровых людей практически все ЛПНП образованы в сосудистом русле из ЛПОНП. В ходе метаболизма богатых ТГ ЛП, содержащих апоВ, вследствие гидролиза ТГ ЛП меньшей плотности и больших размеров преобразуются в более плотные и меньшие по размеру ЛППП и затем ЛПНП. Некоторое количество остаточных компонентов ЛПОНП и ЛППП покидает сосудистое русло посредством взаимодействия со специфическими апоВ/Е-рецепторами на поверхности эндотелиальных клеток. В процессе метаболического превращения ЛППП в ЛПНП, кроме гепаринзависимой липопротеинлипазы, принимает участие еще один липолитический фермент - триглицеридлипаза печени. Этот фермент способен действовать не только как гидролаза ТГ, но и как фосфолипаза.
ЛПНП у человека содержат большую часть циркулирующего холестерина и транспортируют его к периферическим тканям для формирования мембран и процессов стероидогенеза. ЛПНП взаимодействуют с рецепторами на плазматической мембране клеток печени, надпочечников и периферических тканей, включая ГМК и фибробласты. После взаимодействия с рецептором ЛПНП подвергаются эндоцитозу и компоненты ЛПНП катаболи- зируются в лизосомах. В результате этих процессов образуются отдельные классы липидов; апобелки разрушаются до аминокислот.
ЛП(а) - сходная с ЛПНП, обогащенная холестерином и белком частица, содержит 2 молекулы апо(а) в дополнение к 1 моле-куле апоВ. Увеличенный уровень ЛП(а) ассоциирован с ранним развитием ИБС: 50% больных ИБС имели уровень ЛП(а) выше 40 мг/л по сравнению с 17% в контрольной группе. Увеличение концентрации ЛП(а) в крови считают независимым фактором риска атеросклероза.
ЛПВП образуются несколькими путями, включая синтез и секрецию насцентных ЛПВП печенью и кишечником, в процессе липолиза ЛП, богатых ТГ, или в результате взаимодействия новообразованных апопротеинов с мембранами клеток. Насцентные ЛПВП - бислойные диски, состоящие из апоЛП и фосфолипидов. Свободный холестерин диффундирует в ЛПВП из клеток эндотелия, эритроцитов и других клеток. Кроме того, во время липолиза ЛПОНП или ХМ поверхностные липиды (фосфолипиды и холестерин) и белки (апоА-1, апоА-П, апопротеины группы С) переносятся во фракцию ЛПВП. Эти компоненты могут сформировать насцентные дискоидальные частицы ЛПВП или быть включены в уже существующие частицы ЛПВП. Дис-коидальные насцентные ЛПВП в плазме крови взаимодействуют с ферментом ЛХАТ, в результате образуются эфиры холестерина, быстро перемещающиеся в ядро частицы; формируются сферические (зрелые) частицы ЛПВП - ЛПВП3.
У человека большая часть эфиров холестерина, образованных в ЛПВП, переносится в более крупные ЛП, богатые ТГ. Белок,
переносящий эфиры холестерина, способствует обмену эфиров холестерина из ЛПВП на ТГ из ЛП, содержащих апоВ, что приводит к снижению содержания эфиров холестерина и уве-личению содержания ТГ в ЛПВП. Комбинированный эффект липолиза и переноса липидов приводит к обогащению частиц ЛПВП дополнительными липидами и апопротеинами, в результате чего происходят увеличение размера и снижение плотности частиц ЛПВП, сопровождающиеся превращением частиц ЛПВПз в ЛПВПг- Частицы ЛПВП2затем подвергаются воздействию триглицеридлипазы печени - фермента, обнаруженного в основном в эндотелиальных клетках синусоидов печени. Триг- лицеридлипаза печени обладает как триглицеридгидролазной, так и фосфолипазной активностью, что приводит к снижению содержания ТГ и фосфолипидов в ЛПВП и уменьшению размера частицы.
Таким образом, ЛП плазмы крови, традиционно разделяемые на классы физическими, химическими и иммунохимическими методами, метаболически составляют единую систему.
Содержание классов ЛП в сыворотке крови можно измерить количественно. Такие измерения проводят методом аналитического ультрацентрифугирования, который требует сложного аналитического оборудования. Возможно также рассчитать концентрацию ЛП, исходя из анализа каждого компонента. Этот подход требует количественного выделения и очистки каждого класса ЛП методом препаративного ультрацентрифугирования. Уровень ЛП в плазме крови в клинической биохимии обычно оценивают по содержанию в них холестерина. Содержание ТГ в отдельных классах ЛП, как правило, не исследуют, поскольку оно подвержено более значительным колебаниям, чем уровень холестерина. Следует подчеркнуть, что большая часть эпидемиологических данных о связи ЛП и развития заболеваний ССС основана на измерениях именно содержания холестерина в отдельных классах ЛП.
Соотношение общего холестерина плазмы крови и холестерина основных классов ЛП можно выразить следующей формулой:
холестерин крови = холестерин ЛПОНП +
+ холестерин ЛПНП {- холестерин ЛПВП.
Для измерения холестерина классов ЛП определяют общий холестерин и ТГ сыворотки крови и концентрацию холестерина ЛПВП в супернатанте после преципитации ЛП, содержащих апоВ (ЛПОНП и ЛПНП). Концентрацию холестерина ЛПНП рассчитывают по формуле Фридвальда:
холестерин ЛПНП = общий холестерин -
- холестерин ЛПВП - холестерин ЛПОНП.
Для ориентировочной оценки концентрации холестерина ЛПОНП используют величину ТГ/5, если концентрации холестерина и ТГ выражены в мг/дл, или ТГ/2,2, если концентрации холестерина и ТГ выражены в ммоль/л. Применение этой формулы при концентрации ТГ выше 400 мг/дл приводит к завышению содержания холестерина ЛПОНП и занижению содержания холестерина ЛПНП. Для оценки распределения ЛП у больных с очень высоким уровнем ТГ и наличием значительного количества ХМ необходимо предварительное ультрацентрифугирование для отделения ХМ и ЛПОНП. После отделения фракции ЛПОНП и преципитации ЛПНП в супернатанте можно определить концентрацию холестерина ЛПВП.
Достоверный метод оценки содержания отдельных классов ЛП в сыворотке крови - метод ЭФ на одной из поддерживающих сред: геле агарозы, геле полиакриламида, ацетате целлюлозы, бумаге. Эти методы позволяют получить только процентное соотношение отдельных классов ЛП, включая ХМ, |3-, пре-ф- и а-ЛП. ЭФ - основа фенотипирования ГЛП по классификации, предложенной Фредриксоном (табл. 1.10). Окончательное решение о фенотипе ГЛП проводят по данным ЭФ ЛП с учетом концентрации в крови холестерина и ТГ, а также холестерина ЛПНП.
Клиническое значение определения липопротеинов в крови
Многочисленными клиническими и эпидемиологическими исследованиями установлено, что повышение уровня холестерина в крови - один из основных факторов риска атеросклероза. Раз-
Таблица 1.10. Классификация гиперлипопротеинемий, принятая ВОЗ Тип ХМ лпонп лпнп Холестерин ТГ Характер
нарушений I т N N N ТТ Т хм Па - N ТТ ТТ N т лпнп Пь - т т т т Т лпнп и лпонп III - Флотирующие р-ЛП т т т ОКХ и ЛППП IV - т N N (Т) т Т лпонп V т 1 N N (Т) тт Т хм и лпонп Примечание. | - повышение; ОКХ - остаточные компоненты ХМ.
витие ГЛП может быть обусловлено генетическими аномалиями и факторами среды (первичные ГЛП), а также такими заболеваниями, как сахарный диабет, патология печени, почек, гормональными нарушениями (вторичные ГЛП). В формировании первичных ГЛП основную роль играет наследственная предрасположенность. В популяции белых американцев вариабельность уровня холестерина на 63% обусловлена генетической изменчивостью. По данным обследования моно- и дизиготных близнецов в России, изменчивость общего холестерина на 82% обусловлена генетическими факторами.
В настоящее время изучено много наследственных аномалий обмена ЛП, но только для некоторых известны точные биохимические дефекты, позволяющие диагностировать заболевание. К таким формам ГЛП относят семейную дисбеталипопротеине- мию (ГЛП типа III) и семейную гиперхиломикронемию (ГЛП типа I).
СЕМЕЙНАЯ ГИПЕРХОЛЕСТЕРИНЕМИЯ
Среди пациентов с первичной гиперхолестеринемией наиболее изучены нарушения обмена липидов при семейной гиперхо- лестеринемии. Это наследственное моногенное заболевание, при котором отмечают высокий уровень холестерина в сыворотке крови и ЛПНП, а также раннее развитие атеросклероза и ИБС.
Хип наследования при семейной гиперхолестеринемии - ауто- сомно-доминантный. У гомозигот заболевание протекает более тяжело, чем у гетерозигот. У 60% гомозигот ИБС развивается в возрасте до 10 лет. Уровень холестерина сыворотки крови у гетерозигот варьирует от 250 до 550 мг/дл, у гомозигот отмечено повышение до 800 мг/дл. Ксантомы сухожилий считают патогномоничными для больных с семейной гиперхолестерине- мией. Частота распространения семейной гиперхолестеринемии среди популяций Европы и Америки составляет 1: 1 000000 для гомозигот и 1: 500 для гетерозигот, хотя существуют популяции с более высокой частотой проявления заболевания - 1: 100 (канадские французы, финны, европеоиды в ЮАР).
Молекулярный дефект, определяющий развитие семейной гиперхолестеринемии, был описан Брауном и Гольдштейном. Причина гиперхолестеринемии - дефект ЛПНП-рецептора, вызывающий резкое снижение поглощения ЛПНП, вследствие чего происходят накопление частиц ЛПНП и соответственно возрастание в крови их концентрации. Степень снижения активности ЛПНП-рецептора у пациентов с семейной гиперхолестеринемией коррелирует с уровнем холестерина ЛПНП, но не с клиническими проявлениями заболевания (ранние проявления ИБС, наличие сухожильных ксантом). Установлено 4 типа генетических дефектов ЛПНП-рецептора:
полное отсутствие белка-рецептора;
нарушение транспорта белка-рецептора к поверхности клетки;
дефект рецептора, препятствующий связыванию ЛПНП;
дефект рецептора, препятствующий его интернализации после связывания с ЛПНП. В настоящее время выявлено более 150 мутаций этого белка.
Несмотря на установление генетического дефекта, определяющего развитие семейной гиперхолестеринемии, характеристику клинических проявлений заболевания и нарушений липидного обмена, критерии диагностики семейной гиперхолестеринемии окончательно не определены. К сожалению, определение активности ЛПНП-рецептора для диагностики семейной гиперхолестеринемии не нашло широкого распространения. Полагают, что использование анализа ДНК для диагностики семейной гиперхо- лестеринемии нецелесообразно вследствие большого количества мутаций. Увеличение уровня холестерина - нечеткий диагностический критерий семейной гиперхолестеринемии, поскольку описаны пациенты, имеющие сниженную активность апоВ-ре- цептора и нормальный уровень холестерина.
Снижение связывания частиц ЛПНП с рецептором может быть обусловлено не только генетическими дефектами, вызывающими отсутствие ЛПНП-рецептора или его функциональные нарушения, но и наследственными изменениями лиганда - апоВ-100. Анализ ДНК пациентов с гиперхолестеринемией позволил выявить мутацию гена, кодирующего синтез апоВ-100, в результате которой аргинин в положении 3500 заменен глутамином. Эта патология получила название «наследственный дефект апоВ-100». Частота распространения этого дефекта в популяции сопоставима с таковой для семейной гиперхолестеринемии (1: 500), по другим данным значительно ниже (1: 1300).
СЕМЕЙНАЯ КОМБИНИРОВАННАЯ ГИПЕРЛИПИДЕМИЯ Термин «семейная комбинированная гиперлипидемия» (СКГЛ) впервые предложен Гольдштейном и соавт. в 1973 г. для характеристики наследственного нарушения липидного обмена, при котором у пробанда и его родственников I степени родства отмечают разные типы ГЛП (На, Нь, IV). Нарушения липидного обмена проявляются обычно после 20 лет, но могут быть выявлены и в более раннем возрасте. Частота встречаемости СКГЛ в популяции составляет, по данным разных исследователей, от 0,5 до 2%, тогда как среди больных инфарктом миокарда - 5%, а среди пациентов в возрасте до 60 лет с ангиографически документированной ИБС - 15-20%. Изучение родословных показало, что СКГЛ - наследственное заболевание с аутосомно-доминантным типом наследования. Несмотря на многочисленные исследования, биохимический маркер СКГЛ не определен; точный молекулярный дефект, приводящий к формированию СКГЛ, пока неизвестен.
ПОЛИГЕННАЯ ГИПЕРХОЛЕСТЕРИНЕМИЯ Среди наследственных нарушений липидного обмена наиболее распространена полигенная гиперхолестеринемия. При этом заболевании распределение уровней холестерина у членов таких
семей сдвинуто в сторону более высоких значений, чем в среднем в популяции. Кроме того, распределение унимодально, что указывает на отсутствие моногенного наследования. Полагают, что наличие полигенной гиперхолестеринемии обусловлено суммарным влиянием нескольких генов, при этом проявление полигенной гиперхолестеринемии в большой степени провоцируют сре- довые факторы, особенно характер питания. Термин «полигенная гиперхолестеринемия» применяют для характеристики пациентов с наследственной гиперхолестеринемией с неустановленным генетическим дефектом и при отсутствии признаков моногенного наследования.
Учитывая полигенный характер наследования гиперхолестеринемии, генетический дефект, определяющий заболевание, и его биохимический маркер (или маркеры) неизвестны. Рас-пространенность полигенной гиперхолестеринемии в популяции и среди больных ИБС зависит от значений уровня холестерина, принятых в качестве нормальных: при снижении нормы доля пациентов с полигенной гиперхолестеринемией, естественно, увеличивается.
СЕМЕЙНАЯ ГИПЕРТРИГЛИЦЕРИДЕМИЯ
Семейная гипертриглицеридемия - наследственное нарушение липидного обмена, при котором у пробанда и его родственников выявляют умеренное или значительное увеличение уровня ТГ при нормальной концентрации холестерина ЛПНП - ГЛП IV или V типа. Заболевание обычно проявляется после 30 лет. У таких пациентов часто, но не всегда отмечают снижение уровня холестерина ЛПВП. Тип наследования - аутосомно-доминант- ный. Семейная гипертриглицеридемия встречается в популяции с частотой 1%, а у больных ИБС в возрасте до 60 лет с частотой 5-8%. Генетический дефект, обусловливающий развитие семейной гипертриглицеридемии, и биохимический маркер этого заболевания в настоящее время неизвестны.
Диагностические критерии гиперлипопротеинемий
Для установления ГЛП необходимы четкие биохимические критерии. В настоящее время есть два подхода к этому вопросу. Долгие годы нормальный уровень холестерина и ТГ сыворотки крови, а также холестерина ЛПНП и ЛПВП устанавливали по результатам эпидемиологических исследований. Для каждой популяции определяли свои критерии нормы показателей липидного обмена в зависимости от 5-10% минимальных и максималь-ных значений при гауссовском распределении. Гиперлипидемией в этом случае считали такую концентрацию холестерина и ТГ, которая превышала 90 или 95% уровень на графике Гаусса (в соответствии с полом и возрастом). Именно эти критерии значений нормы положены в основу классификации ГЛП, предложенной Фредриксоном и соавт. и после некоторых дополнений принятой ВОЗ. Впоследствии, учитывая накопленные данные о взаимосвязи повышенного уровня холестерина и риска ИБС, экспертными группами Национальной образовательной программы по холестерину в США и Европейского общества по изучению атеросклероза предложена иная классификация ГЛП. В зависимости от риска ИБС предложено выделять желаемый уровень холестерина в крови 6,2 или > 6,5 ммоль/л соответственно для мужчин и женщин старше 20 лет. На основании названных критериев предложено выделять умеренную гиперхолестеринемию (6,2-7,5 или 6,5-7,8 ммоль/л) и тяжелую гиперхолестеринемию (> 7,5 или > 7,8 ммоль/л).
Соответственно изменены критерии и для холестерина ЛПНП, тогда как уровень нормы для ТГ и холестерина ЛПВП оставлен прежним. Хотя для американской популяции умеренная гиперхолестеринемия соответствует 75-95% уровню гауссовского распределения, эти критерии используют для типирования ГЛП в США и России. Выбор более низких значений нормы, несомненно, оправдан с точки зрения первичной и вторичной профилактики ИБС. Кроме того, применение единых критериев для диагностики позволяет сопоставить распространенность ГЛП в популяциях с различным средним уровнем холестерина. С другой стороны, 95% лиц с ГЛП, установленной по эпидемиологическим критериям, имеют наследственно обусловленную ГЛП. Соответственно по мере снижения нормальных значений будет возрастать доля лиц, ГЛП которых обусловлена только факторами среды. В этой связи при поиске пациентов с тяжелыми наследственными формами гиперхолестеринемии
по-прежнему применяют эпидемиологически установленный 90- 95% уровень холестерина в соответствии с полом и возрастом
пациента.
Гиполипопротеинемии
Гиполипопротеинемии возникают в результате некоторых дефектов, регулирующих липидный метаболизм ферментов либо мутаций гена апоВ.
Абеталипопротеинемия и гипобеталипопротеинемия - наследуемые расстройства, вызванные мутациями гена апоЛП В, главного апоЛП ХМ и ЛПОНП. Специфическая терапия этих расстройств не разработана.
Абеталипопротеинемия: в крови отсутствуют р-липопроте- ины, ХМ, ЛП с плотностью ниже 1,063 (ЛПНП и ЛПОНП), эритроциты имеют множественные шиповидные выросты (акан- тоциты), развивается дефицит витаминов Е и А (результат отсутствия ЛПНП, транспортирующих жирорастворимые витамины). Часты нарушения всасывания в кишечнике, а также нарушения координации (вследствие демиелинизации аксонов), атаксия, нистагм, пигментная дегенерация сетчатки и отставание в умственном развитии. Стеаторея возникает вследствие недостатка апоВ, определяющего образование ХМ в клетках кишечника. При биопсии тонкой кишки обнаруживают жировые включения в эпителиальных клетках.
Гипобеталипопротеинемия сопровождается уменьшением содержания р-липопротеинов и часто протекает бессимптомно. Содержание ЛПВП или в норме или увеличено, триглицеридов 70-120 мг%, ЛПНП 20-70 мг%. Всасывание жиров обычно не нарушено.
ТРИГЛИЦЕРИДЫ
Триглицериды (ТГ; триацилглицерины) - эфиры трехатомного спирта глицерина и длинноцепочечных жирных кислот, среди которых в организме человека доминируют стеариновая, пальмитиновая, пальмитоолеиновая, олеиновая, линолевая кислоты, Реже присутствуют полиненасыщенные линоленовая и арахидо- новая кислоты. Клетки тканей человека не синтезируют лино- левую кислоту, поэтому ее относят к эссенциальным кислотам. Распределение жирных кислот в молекуле ТГ зависит от длины цепи и степени ненасыщенное™. В средней позиции чаще находится кислота с наибольшей длиной цепи и наибольшим числом двойных связей, крайние позиции обычно занимают насыщенные кислоты.
Жирные кислоты поступают в организм с пищей в форме триацилглицеринов, которые в тонкой кишке под действием липазы подвергаются гидролизу. Образовавшиеся глицерин и свободные жирные кислоты абсорбируются клетками эпителия, где происходит ресинтез ТГ. Формирование ТГ происходит в печени; жирные кислоты поступают в печень из жировой ткани или синтезируются в гепатоцитах. Жирнокислотный состав ТГ в организме в большей степени, чем фосфолипидов, зависит от состава жиров пищи. Длинноцепочечные жирные кислоты, входящие в состав ТГ, - оптимальный источник макроэргиче- ских соединений, образуемых при р-окислении жирных кислот в митохондриях. Большинство органов реализует именно ТГ для покрытия энергетических потребностей. Исключение составляет мозг; его энергетическое обеспечение происходит за счет метаболизма глюкозы (аэробного гликолиза).
В плазме крови ТГ, как и другие липиды (холестерин и его эфиры, фосфолипиды), транспортируются в составе макромоле- кулярных мицеллярных комплексов - ЛП. ТГ присутствуют во всех классах ЛП, в ЛПОНП и ХМ они составляют большую часть липидов. При определенном опыте внешний вид плазмы позволяет ориентировочно определить содержание в ней ТГ. Если плазма крови прозрачна, уровень ТГ в ней не превышает 180-200 мг%; выраженная опалесценция плазмы крови характерна для гипертриглицеридемии порядка 300-500 мг%, при уровне ТГ, превышающем 600 мг%, плазма и сыворотка крови непрозрачны. Наличие ХМ в крови можно выявить при проведении «теста стояния» - появление «крема» на поверхности плазмы при ее хранении в течение 16 ч при температуре 4 °С. Прозрачная сыворотка крови и наличие ХМ свидетельствуют о I типе ГЛП, отсутствие ХМ при выраженной опалесценции или молочном цвете сыворотки указывает на IV тип ГЛП; присутствие ХМ на поверхности образца сыворотки с выраженной опалесценцией или молочного цвета позволяет предположить у пациента III или V тип ГЛП.
Клиническое значение определения концентрации ТГ
Необходимо помнить, что ХМ могут присутствовать в сыворотке крови здорового человека, принимавшего пищу менее чем за 12 ч до взятия крови. В силу значительного влияния на уровень ТГ субстратной индукции уровень ТГ в крови может изменяться в течение суток в значительных пределах. Гипертриглице- ридемия может быть охарактеризована как физиологическая или патологическая. Физиологическая гипертриглицеридемия возникает после приема пищи и может продолжаться 12-14 ч; степень ее проявления зависит от характера и количества принятой пищи. Гипертригл ицеридемия во Н-Ш триместре беременности также физиологическая. Патологическая гипертриглицеридемия по своим патогенетическим характеристикам может быть дифференцирована на первичную и вторичную. Первичная гипертриглицеридемия обусловлена генетическими нарушениями метаболизма ЛП или усиленной субстратной индукцией (переедание). Нарушения обмена ТГ, возникающие как осложнение основного патологического процесса, относят к вторичным гипертриглице- ридемиям.
Известно несколько первичных форм гипертриглицеридемий, обусловленных наследственными дефектами метаболизма ЛП: семейная гипертриглицеридемия, семейная комбинированная гиперлипидемия, семейная дисбеталипопротеинемия (тип III ГЛП), семейная гиперхиломикронемия (тип I ГЛП). К первичным формам нарушения обмена ТГ могут быть отнесены и гипотригли- церидемии, вызванные генетическими изменениями синтеза или секреции апоВ. При этих заболеваниях (наследственной абета- липопротеинемии или гомозиготной гипобеталипопротеинемии) содержание триглицеридов может быть снижено до 19-20 мг%.
Развитие вторичных гипертриглицеридемий отмечено при сахарном диабете, гипотиреозе, нефротическом синдроме, приеме пероральных контрацептивов, лечении глюкокортикоидами. Как правило, патогенетическое лечение, направленное на основной патологический процесс, сопровождается снижением уровня ТГ.
Коррекция уровня гликемии инсулином или пероральными гипо- гликемическими препаратами способствует нормализации содержания ТГ в крови. При купировании нефротического синдрома уровень ТГ в крови также снижается, но иногда остается повышенным в течение длительного времени. Прекращение приема гормональных контрацептивов, глюкокортикоидов и ряда других препаратов (р-блокаторы) также способствует нормализации уровня ТГ.
Гипертриглицеридемию отмечают при жировой инфильтрации печени. В основе патогенеза этого заболевания лежит повышенное поступление свободных жирных кислот в печень (усиленный липолиз) или их усиленный синтез (ресинтез), с одной стороны, и нарушение секреции ЛПОНП печенью - с другой. Многие токсичные соединения, алкоголь, фармакологические препараты способствуют нарушению синтеза апоЛП, в частности апоВ, в отсутствие или при недостатке которых не происходит формирования ЛПОНП. В этих условиях поступающие из крови или вновь синтезируемые ТГ оказываются в печени, формируя структуру «свиной печени», вызывая ее жировую инфильтрацию и цирроз.
Гипертриглицеридемию обычно отмечают у пациентов с острым панкреатитом. Однако увеличение уровня ТГ нехарактерно для острого поражения поджелудочной железы, напротив, выраженная гипертриглицеридемия (>1000 мг%), как первичная, так и вторичная, способствует развитию заболеваний поджелу-дочной железы и зачастую осложняется панкреатитом. Панкреатит считают одним из клинических проявлений семейных форм гипертриглицеридемии. Развитие панкреатита часто отмечают у больных сахарным диабетом, при алкоголизме, у женщин, принимавших гормональные контрацептивы.
Данные о взаимосвязи гипертриглицеридемии и ИБС противоречивы, хотя эпидемиологическими исследованиями на многих популяциях показана независимость ТГ как фактора риска ИБС. Оценка независимости ТГ как фактора риска ИБС крайне затруднена вследствие наличия отрицательной зависимости между уровнем ТГ и холестерина ЛПВП, независимого отрицательного фактора риска ИБС. Тем не менее если повышение концентрации ТГ и не является независимым фактором риска ИБС, тесная связь метаболизма ТГ и ЛПВП не дает оснований отрицать значение гипертриглицеридемии в генезе ИБС. Биохимической основой этого может быть накопление в крови ЛППП, обладающих выраженной атерогенностью.
Значение гипертриглицеридемии в формировании патологии периферических и церебральных сосудов в отличие от ИБС более определенно. Исследования японских специалистов показали, что в этой популяции при низком уровне холестерина крови и частоты возникновения инфаркта миокарда гипертриглице-ридемия - фактор риска патологии периферических артерий. Аналогичные данные получены и на других популяциях.
Исследование содержания ТГ в сыворотке крови имеет много общего с определением холестерина. Как холестерин, так и ТГ - структурные компоненты ЛП, которые необходимо разрушить, чтобы сделать липиды доступными для химических или ферментативных реакций. Оба соединения гидрофобны, обладают низкой растворимостью в водных растворах. Однако в определении содержания в сыворотке крови ТГ имеются и существенные отличия: для оценки содержания ТГ определяют глицерин, освободившийся после гидролиза ТГ.
Методы определения содержания ТГ в крови
Существуют два способа определения содержания ТГ в сыворотке крови - химический и ферментативный; каждый из них включает многочисленные методические приемы. В настоящее время определение ТГ возможно также методом «сухой химии».
Химические методы. Для определения ТГ прямые химические методы (без предварительной экстракции органическим растворителем) неприемлемы, так как в сыворотке крови присутствуют многие соединения, которые в процессе химического гидролиза (омыления жирных кислот) также могут превращаться в глицерин, например фосфолипиды и глюкоза. В этой связи в химических методах определения ТГ первая стадия - экстракция ТГ и удаление интерферирующих соединений. Для экстракции используют органические растворители: метанол, эта-нол, изопропанол и хлороформ. Растворители денатурируют ЛП и способствуют диссоциации ТГ. Интерферирующие соединения удаляют экстракцией органическими растворителями (гексан, нонан) или при использовании адсорбентов (цеолит, кремниевая кислота, флоризил).
Вторая стадия химического определения ТГ - гидролиз ТГ с образованием глицерина и свободных жирных кислот - обычно осуществляется реакцией с этанольным раствором гидроокиси калия при повышенной температуре (омыление).
После окисления глицерина в формальдегид его определяют одной из перечисленных ниже реакций:
реакцией формальдегида со смесью хромотроповой и серной кислот (измерение при 570 нм);
реакцией со смесью фенилгидразина, цианида железа и соляной кислоты (измерение при 540 нм);
реакцией со смесью З-метил-2-бензотиазолина и хлорида железа (измерение при 620 нм);
реакцией с ацетатом аммония и ацетилацетоном.
Продукт реакции может быть измерен колориметрически при
412 нм или флюорометрически. Более распространен флюоро- метрический метод, он адаптирован к полуавтоматическим ана-лизаторам.
Ферментативные методы. При ферментативных методах определения ТГ разрушение ЛП-комплексов, гидролиз ТГ проводят в присутствии детергентов, липазы и протеазы. В условиях ферментативного гидролиза ТГ не происходит освобождения глицерина из фосфолипидов и глюкозы, следовательно, повышается специфичность определения.
Для определения глицерина, образованного при гидролизе ТГ, применяют множество методических приемов, используя в каждом случае ряд сопряженных ферментативных реакций. Ферментативные методы позволяют определять ТГ по уменьшению или увеличению концентрации НАДН при длине волны 340 нм. Предложены и колориметрические методы: восстановление солей тетразолия нитросинего при помощи НАДН и измерение образованного формазана (длина волны 505 нм) или окисление 4-аминофеназона в присутствии пероксидазы с образованием хромогена (длина волны 510 нм).
Использование ферментативного метода позволяет избежать применения органических растворителей, очистки экстрактов для удаления интерферирующих соединений, использования концентрированной гидроокиси калия и проведения гидролиза при высокой температуре. По сравнению с химическими методами ферментативное определение ТГ обладает более высокой специфичностью и точностью, что удобно для автоматизации.
Основные сложности ферментативного определения ТГ - возможность неполной деструкции комплекса ЛП и неполного гидролиза с освобождением свободного глицерина. Из экспериментальных работ известно, что ТГ, образованные насыщенными и ненасыщенными жирными кислотами, с разной скоростью гидролизируются при действии липазы; соотношение коротко- и длинноцепочечных жирных кислот также имеет значение для скорости гидролиза.
Поскольку определение содержания ТГ в сыворотке крови как в химических, так и ферментативных методах основано на освобождении глицерина, необходимо принимать во внимание содержание свободного глицерина в сыворотке крови. Для большинства свежих образцов сыворотки крови при уровне ТГ ниже 700 мг% изменение свободного глицерина не оказывает существенного влияния на получаемые результаты. Накопление свободного глицерина в сыворотке крови происходит при сахарном диабете, некоторых видах патологии печени, в условиях гемодиализа, при острой и хронической почечной недостаточности, приеме нитроглицерина, терапии гепарином. Острые инфекции, кратковременные состояния стресса, прием лекарственных препаратов, обладающих липолитическим свойством, значительно увеличивают содержание свободного глицерина в крови. Спонтанный гидролиз ТГ происходит и в сыворотке крови, особенно при хранении образцов при комнатной температуре.
Для точности определения уровня ТГ следует измерить уровень свободного глицерина и результат вычесть из общего значения. При определении уровня свободного глицерина колориметрическим и флюорометрическим методами он может быть разным в одной и той же пробе плазмы крови. Флюорометрия Дает более стабильные результаты, но менее специфична: в усло-виях почечной недостаточности и активированного протеолиза флюоресцирующие пептиды могут значительно завышать результаты.
Для получения точных данных при определении ТГ необходимо использовать первичный стандарт - раствор триацил- глицеринов в органических растворителях. В качестве стандарта обычно применяют триолеин, хотя смесь триолеина и трипаль- митина (2:1 по массе) более сходна по степени ненасыщенности жирных кислот с ТГ человека. В качестве стандарта раствор триолеина в изопропаноле можно применять только для химических методов исследования, так как изопропанол неспецифически ак-тивирует ферменты, используемые для определения ТГ. В этой связи при определении ТГ ферментативными методами используют вторичные стандарты (калибраторы) - сыворотки крови человека, содержание ТГ в которых определено референсными методами, с применением первичных стандартов.
Взятие крови для определения ТГ необходимо проводить через 12-14 ч после приема пищи. Значения ТГ, определенные в плазме крови, на 2-4% ниже таковых в сыворотке. Это обусловлено разведением за счет жидкости, теряемой эритроцитами под действием антикоагулянта. Некоторые антикоагулянты (фторид, цитрат и оксалат) вызывают более значительную потерю жидкости эритроцитами, что находит отражение в большем снижении значений ТГ (до 10%). В качестве антикоагулянта при определении ТГ предпочтительнее использовать ЭДТА (1 мг/мл цельной крови). Для уменьшения эффекта разбавления необходимо центрифугировать образцы крови и отделить полученную плазму в течение 2 ч после взятия крови.
ХОЛЕСТЕРИН
Холестерин, З-р-гидроксихолест-5-ен, - мононенасыщенный стерин состава С27Н40(0Н)б; по химической структуре это од-ноатомный вторичный спирт (холестерол). Молекула холестерина имеет в основе структуру циклопентанпергидрофенантрена, в третьем положении расположена гидроксильная группа, присутствуют двойная связь между 5-м и 6-м углеродными атомами и боковая алифатическая цепь из 8 атомов углерода. Термин «холестерин» определяет источник, из которого он впервые был выделен (от греч. сЬо1е - желчь). Молекулярная масса холестерина 387 Д, в чистом виде это порошок белого цвета. Холестерин
хорошо растворяется в органических растворителях (особенно хорошо в спиртах с 6-7 атомами углерода). Растворимость холестерина в воде крайне низкая, составляет 4,7 мкмоль/л при 25 °С.
Химическая структура холестерина ограничивает число биохимических реакций, в которые он вступает. Основное биохимическое превращение холестерина (как спирта) - образование эфирной связи с кислотами, в частности со свободными жирными кислотами, с формированием эфиров холестерина. 80% холестерина в организме составляет свободный холестерин, почти весь он входит в состав биологических мембран. В то же время в сыворотке крови 2/3 холестерина представлено в форме эфиров с жирными кислотами.
Холестерин входит в состав всех живых клеток. В биологических мембранах холестерин выполняет структурную роль. Со структурой неразрывно связана и его функция, определяющая проницаемость мембраны и создание микроокружения для встроенных в мембрану ферментов. Важная функция холестерина - его роль как предшественника синтеза стероидных гормонов.
Все клетки организма синтезируют холестерин из ацетата, последовательно проходя стадии ацетоацетата, метилглута- рил-КоА и, наконец, мевалоновой кислоты. Превращение метил- глутарил-КоА в мевалоновую кислоту при действии гидрокси- метилглутарил-КоА-(ГМГ-КоА)-редуктазы - ключевой момент синтеза холестерина. Образованная мевалоновая кислота может быть использована только в синтезе холестерина, поэтому активация ГМГ-КоА-редуктазы всегда приводит к гиперхолестеринемии.
Основная масса холестерина синтезируется в печени и поступает с пищей; в сутки организм синтезирует 1 г холестерина. Часть холестерина окисляется в желчные кислоты, часть удаляется с калом. Существуют реципрокные отношения между холе-стерином, поступающим с пищей, и синтезируемым в организме. Холестерин, как и другие липиды, транспортируется в сыворотке крови в составе ЛП.
Основную массу холестерина сыворотки крови содержат ЛПНП; их основной апоЛП - апоВ-100. ЛПНП - основная форма транспорта холестерина и полиеновых жирных кислот от
Клиническая биохимия печени к периферическим тканям. ЛПНП при взаимодействии с апоВ-рецепторами плазматической мембраны подвергаются интернализации; в липосомах происходит деструкция ЛП-частицы с освобождением свободного холестерина. Клетка использует холестерин для построения мембран, часть его этерифицирует- ся. Молекулы холестерина плазматических мембран, оказавшись в сыворотке крови, начинают афферентный путь к печени в составе ЛПВП. Поступивший в печень в составе ЛПВП холестерин подвергается микросомальному гидроксилированию, превращаясь в желчные кислоты, а также экскретируется с желчью в форме свободного холестерина или его эфиров.
На уровень холестерина влияют характер пищи и прием алкоголя, интенсивные физические тренировки, фармакологические препараты, включая гормональные контрацептивы, стероиды, гиполипидемические препараты. Сезонные и дневные вариации не оказывают существенного влияния на уровень холестерина в сыворотке крови. Индивидуальные колебания значений холестерина составляют, по данным разных авторов, от 3 до 14%. Суточные колебания уровня холестерина у мужчин и женщин (0,7-4,3%) значительно ниже, чем колебания в течение
мес (2,0-12,2%) и 1 года (3,9-10,9%).
Клиническое значение гиперхолестеринемии
Гиперхолестеринемия - наиболее документированный фактор риска коронарного атеросклероза. Это подтверждено многочисленными эпидемиологическими и клиническими исследованиями, установившими связь гиперхолестеринемии с коронарным атеросклерозом, частотой клинических проявлений ИБС (стенокардии и инфаркта миокарда). При вторичной профилактике коронарного атеросклероза в ходе гиполипидемической терапии продемонстрирована регрессия клинической картины ИБС и кардиосклероза при нормализации уровня холестерина сыворотки крови. Для более детальной характеристики факторов риска коронарного атеросклероза исследуют содержание холестерина в отдельных классах ЛП; наибольшее распространение получило исследование холестерина ЛПНП и ЛПВП, являющихся достоверными факторами соответственно риска и антириска коронарного атеросклероза.
Исследование холестерина не несет диагностической информации относительно определенного заболевания, а характеризует патологию обмена липидов и ЛП. Определение уровня холестерина имеет социальное значение, так как повышенный популя-ционный уровень холестерина требует осуществления организационных мероприятий по первичной профилактике коронарного атеросклероза. Наиболее высокие цифры гиперхолестеринемии отмечаются при генетических нарушениях обмена ЛП: семейной гомо- и гетерозиготной гиперхолестеринемии, семейной комбинированной гиперлипидемии, полигенной гиперхолестеринемии.
При ряде патологических состояний развивается вторичная гиперхолестеринемия. К таким процессам относят гипотиреоз, нефротический синдром, сахарный диабет и др. Гипохолестери- немия возникает при гипертиреозе, при тяжелых клинических ситуациях, сопровождающих разные нозологические формы заболеваний, при острой стадии воспалительного процесса, а также онкологических заболеваниях.
Методы определения холестерина в сыворотке
Методы определения холестерина в сыворотке крови многочисленны; можно выделить химические и ферментативные методы. В свою очередь среди химических методов выделяют прямые и непрямые (экстракционные) методы.
ХИМИЧЕСКИЕ МЕТОДЫ ОПРЕДЕЛЕНИЯ КОНЦЕНТРАЦИИ ХОЛЕСТЕРИНА
Прямые методы. Основа прямых методов определения холестерина - реакция Либермана-Бурхарда, в которой холестерин взаимодействует со смесью серной и уксусной кислот, а также уксусного ангидрида. Реакция протекает в сильнокислой безводной среде. В ходе реакции молекула холестерина последовательно окисляется и образуется окрашенное соединение хо- лестагексаен - серная кислота с максимумами абсорбции при 410 и 610 нм. Развивающаяся окраска неустойчива, поэтому следует точно выдерживать время фотометрирования. В литературе можно встретить разное соотношение ингредиентов в реактиве Либермана-Бурхарда; чем выше содержание уксусного ангидрида, тем с большей скоростью протекает реакция. Реак
ция холестерина со смесью Либермана-Бурхарда неспецифична, В реакции прямого определения холестерина окраска реакционной смеси со стандартным раствором имеет изумрудный цвет; пробы сыворотки могут давать зеленый, голубой, бурый цвет. Это связано с тем, что в реакцию вступают многие компоненты сыворотки крови. Кроме того, в реакции Либермана-Бурхарда свободный холестерин и его эфиры формируют окрашенные комплексы с разным коэффициентом молярного поглощения: в случае эфиров холестерина оптическая плотность оказывается более высокой. Это изначально вносит ошибку в исследование, поскольку неизвестно отношение свободный холестерин/эфиры холестерина в каждой из проб.
Реакция Киллиани-Зака - другой распространенный метод определения холестерина (реакция холестерина с солями железа, уксусной и серной кислотами). Эта химическая реакция протекает в присутствии ионов трехвалентного железа с последовательным формированием в молекуле холестерина 2, 3 и 4 двойных сопряженных связей, при этом максимум поглощения образовавшихся соединений меняется от 420 до 563 нм. В реакции Киллиани-Зака свободный холестерин и его эфиры дают сходные цветные комплексы.
Непрямые методы. К непрямым методам относят приемы, когда липиды из сыворотки крови вначале экстрагируют органическими растворителями, а после упаривания проводят реакцию Либермана-Бурхарда. Такой метод более воспроизводим и точен, поскольку удается убрать интерферирующие вещества, остающиеся в водной фазе. Для экстракции используют системы этанол- диэтиловый эфир, этанол-ацетон, метанол-хлороформ; наиболее часто применяют гексан и изопропиловый спирт. Включение этапа экстракции повышает специфичность метода; результаты определения холестерина на 7% ниже, чем при прямом методе.
Референсные методы определения холестерина проходят 3 стадии; они включают экстракцию липидов в неполярный растворитель, омыление эфиров холестерина и повторную экстракцию холестерина из омыленной смеси. В качестве референсного метода наиболее предпочтителен метод Абелл-Кендалла, пред* ложенный в 1952 г.: липидный экстракт омыляют спиртовым раствором гидроокиси калия и экстрагируют петролейным эфи* ром. Реакцию Либермана-Бурхарда проводят после упаривания петролейного эфира.
Трудности приготовления стандартных растворов холестерина связаны с его гидрофобностью. Длительное время в качестве стандартного раствора применяли раствор холестерина в ледяной уксусной кислоте. Далее в качестве стандартного был предложен раствор холестерина в изопропаноле, однако, такой стандарт оказался неудобным при ферментативных методах определения холестерина, поскольку изопропанол активировал ферментные системы, завышая оптическую плотность. Для определения холестерина при работе с биохимическими анализаторами общепринято использование вторичных стандартов (калибраторов) - сывороток крови, уровень холестерина в которых определен ре- ференсными методами.
Несмотря на относительную простоту исполнения и дешевизну процедуры, химические методы токсичны, применение их на современных анализаторах связано с коррозией системы.
ФЕРМЕНТАТИВНЫЕ МЕТОДЫ ОПРЕДЕЛЕНИЯ КОНЦЕНТРАЦИИ
ХОЛЕСТЕРИНА
Преимущества ферментативного определения: проведение реакции в водной фазе; удобство автоматизации; высокая чувствительность и специфичность.
Ферментативное определение холестерина также включает несколько этапов: ферментативный гидролиз эфиров холестерина при действии холестеролэстеразы с образованием свободного холестерина и свободных жирных кислот; окисление холестерина кислородом, растворенным в реакционной среде при действии холестеролоксидазы с образованием холест-4-ен-З-ола и Н2О2. В наиболее широко используемых методах образующаяся перекись водорода окисляет некоторые соединения с образованием хромогенов, которые могут быть измерены фотометрически. К таким хромогенам могут быть отнесены 4-аминофеназон (максимум абсорбции 500-525 нм), а также образование гетероцикла Хенача в реакции с метанолом (максимум абсорбции 405 нм). Возможна регистрация образованной перекиси водорода в реакции с НАД-зависимыми дегидрогеназами и измерением оптической плотности при длине волны 340 нм.
Как и в химических методах, некоторые компоненты сыворотки крови холестерина могут оказывать влияние на результаты определения. Билирубин ингибирует реакцию пероксидазы с 4-аминоантипирином и фенолом. Восстановители сульфгид- рильных групп (глутатион, цистеин, дитиотреитол) также оказывают влияние на результаты реакции. Методы, основанные на разных ферментативных реакциях, могут давать различные результаты.
Методические трудности ферментативного определения холестерина связаны с гетерогенностью распределения холестерина между ЛП, а также с его этерификацией в крови. Полнота гидролиза эфиров холестерина зависит от источника выделения фермента. Микробиальная холестеролэстераза более активно гидролизует эфиры холестерина, образованные насыщенными жирными кислотами, тогда как панкреатическая - полиненасы- щенными жирными кислотами.
Взятие крови для определения всех липидов обычно проводят после 12-16 ч голодания. Концентрация холестерина в крови не меняется до и после приема пищи, однако выраженная опалесценция сыворотки крови, обусловленная наличием ХМ и ЛПОНП, может мешать определению. Желательно, чтобы в течение по меньшей мере 2 нед до взятия крови пациент не менял привычного питания. Для определения уровня холестерина можно использовать сыворотку или плазму крови. В качестве антикоагулянта для получения плазмы обычно используют сухую ЭДТА (1 мг/мл крови), которая обладает также антиоксидант- ными свойствами. Концентрация холестерина в сыворотке крови на 2-4% ниже, чем в плазме крови.
Липиды –– жироподобные вещества, входящие в состав всех живых клеток. Это это широкая группа органических соединений, включающая жирные кислоты, а также их производные - как по радикалу, так и по карбоксильной группе. Липиды играют очень важную роль в жизненных процессах.
Липиды выполняют самые разнообразные функции. Они входят в состав клеточных мембран, служат предшественниками стероидных гормонов, желчных кислот, простагландинов и фосфоинозитидов.
В крови содержатся отдельные компоненты липидов - насыщенные жирные кислоты (НЖК), мононенасыщенные жирные кислоты (МНЖК) и полиненасыщенные жирные кислоты (ПНЖК), триглицериды, холестерин , эфиры холестерина и фосфолипиды.
Липиды - основные компоненты биологических мембран, влияющие на проницаемость клеток и активность многих ферментов, участвуют в передаче нервного импульса, в мышечном сокращении, создании межклеточных контактов, в иммунохимических процессах.
Липиды характеризуются растворимостью в органических растворителях и, как правило, нерастворимых в воде; входят в состав всех живых клеток. Липиды образуют энергетический резерв и создают защитные водоотталкивающие и термоизоляционные покровы у животных и растений, а также защают различные органы от механических воздействий. (по материалам БСЭ, Википедия, СМТ).
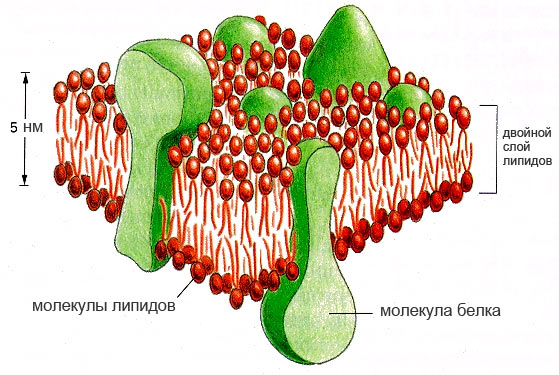
Мембрана клетки
Липиды транспортируются по кровеносному руслу к различным органам и тканям в составе сложных макромолекулярных соединений - липопротеидов , представляющих собой соединения различных белков (апобелков) и липидов.
Липопротеиды подразделяются на:
Липопротеиды различаются между собой размерами, составом и ролью в атерогенезе.
В клинической практике наибольшее значение в развитии сосудистой патологии играют ЛПНП и ЛПВП.
ЛПНП в основном состоят из эфиров холестерина. Их функциональным апопротеином является белок апоВ-100.
Для того чтобы ЛПНП стали атерогенными, они должны подвергнуться модификации, причиной которой чаще всего служит процесс их перекисного окисления. Окисленные ЛПНП изменяют свои свойства. Сначала нарушается их взаимодействие с рецепторами печени, потом они становятся активными раздражителями для моноцитов. Активированные моноциты крови проникают в субэндотелиальное пространство сосуда, превращаясь в макрофаги, которые фагоцитируют модифицированные ЛПНП и превращаются в пенистые клетки, т.е. клетки, переполненные эфирами холестерина. Именно холестерин-ЛПНП, учитывая его важную роль в формировании атеросклеротической бляшки, является главной мишенью гиполипидемической терапии.
ЛПВП осуществляют обратный транспорт холестерина из сосудистой стенки и макрофагов в печень, откуда они выводятся из организма в составе желчных кислот. Выделяют два подкласса ЛПВП: ЛПВП-2 и ЛПВП-3. Именно ЛПВП-3 осуществляют захват холестерина из периферических клеток и макрофагов, превращаясь в ЛПВП-2, которые состоят из эфиров холестерина и фосфолипидов. ЛПВП связываются срецепторами печени и клетками сосудистой стенки посредством апобелков (апоА-1 и апоА-2).
Липопротеиды низкой плотности – это соединения белков с холестерином, которые транспортируют последний из печени к тканям. ЛПНП переносят к тканям необходимый им холестерин, однако при повышении их концентрации, увеличивается риск развития атеросклероза.
Под атерогенными следует понимать липопротеиды, высокая коцентрация которых в плазме крови достоверно связана с развитием атеросклероза.
Макрофаги - крупные клетки, образовавшиеся из моноцитов, способные к фагоцитозу - процессу активного захватывания и поглощения живых и неживых частиц одноклеточными организмами или особыми клетками (фагоцитами) многоклеточных животных организмов.
Литература
Диагностика и коррекция нарушений липидного обмена с целью профилактики и лечения атеросклероза. Российские рекомендации. Разработаны Комитетом экспертов ВНОК. М., 2004.
Диагностика и коррекция нарушений липидного обмена с целью профилактики и лечения атеросклероза // Краткие Российские рекомендации. Разработаны группой экспертов ВНОК. М., 2005.
Орлов Р.С., Ноздрачёв А.Д. Нормальная физиология: Учебник. – М.: ГЭОТАР-Медиа, 2009.
Погожева А.В. Основы рациональной диетотерапии при сердечно-сосудистых заболеваниях // Клиническая диетология. 2004. Т.1. № 2.
Погожева А.В. Современные представления о диетотерапии артериальной гипертонии // Медицина. Качество жизни. 2005. № 3.
Рацион, питание и предупреждение хронических заболеваний. ВОЗ. Женева, 1993.
Реабилитация при заболеваниях сердечно-сосудистой системы / Под ред. И.Н. Макаровой. М., 2010.

