Рефлекс - ответная реакция организма на раздражение рецепторов, осуществляемая нервной системой.
Рефлекторная дуга - путь, по которому проходит нервный импульс во время осуществления рефлекса. Состоит из 5 частей:
- рецептор - чувствительное образование, способное реагировать на определенный вид раздражителя;
- чувствительный нейрон проводит импульсы в мозг;
- вставочный нейрон связывает чувствительные и исполнительные нейроны, находится в спинном или головном мозге;
- исполнительный (двигательный) нейрон проводит импульсы от мозга к исполнительным органам;
- исполнительный орган - мышца (сокращается), железа (выделяет секрет) и т.п.
Безусловные рефлексы
- имеются с рождения;
- в течение жизни не изменяются и не исчезают;
- одинаковы у всех организмов одного вида;
- приспосабливают организм к постоянным условиям.
Условные рефлексы
- приобретаются в течение жизни;
- могут изменяться или исчезать;
- у каждого организма свои собственные;
- приспосабливают организм изменяющимся условиям.
Тесты
1) Какое звено служит началом рефлекторной дуги
А) вставочный нейрон
Б) чувствительный нейрон
В) рецептор
Г) исполнительный нейрон
2. В какой последовательности компоненты рефлекторной дуги включаются в осуществление рефлекса
А) исполнительный орган, двигательный нейрон, вставочный нейрон, чувствительный нейрон, рецептор
Б) вставочный нейрон, чувствительный нейрон, двигательный нейрон, рецептор, исполнительный орган
В) рецептор, чувствительный нейрон, вставочный нейрон, двигательный нейрон, исполнительный орган
Г) чувствительный нейрон, вставочный нейрон, рецептор, исполнительный орган, двигательный нейрон
3. Начальный этап рефлекторной дуги в слюноотделительном рефлексе
А) слюнная железа
Б) рецептор
В) вставочный нейрон
Г) исполнительный нейрон
4. Действие раздражителей вызывает возникновение нервного импульса в
А) чувствительных нейронах
Б) двигательных нейронах
В) рецепторах
Г) вставочных нейронах
5. По чувствительному нейрону возбуждение направляется
А) в центральную нервную систему
Б) к исполнительному органу
В) к рецепторам
Г) к мышцам
6. Рецепторы - это чувствительные образования, которые
А) передают импульсы в центральную нервную систему
Б) передают нервные импульсы со вставочных нейронов на исполнительные
В) воспринимают раздражения и преобразуют энергию раздражителей в процесс нервного возбуждения
Г) воспринимают нервные импульсы от чувствительных нейронов
7. Нервные импульсы передаются в мозг по нейронам
А) двигательным
Б) вставочным
В) чувствительным
Г) исполнительным
8. Рефлекторная дуга заканчивается
А) исполнительным органом
Б) чувствительным нейроном
В) рецептором
Г) вставочным нейроном
9. Нервные импульсы от рецепторов в центральную нервную систему проводят
А) чувствительные нейроны
Б) двигательные нейроны
В) чувствительные и двигательные нейроны
Г) вставочные и двигательные нейроны
10. Нервные образования в теле человека, воспринимающие раздражения из внешней среды, - это
А) нервы
Б) рецепторы
В) головной мозг
Г) нервные узлы
11. Преобразование раздражений в нервный импульс происходит в
А) двигательных нервах
Б) головном мозге
В) рецепторах
Г) спинном мозге
12. Нервные импульсы передаются от органов чувств человека по нейронам
А) чувствительным
Б) двигательным
В) вставочным
Г) исполнительным
13. Воспринимают информацию и преобразуют её в нервные импульсы
А) рецепторы
Б) нервные узлы
В) исполнительные органы
Г) вставочные нейроны
14. Условный рефлекс
А) наследуется потомством, но не сохраняется в течение жизни
Б) легко приобретается и теряется организмом в течение жизни
В) наследуется потомством от родителей и сохраняется в течение
Г) является постоянной реакцией организма на строго определенные раздражители внешней среды
15. Пример условного рефлекса у кошки
А) выделение слюны во время еды
Б) отдергивание лапы при ожоге
В) реакция на кличку
Г) реакция на резкий звук
16. Слюноотделение у человека при виде лимона - рефлекс
А) условный
Б) безусловный
В) защитный
Г) ориентировочный
17) Условные рефлексы у человека образуются в процессе
А) индивидуального развития
Б) формирования вида
В) исторического развития
Г) образования зародыша
18. Примером условного рефлекса является
А) выделение слюны при виде разрезанного лимона
Б) выделение слюны во время кормления
В) сужение зрачка при воздействии сильного света
Г) отдергивание руки при внезапном уколе
19. Реакция ребёнка на бутылочку с молоком - это рефлекс, который
А) передаётся по наследству
Б) формируется без участия коры больших полушарий
В) приобретаются в течение жизни
Г) сохраняется в течение всей жизни
20. Защитный рефлекс чихания
А) не передается по наследству
Б) является условным
В) ослабевает в течение жизни
Г) характерен для всех особей вида
21. Реакции, приобретенные человеком и животными в течение жизни и обеспечивающие приспособление к меняющимся условиям среды, называют
А) условными рефлексами
Б) инстинктами
В) безусловными рефлексами
Г) рефлексами, передающимися по наследству
22. Безусловный рефлекс
А) передается по наследству
Б) приобретается в течение жизни
В) вырабатывается на определенные сигналы
Г) лежит в основе различных внешних сигналов
23. Безусловные рефлексы человека и животных обеспечивают
В) освоение организмом новых двигательных умений
24. Реакция человека на зеленый цвет светофора - это рефлекс
А) врожденный
Б) приобретенный
В) безусловный
Г) наследуемый
25. Выделение желудочного сока в ответ на раздражение пищей рецепторов ротовой полости
А) возбуждение
Б) торможение
В) безусловный рефлекс
Г) саморегуляция
26. Биологическое значение условных рефлексов для организма состоит в том, что они
А) поддерживают постоянство внутренней среды
Б) видоспецифичны и сохраняются в течение всей жизни
В) способствуют взаимодействию функциональных систем
Г) обеспечивают приспособление к изменяющимся условиям среды
27. В процессе жизни животные приобретают рефлексы
А) безусловные
Б) передающиеся по наследству
В) характерные для всех особей данного вида
Г) позволяющие им выживать в изменяющихся условиях
28. Условным рефлексом у человека является
А) сужения зрачка при ярком свете
Б) поворот головы на резкий звук
В) выделение слюны при попадании пищи в ротовую полость
Г) реакция на смысл слова
29. Примером условного рефлекса может служить
1) умение кататься на велосипеде
2) выделение слюны при еде
3) желание спать при усталости
4) чихание и кашель
30. Условные рефлексы человека и животных обеспечивают
А) приспособление организма к постоянным условиям среды
Б) приспособление организма к меняющемуся внешнему миру
В) освоение организмов новых двигательных, умений
Г) различение животными команд дрессировщика
31. Врождённые рефлексы
А) являются видовыми признаками организма
Б) характерны для отдельных особей вида
В) требуют дополнительных условий для осуществления
Г) не передаются по наследству
Основной формой нервной деятельности является рефлекс. Рефлекс - причинно-обусловленная реакция организма на изменения внешней или внутренней среды, осуществляемая при обязательном участии центральной нервной системы в ответ на раздражение рецепторов. За счет рефлексов происходит возникновение, изменение или прекращение какой-либо деятельности организма.
Нервный путь, по которому распространяется возбуждение при осуществлении рефлексов, называют рефлекторной дугой .
Рефлекторные дуги состоят из пяти компонентов: 1) рецептор; 2) афферентный нервный путь; 3) рефлекторный центр; 4) эфферентный нервный путь; 5) эффектор (рабочий орган).
Рецептор - это чувствительное нервное окончание, воспринимающее раздражение. В рецепторах энергия раздражителя превращается в энергию нервного импульса. Различают: 1) экстерорецепторы - возбуждаются под влиянием раздражений из окружающей среды (рецепторы кожи, глаза, внутреннего уха, слизистой оболочки носа и ротовой полости); 2) интерорецепторы - воспринимают раздражения из внутренней среды организма (рецепторы внутренних органов, сосудов); 3) проприорецепторы - реагируют на изменение положения отдельных частей тела в пространстве (рецепторы мышц, сухожилий, связок, суставных сумок).
Афферентный нервный путь представлен отростками рецепторных нейронов, несущих возбуждения в центральную нервную систему.
Рефлекторный центр состоит из группы нейронов, расположенных на различных уровнях центральной нервной системы и передающих нервные импульсы с афферентного на эфферентный нервный путь.
Эфферентный нервный путь проводит нервные импульсы от центральной нервной системы к эффектору.
Эффектор - исполнительный орган, деятельность которого изменяется под влиянием нервных импульсов, поступающих к нему по образованиям рефлекторной дуги. Эффекторами могут быть мышцы или железы.
Рефлекторные дуги могут быть простыми и сложными. Простая рефлекторная дуга состоит из двух нейронов - воспринимающего и эффекторного, между которыми имеется один синапс. Схема такой двухнейронной рефлекторной дуги приведена на рис. 71.
Примером простой рефлекторной дуги являются рефлекторные дуги сухожильных рефлексов, например рефлекторная дуга коленного рефлекса.
Рефлекторные дуги большинства рефлексов включают не два, а большее количество нейронов: рецепторный, один или несколько вставочных и эффекторный. Такие рефлекторные дуги называют сложными, многонейронными. Схема сложной (трехнейронной) рефлекторной дуги приведена на рис. 72.
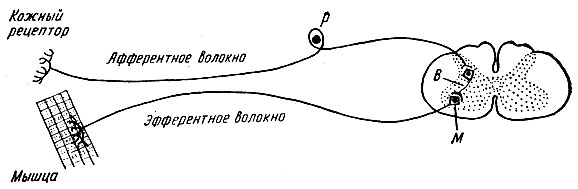
В настоящее время установлено, что во время ответной реакции эффектора возбуждаются многочисленные нервные окончания, имеющиеся в рабочем органе. Нервные импульсы теперь уже от эффектора вновь поступают в центральную нервную систему и информируют ее о правильности ответа рабочего органа. Таким образом, рефлекторные дуги являются не разомкнутыми, а кольцевыми образованиями.
Рефлексы отличаются большим многообразием. Их можно классифицировать по ряду признаков: 1) по биологическому значению (пищевые, оборонительные, половые); 2) в зависимости от вида раздражаемых рецепторов: экстероцептивные, интероцептивные и проприоцептивные; 3) по характеру ответной реакции: двигательные или моторные (исполнительный орган - мышца), секреторные (эффектор - железа), сосудодвигательные (сужение или расширение кровеносных сосудов).
Все рефлексы целостного организма могут быть разделены на две большие группы: безусловные и условные. Различия между ними будут разобраны в главе XII.
Деятельности нервной системы присущ рефлекторный характер. Еще в XVIIв. французский философ и математик Рене Декарт дал описание рефлекторному акту. Он отметил реакцию организма на раздражение и высказал предположение о существовании пути, по которому проходит нервное возбуждение. Сам термин «рефлекс» был выдвинут позже – в XVIIIв.– чешским ученым Дж. Прохазкой (с латинского «рефлекс» – отраженное действие). В дальнейшем И. М. Сеченов в своей работе «Рефлексы головного мозга» доказал, что ответы нервной системы на различного рода раздражения протекают по рефлекторному механизму, т.е. все сознательные и бессознательные действия имеют рефлекторное происхождение. Под рефлексом понимается конкретная реакция организма на раздражитель внутренней среды либо поступивший извне с обязательным участием центральной нервной системы. Рефлексы принято называть функциональными единицами нервной деятельности.
Механизмы рефлексов рефлекторной дуги
Рефлексы поддаются классификации по различным критериям. Так, в зависимости от уровня замыкания дуги, т.е. по месту локализации рефлекторного центра, рефлексы подразделяют на спинальные (рефлекс замыкается в спинном мозге), бульбарные (рефлекторный центр – продолговатый мозг), мезэнцефальные (замыкание строения рефлекторной дуги осуществляется в среднем мозге), диэнцефальные и кортикальные рефлекторные центры находятся в конечном мозге и коре больших полушарий соответственно. По эффекторному признаку они бывают соматические, когда эфферентный путь рефлекса осуществляет двигательную иннервацию скелетной мускулатуры, и вегетативные, когда эффекторами являются внутренние органы. В зависимости от вида раздражаемых рецепторов рефлексы делят на экстероцептивные (если рецептор воспринимает информацию из внешней среды), проприо-цептивные (рефлекторная дуга начинается от рецепторов костно-мышечно-сухожильного аппарата) и интероцептивные (от рецепторов внутренних органов). Интероцептивные рефлексы, в свою очередь, подразделяются на висцеро-висцеральные (рефлекторная дуга связывает два внутренних органа), висцеро-мышечные (рецепторы находятся на мышечно-сухожильном аппарате, эффектор – внутренний орган) и висцеро-кутанные (рецепторы локализованы в коже, рабочие органы – внутренности). По Павлову, рефлексы делят на условные (выработанные в течение жизни, специфичные для каждого индивида) и безусловные (врожденные, видоспецифичные: пищевые, половые, оборонительно-двигательные, гомеостатические и др.).
Как устроена рефлекторная дуга
Независимо от вида рефлекса его рефлекторная дуга содержит рецептор, афферентный путь, нервный центр, эфферентный путь, рабочий орган и обратную связь. Исключением являются аксон-рефлексы, строение рефлекторной дуги которого располагается в пределах одного нейрона: чувствительные отростки генерируют центростремительные импульсы, которые, проходя через тело нейрона, по аксону распространяются в центральную нервную систему, а по ответвлению аксона импульсы доходят уже до эффектора. Подобные рефлексы относят к функционированию метасимпатической нервной системы, через них, например, осуществляются механизмы регулирования тонуса сосудов и деятельности желез кожи.
Функцию восприятия раздражения и превращения его в энергию возбуждения выполняют рецепторы рефлекторных дуг. Ре-цепторная энергия возбуждения носит характер локального ответа, что имеет значение в градации возбуждения по силе.
Исходя из строения и происхождения рецепторов, их можно разделить на первично-чувствующие, вторично-чувствующие и свободные нервные окончания. У первых в качестве рецептора действует сам нейрон (развивается из нейроэпителия), т.е. между раздражителем и первым афферентным нейроном нет структур-посредников. Локальный ответ первично-чувствующих рецепторов – рецепторный потенциал – является и генераторным потенциалом, т.е. вызывающим возникновение потенциала действия на мембране афферентного волокна. К первично-чувствующим рецепторам относят зрительные, обонятельные, хемо– и ба-рорецепторы сердечно-сосудистой системы.
Вторично-чувствующие клетки представляют собой специальные структуры ненервного происхождения, которые с помощью синаптических нейрорецепторных контактов взаимодействуют с дендритами псевдоуниполярных чувствительных клеток. Рецеп-торный потенциал, возникающий под действием раздражителя, во вторично-чувствующих клетках не является генераторным и не вызывает возникновения потенциала действия на мембране афферентного волокна. Возбуждающий постсинаптический потенциал возникает лишь через механизм выделения рецептор-ной клеткой медиатора. Градация силы раздражителя осуществляется посредством экскреции различных количеств медиатора (чем больше выделяется медиатора, тем сильнее раздражитель).
Ко вторично-чувствующим клеткам относят слуховые, вестибулярные, каротидные, тактильные и другие рецепторы. Иногда в связи с особенностями функционирования к этой группе относят фоторецепторы, которые с анатомической точки зрения и в связи с происхождением из нейроэпителия являются вторично-чувствующими.
Свободные нервные окончания представляют собой ветвления дендритов псевдоуниполярных чувствительных клеток и локализуются почти во всех тканях человеческого тела.
По энергетической природе раздражителя, на который реагирует рецептор, они делятся на механорецепторы (тактильные, барорецепторы, волюморецепторы, слуховые, вестибулярные; они, как правило, воспринимают механическое раздражение при помощи выростов клетки), хеморецепторы (обонятельные), хемо-рецепторы сосудов, центральной нервной системы, фоторецепторы (воспринимают раздражение через палочко– и колбочковид-ные выросты клетки), терморецепторы (реагируют на изменение «тепло-холод» – тельца Руфини и колбы Краузе слизистых оболочек) и ноцицепторы (неинкапсулированные болевые окончания).
Пострецепторные образования рефлекторной дуги
Пострецепторным образованием строения рефлекторных дуг является афферентный путь, образованный псевдоуниполярным чувствительным нейроном, тело которого лежит в спинальном ганглии, а аксоны образуют задние корешки спинного мозга. Функция афферентного пути – проведение информации к центральному звену, более того, на данном этапе происходит кодирование информации. Для этих целей в организме позвоночных применяется двоичный код, составленный из пачек (залпов) импульсов и промежутков между ними. Существует два основных вида кодирования: частотное и пространственное.
Первое заключается в формировании различного числа импульсов в пачке, разного количества пачек, их длительности и длительности перерывов между ними в зависимости от силы нанесенного на рецептор раздражения. Пространственное кодирование осуществляет градацию силы раздражителя, задействуя различное количество нервных волокон, по которым одновременно проводится возбуждение.
В состав афферентного пути входят преимущественно А-α, А-β и А-δ волокна.
Пройдя по волокнам, нервный импульс попадает в рефлекторный центр, который в анатомическом смысле представляет собой совокупность нейронов, расположенных на определенном уровне центральной нервной системы и принимающих участие в формировании данного рефлекса. Функция рефлекторного центра состоит в анализе и синтезе информации, а также в переключении информации с афферентного на эфферентный путь.
Функции рефлекторной дуги
В зависимости от отдела нервной системы (соматического и автономного) рефлексы, центр которых расположен в спинном мозге, различаются по локализации вставочных нейронов. Так, для соматической нервной системы рефлекторный центр расположен в промежуточной зоне между передними и задними рогами спинного мозга. Рефлекторный центр вегетативной нервной системы (тела вставочных нейронов) лежит в задних рогах. Соматический и вегетативный отделы нервной системы также отличаются по локализации эфферентных нейронов. Тела моторных нейронов соматической нервной системы лежат в передних рогах спинного мозга, тела преганглионарных нейронов автономной системы – на уровне средних рогов.
Аксоны обоих типов клеток формируют эфферентный путь рефлекторной дуги. В соматической нервной системе он непрерывающийся, его составляют волокна типа А-α. Исключением являются лишь А-γ волокна, проводящие возбуждение от клеток спинного мозга к интрафузальным волокнам мышечных веретен. Эфферентный путь автономной нервной системы прерывается в вегетативном ганглии, расположенном или интрамурально (парасимпатическая часть), или близ спинного мозга (отдельно или в симпатическом стволе – симпатическая часть). Преганглио-нарное волокно относится к В-волокнам, постганглионарное – к группе С.
Рабочим органом для соматического отдела нервной системы является поперечно-полосатая скелетная мышца, в вегетативной дуге эффектор – железа либо мышца (гладкая или поперечно-полосатая сердечная). Между эфферентным путем и рабочим органом расположен химический мионевральный либо нейросекре-торный синапс.
Рефлекторная дуга замыкается в кольцо благодаря обратной афферентации – потоку импульсов от рецепторов эффектора обратно в рефлекторный центр. Функция обратной связи – сигнализация в центральную нервную систему о выполненном действии. Если оно выполнено недостаточно, нервный центр возбуждается – рефлекс продолжается. Также за счет обратной афферентации осуществляется контроль периферической деятельности центральной нервной системой.
Различают отрицательную и положительную обратные связи. Первая при выполнении определенной функции запускает механизм, угнетающий эту функцию. Положительная обратная связь заключается в дальнейшей стимуляции функции, которая уже выполняется или в угнетении функции, которая уже угнетена. Положительная обратная афферентация встречается редко, так как приводит биологическую систему в неустойчивое положение.
Простые (моносинаптические) рефлекторные дуги состоят лишь из двух нейронов (афферентного и эфферентного) и различаются только в проприоцептивных рефлексах. Остальные дуги включают все выше указанные компоненты.
Рефлекторная дуга - это цепь нейронов от периферического рецептора через центральную нервную систему к периферическому эффектору. Элементами рефлекторной дуги являются периферический рецептор, афферентный путь, один или больше вставочных нейронов, эфферентный путь и эффектор.
Все рецепторы участвуют в тех или иных рефлексах, так что их афферентные волокна служат афферентным путем соответствующей рефлекторной дуги. Число вставочных нейронов всегда больше одного, кроме моносинаптического рефлекса растяжения. Эфферентный путь представлен либо двигательными аксонами, либо постганглионарными волокнами вегетативной нервной системы, а эффекторами являются скелетные мышцы и гладкие мышцы, сердце, железы.
Время от начала стимула до реакции эффектора называется временем рефлекса. В большинстве случаев оно определяется в основном временем проведения в афферентных и в эфферентных путях и в центральной части рефлекторной дуги, к которому следует прибавить время трансформации стимула в рецепторе в распространяющийся импульс, время передачи через синапсы в центральной нервной системе (синаптическая задержка), время передачи от эфферентного пути к эффектору и время активации эффектора.
Рефлекторные дуги делятся на несколько типов
1. Моносинаптические рефлекторные дуги - в такой дуге участвует только один синапс, находящийся в центральной нервной системе. Такие рефлексы весьма обычны у всех позвоночных, они участвуют в регуляции мышечного тонуса и позы (например, коленный рефлекс). В этих дугах нейроны не доходят до головного мозга, и рефлекторные акты осуществляются без его участия, так как они стереотипны и не требуют обдумывания или сознательного решения. Они экономны в отношении числа участвующих центральных нейронов и обходятся без вмешательства головного мозга.
2. Полисинаптические спинномозговые рефлекторные дуги - в них участвуют по меньшей мере два синапса, находящиеся в ЦНС, так как в дугу включен третий нейрон - вставочный, или промежуточный нейрон. Здесь имеются синапсы между сенсорным нейроном и вставочным нейроном и между вставочным и двигательным нейронами. Такие рефлекторные дуги позволяют организму осуществлять автоматические непроизвольные реакции, необходимые для приспособления к изменениям внешней среды (например, зрачковый рефлекс или сохранение равновесия при передвижении) и к изменениям в самом организме (регуляция частоты дыхания, кровяного давления и т.п.).
3. Полисинаптические рефлекторные дуги с участием как спинного, так и головного мозга - в рефлекторных дугах этого типа имеется синапс в спинном мозге между сенсорным нейроном и нейроном, посылающим импульсы в головной мозг.
Рефлексы поддаются классификации по различным критериям. Так, в зависимости от уровня замыкания дуги, т.е. по месту локализации рефлекторного центра, рефлексы подразделяют на спинальные (рефлекс замыкается в спинном мозге), бульбарные (рефлекторный центр - продолговатый мозг), мезэнцефальные (замыкание рефлекторной дуги осуществляется в среднем мозге), диэнцефальные и кортикальные рефлекторные центры находятся в конечном мозге и коре больших полушарий соответственно.
По эффекторному признаку они бывают соматические, когда эфферентный путь рефлекса осуществляет двигательную иннервацию скелетной мускулатуры, и вегетативные, когда эффекторами являются внутренние органы.
В зависимости от вида раздражаемых рецепторов рефлексы делят на экстероцептивные (если рецептор воспринимает информацию из внешней среды), проприоцептивные (рефлекторная дуга начинается от рецепторов костно-мышечно-сухожильного аппарата) и интероцептивные (от рецепторов внутренних органов).
Интероцептивные рефлексы, в свою очередь, подразделяются на висцеро-висцеральные (рефлекторная дуга связывает два внутренних органа), висцеро-мышечные (рецепторы находятся на мышечно-сухожильном аппарате, эффектор - внутренний орган) и висцеро-кутанные (рецепторы локализованы в коже, рабочие органы - внутренности).
По Павлову, рефлексы делят на условные (выработанные в течение жизни, специфичные для каждого индивида) и безусловные (врожденные, видоспецифичные: пищевые, половые, оборонительно-двигательные, гомеостатические и др.).
Независимо от вида рефлекса его рефлекторная дуга содержит рецептор, афферентный путь, нервный центр, эфферентный путь, рабочий орган и обратную связь. Исключением являются аксон-рефлексы, рефлекторная дуга которого располагается в пределах одного нейрона: чувствительные отростки генерируют центростремительные импульсы, которые, проходя через тело нейрона, по аксону распространяются в центральную нервную систему, а по ответвлению аксона импульсы доходят уже до эффектора. Подобные рефлексы относят к функционированию метасимпатической нервной системы, через них, например, осуществляются механизмы регулирования тонуса сосудов и деятельности желез кожи.
Функцию восприятия раздражения и превращения его в энергию возбуждения выполняют рецепторы рефлекторных дуг. Рецепторная энергия возбуждения носит характер локального ответа, что имеет значение в градации возбуждения по силе.
Исходя из строения и происхождения рецепторов, их можно разделить на первично-чувствующие, вторично-чувствующие и свободные нервные окончания. У первых в качестве рецептора действует сам нейрон (развивается из нейроэпителия), т.е. между раздражителем и первым афферентным нейроном нет структур-посредников. Локальный ответ первично-чувствующих рецепторов - рецепторный потенциал - является и генераторным потенциалом, т.е. вызывающим возникновение потенциала действия на мембране афферентного волокна. К первично-чувствующим рецепторам относят зрительные, обонятельные, хемо- и барорецепторы сердечно-сосудистой системы.
Вторично-чувствующие клетки представляют собой специальные структуры ненервного происхождения, которые с помощью синаптических нейрорецепторных контактов взаимодействуют с дендритами псевдоуниполярных чувствительных клеток. Рецепторный потенциал, возникающий под действием раздражителя, во вторично-чувствующих клетках не является генераторным и не вызывает возникновения потенциала действия на мембране афферентного волокна. Возбуждающий постсинаптический потенциал возникает лишь через механизм выделения рецепторной клеткой медиатора. Градация силы раздражителя осуществляется посредством экскреции различных количеств медиатора (чем больше выделяется медиатора, тем сильнее раздражитель).
Ко вторично-чувствующим клеткам относят слуховые, вестибулярные, каротидные, тактильные и другие рецепторы. Иногда в связи с особенностями функционирования к этой группе относят фоторецепторы, которые с анатомической точки зрения и в связи с происхождением из нейроэпителия являются вторично-чувствующими.
Свободные нервные окончания представляют собой ветвления дендритов псевдоуниполярных чувствительных клеток и локализуются почти во всех тканях человеческого тела.
По энергетической природе раздражителя, на который реагирует рецептор, они делятся на механорецепторы (тактильные, барорецепторы, волюморецепторы, слуховые, вестибулярные; они, как правило, воспринимают механическое раздражение при помощи выростов клетки), хеморецепторы (обонятельные), хеморецепторы сосудов, центральной нервной системы, фоторецепторы (воспринимают раздражение через палочко- и колбочковид-ные выросты клетки), терморецепторы (реагируют на изменение «тепло-холод» - тельца Руфини и колбы Краузе слизистых оболочек) и ноцицепторы (неинкапсулированные болевые окончания).
Пострецепторным образованием рефлекторных дуг является афферентный путь, образованный псевдоуниполярным чувствительным нейроном, тело которого лежит в спинальном ганглии, а аксоны образуют задние корешки спинного мозга. Функция афферентного пути - проведение информации к центральному звену, более того, на данном этапе происходит кодирование информации. Для этих целей в организме позвоночных применяется двоичный код, составленный из пачек (залпов) импульсов и промежутков между ними. Существует два основных вида кодирования: частотное и пространственное.
Первое заключается в формировании различного числа импульсов в пачке, разного количества пачек, их длительности и длительности перерывов между ними в зависимости от силы нанесенного на рецептор раздражения. Пространственное кодирование осуществляет градацию силы раздражителя, задействуя различное количество нервных волокон, по которым одновременно проводится возбуждение.
В состав афферентного пути входят преимущественно А-б, А-в и А-д волокна.
Пройдя по волокнам, нервный импульс попадает в рефлекторный центр, который в анатомическом смысле представляет собой совокупность нейронов, расположенных на определенном уровне центральной нервной системы и принимающих участие в формировании данного рефлекса. Функция рефлекторного центра состоит в анализе и синтезе информации, а также в переключении информации с афферентного на эфферентный путь.
В зависимости от отдела нервной системы (соматического и автономного) рефлексы, центр которых расположен в спинном мозге, различаются по локализации вставочных нейронов. Так, для соматической нервной системы рефлекторный центр расположен в промежуточной зоне между передними и задними рогами спинного мозга. Рефлекторный центр вегетативной нервной системы (тела вставочных нейронов) лежит в задних рогах. Соматический и вегетативный отделы нервной системы также отличаются по локализации эфферентных нейронов. Тела моторных нейронов соматической нервной системы лежат в передних рогах спинного мозга, тела преганглионарных нейронов автономной системы - на уровне средних рогов.
Аксоны обоих типов клеток формируют эфферентный путь рефлекторной дуги. В соматической нервной системе он непрерывающийся, его составляют волокна типа А-б. Исключением являются лишь А-г волокна, проводящие возбуждение от клеток спинного мозга к интрафузальным волокнам мышечных веретен. Эфферентный путь автономной нервной системы прерывается в вегетативном ганглии, расположенном или интрамурально (парасимпатическая часть), или близ спинного мозга (отдельно или в симпатическом стволе - симпатическая часть). Преганглио нарное волокно относится к В-волокнам, постганглионарное - к группе С.
Рабочим органом для соматического отдела нервной системы является поперечно-полосатая скелетная мышца, в вегетативной дуге эффектор - железа либо мышца (гладкая или поперечно-полосатая сердечная). Между эфферентным путем и рабочим органом расположен химический мионевральный либо нейросекреторный синапс.
Рефлекторная дуга замыкается в кольцо благодаря обратной афферентации - потоку импульсов от рецепторов эффектора обратно в рефлекторный центр. Функция обратной связи - сигнализация в центральную нервную систему о выполненном действии. Если оно выполнено недостаточно, нервный центр возбуждается - рефлекс продолжается. Также за счет обратной афферентации осуществляется контроль периферической деятельности центральной нервной системой.
Различают отрицательную и положительную обратные связи. Первая при выполнении определенной функции запускает механизм, угнетающий эту функцию. Положительная обратная связь заключается в дальнейшей стимуляции функции, которая уже выполняется или в угнетении функции, которая уже угнетена. Положительная обратная афферентация встречается редко, так как приводит биологическую систему в неустойчивое положение.
Простые (моносинаптические) рефлекторные дуги состоят лишь из двух нейронов (афферентного и эфферентного) и различаются только в проприоцептивных рефлексах. Остальные дуги включают все выше указанные компоненты.
Физиологические свойства и функциональная значимость нервных волокон
Нервные волокна имеют самую высокую возбудимость, самую высокую скорость проведения возбуждения, самый короткий рефрактерный период, высокую лабильность. Это обеспечивается высоким уровнем обменных процессов и низкой величиной мембранного потенциала.
Функция: проведение нервных импульсов от рецепторов к центральной нервной системе и обратно.
Особенности строения и виды нервных волокон
Нервное волокно - аксон - покрыт клеточной мембраной.
Выделяют 2 вида нервных волокон:
Безмиелиновые нервные волокна - один слой швановских клеток, между ними - щелевидные пространства. Клеточная мембрана на всем протяжении контактирует с окружающей средой. При нанесении раздражения возбуждение возникает в месте действия раздражителя. Безмиелиновые нервные волокна обладают электрогенными свойствами (способностью генерировать нервные импульсы) на всем протяжении.
Миелиновые нервные волокна - покрыты слоями шванновских клеток, которые местами образуют перехваты Ранвье (участки без миелина) через каждые 1 мм. Продолжительность перехвата Ранвье 1 мкм. Миелиновая оболочка выполняет трофическую и изолирующую функции (высокое сопротивление). Участки, покрытые миелином не обладают электрогенными свойствами. Ими обладают перехваты Ранвье. Возбуждение возникает в ближайшем к месту действия раздражителя перехвата Ранвье. В перехватах Ранвье высокая плотность Nа-каналов, поэтому в каждом перехвате Ранвье происходит усиление нервных импульсов.
Перехваты Ранвье выполняют функцию ретрансляторов (генерируют и усиливают нервные импульсы).
Механизм проведения возбуждения по нервному волокну
1885 г. - Л. Герман - между возбужденными и невозбужденными участками нервного волокна возникают круговые токи.
При действии раздражителя имеется разность потенциалов между наружной и внутренней поверхностями ткани (участки несущие различные заряды). Между этими участками возникает электрический ток (движение ионов Nа+). Внутри нервного волокна возникает ток от положительного полюса к отрицательному полюсу, т. е. ток направлен от возбужденного участка к невозбужденному. Этот ток выходит через невозбужденный участок и вызывает его перезарядку. На наружной поверхности нервного волокна ток идет от невозбужденного участка к возбужденному. Этот ток не изменяет состояние возбужденного участка, т. к. он находится в состоянии рефрактерности.
Доказательство наличия круговых токов: нервное волокно помещают в раствор NaCl и регистрируют скорость проведения возбуждения. Затем нервное волокно помещают в масло (повышается сопротивление) - скорость проведения уменьшается на 30 %. После этого нервное волокно оставляют на воздухе - скорость проведения возбуждения уменьшается на 50 %.
Особенности проведения возбуждения по миелиновым и безмиелиновым нервным волокнам:
миелиновые волокна - имеют оболочку обладающую высоким сопротивлением, электрогенные свойства только в перехватах Ранвье. Под действием раздражителя возбуждение возникает в ближайшем перехвате Ранвье. Соседний перехват в состоянии поляризации. Возникающий ток вызывает деполяризацию соседнего перехвата. В перехватах Ранвье высокая плотность Nа-каналов, поэтому в каждом следующем перехвате возникает чуть больший (по амплитуде) потенциал действия, за счет этого возбуждение распространяется без декремента и может перескакивать через несколько перехватов. Это сальтаторная теория Тасаки. Доказательство теории - в нервное волокно вводили препараты, блокирующие несколько перехватов, но проведение возбуждения регистрировалось и после этого. Это высоко надежный и выгодный способ, т. к. устраняются небольшие повреждения, увеличивается скорость проведения возбуждения, уменьшаются энергетические затраты;
безмиелиновые волокна - поверхность обладает электрогенными свойствами на всем протяжении. Поэтому малые круговые токи возникают на расстоянии в несколько микрометров. Возбуждение имеет вид постоянно бегущей волны.
Этот способ менее выгоден: большие затраты энергии (на работу Nа-К-насоса), меньшая скорость проведения возбуждения.
Классификация нервных волокон
Нервные волокна классифицируются по:
· длительности потенциала действия;
· строению (диаметру) волокна;
· скорости проведения возбуждения.
Выделяют следующие группы нервных волокон:
· группа А (альфа, бета, гамма, дельта) - самый короткий потенциал действия, самая толстая миелиновая оболочка, самая высокая скорость проведения возбуждения;
· группа В - миелиновая оболочка менее выражена;
· группа С - без миелиновой оболочки.
Морфологические отличия дендритов от аксонов
1. У отдельного нейрона имеется несколько дендритов, аксон всегда один.
2. Дендриты всегда короче аксона. Если размеры дендритов непревышают 1,5-2 мм, то аксоны могут достигать 1м и более.
3. Дендриты плавно отходят от тела клетки и постепеннопостоянный диаметр на значительном протяжении.
4. Дендриты ветвятся обычно под острым углом, и ветвинаправлены от клетки. Аксоны отдают коллатерали чаще всего под прямым углом, ориентация коллатералей не связана непосредственно с положением клеточного тела.
5. Рисунок дендритического ветвления у клеток одного типа более постоянен, чем разветвления аксона этих клеток.
6. Дендриты зрелых нейронов бывают покрыты дендритическими шипиками, которые отсутствуют на соме и начальной части дендритных стволов. Аксоны не имеют шипиков.
7. Дендриты никогда не имеют мякотной оболочки. Аксоны часто окружены миелином.
8. Дендриты имеют более регулярную пространственнуюорганизацию микротрубочек, в аксонах в основном преобладают нейрофиламенты и микротрубочки расположены менее упорядочение
9. В дендритах, в особенности в их проксимальных участках,имеются эндоплазматический ретикулум и рибосомы, чего нет в аксонах.
10. Поверхность дендритов в большинстве случаев контактирует с синоптическими бляшками и имеет активные зоны с постсинаптической специализацией.
Строение дендритов
Если о геометрии дендритов, длине их ветвей, ориентации имеется сравнительно большая литература, то о внутреннем строении, о строении отдельных компонентов их цитоплазмы есть лишь отдельные разрозненные сведения. Эти сведения стали возможными только с внедрением в нейрогистологию электронно - микроскопических исследований.
Основные характерные черты дендрита, которые выделяют его на электронно-микроскопических срезах:
1)отсутствие миелиновой оболочки,
наличие правильной системы микротрубочек,
3) наличие на них активных зон синапсов с ясно выраженной электронной плотностью цитоплазмы дендрита,
4) отхождение от общего ствола дендрита шипиков,
5) специально организованные зоны узлов ветвлений,
6) вкрапление рибосом,
7) наличие в проксимальных участках гранулированного и не гранулированного эндоплазматического ретикулума.
Наиболее примечательной особенностью цитоплазмы дендритов является наличие многочисленных микротрубочек. Они хорошо выявляются как на поперечных срезах, так и на продольных. Начиная от проксимального участка дендрита, микротрубочки идут параллельно длинной оси дендрита до его дистальных разветвлений. Микротрубочки следуют в дендрите параллельно друг другу, не соединяясь и не пересекаясь между собой. На поперечных срезах можно видеть, что расстояния между отдельными трубочками постоянны. Отдельные дендритические трубочки тянутся на довольно большие расстояния, часто следуя изгибам, которые могут быть по ходу дендритов. Число трубочек относительно постоянно на единицу площади поперечного сечения дендрита и составляет примерно 100 на 1 мкм. Это число характерно для любых дендритов, взятых из разных отделов центральной и периферической нервной системы, у разных видов животных.
Функция микротрубочек - транспорт веществ по отросткам нервных клеток.
При разрушении микротрубочек может нарушаться транспорт веществ в дендрите, и, таким образом, конечные отделы отростков лишаться притока питательных и энергетических веществ от тела клетки. Дендриты, для того чтобы сохранить в экстремальных условиях структуру синаптических контактов и тем самым обеспечить функцию межнейронального взаимодействия, восполняют дефицит питательных веществ за счет прилежащих к ним структур (синаптические бляшки, многослойную миелиновую оболочку мягкого волокна, а также фрагменты глиальных клеток).
Если действие патогенного фактора будет своевременно устранено, дендриты восстанавливают структуру и правильную пространственную организацию микротрубочек, тем самым восстанавливается и система транспорта веществ, которая присуща нормальному мозгу. Если же сила и продолжительность патогенного фактора будут значительными, то явления эндоцитоза вместо своей приспособительной функции могут стать для дендритов губительными, т. к. фагоцитированные фрагменты не смогут утилизироваться и, накапливаясь в цитоплазме дендритов, приведут к необратимым его повреждениям.
Нарушение в организации микротрубочек ведет к резкому изменению поведения животных. У животных, в эксперименте у которых были разрушены микротрубочки в дендритах наблюдалась дезорганизация сложных форм поведения при сохранности простых условных рефлексов. У человека это может привести к серьезным нарушениям в высшей нервной деятельности.
О том, что дендриты являются наиболее чувствительным локусом к действию патологического агента при психических заболевания, свидетельствуют некоторые работы американских ученых. Оказалось, что при старческом слабоумии (синильной деменции) и болезни Альцгеймера на препаратах мозга, обработанных по методу Гольджи, не выявляются отростки нервных клеток. Стволы дендритов кажутся как бы обгоревшими и обугленными. Не выявление этих отростков на гистологических препаратах мозга, вероятно, связано также с нарушением в этих отростках системы микротрубочек и нейрофиломентов.
В дендритах встречаются. Они следуют параллельно длинной оси дендрита, могут лежать отдельно или собираться в пучки, однако в цитоплазме нет их строгого расположения. Вероятно, вместе с микротрубочками могут быть эквивалентом нейрофибрилл.
Для всех дендритов ЦНС характерно увеличение поверхности за счет многократного дихотомического деления. При этом образуются в зонах деления особые расширительные площадки или узлы ветвления.
Нормальный анализ показывает, что в узле ветвления, к которому подходят две дендритные ветви, несущие каждая в отдельности собственный сигнал, могут осуществляться следующие операции. Через узел ветвления в общий ствол и дальше к телу нейрона проходят:
или сигнал от одной ветви,
или только от другой,
или результат взаимодействия двух сигналов,
или же сигналы взаимно гасят друг друга.
В цитоплазме узла ветвления имеются почти все компоненты, которые характерны для тела нервной клетки, и участки резко отличаются по своему строению от цитоплазмы общего дендритного ствола и ветвей, полученных при делении. В узлах ветвления содержится повышенное число митохондрий, гранулированный и гладкий ретикулум, видны скопления одиночных рибосом и рибосом, собранных в розетки. Эти компоненты (гранулированный и гладкий ретикулум, рибосомы) принимают непосредственное участие в синтезе белка. Скопление митохондрий в этих местах указывает на интенсивность окислительных процессов.
Функции дендритов
Хотелось бы отметить, что основные трудности, с которыми сталкивается исследователь при изучении функции дендритов, - это отсутствие сведений о свойствах мембраны дендрита (в отличие от мембраны тела нейрона) из-за невозможности введения микроэлектрода внутрь дендрита.
Оценивая в целом геометрию дендритов, распределение синапсов и особое строение цитоплазмы в местах дендритных ветвлений, можно говорить о специальных локусах нейрона со своей собственной функцией. Самое простое, что можно было бы приписать дендритным площадкам в местах ветвления - это трофическая функция.
Из всего вышесказанного следует, что цитоплазма дендритов содержит много ультраструктурных компонентов, способных обеспечивать их важные функции. В дендрите есть определенные локусы, где его работа имеет свои особенности.
Главное назначение многочисленных дендритных разветвлений нервной клетки - это обеспечить взаимосвязь с другими нейронами. В коре головного мозга млекопитающих большая доля аксодендрических связей приходится на контакты с особыми специализированными выростами дендритов - дендрическими шипиками. Дендритические шипики являются филогенетически самыми молодыми образованиями в нервной системе. В онтогенезе они созревают значительно позже других нервных структур и представляют собой наиболее пластичный аппарат нервной клетки.
Как правило, дендрический шипик имеет в коре мозга млекопитающих характерную форму. (рис. 2). От основного дендритного ствола отходит сравнительно узкая ножка, которая заканчивается расширением - головкой. Вероятно такая форма дендритического придатка (наличие головки) связана, с одной стороны с увеличением площади синаптического контакта с аксонным окончанием, с другой служит для размещения внутри шипика специализированных органел, в частности шипикового аппарата, который имеется только в дендритических шипиках коры мозга млекопитающих. В этой связи кажется уместной аналогия с формой синаптического аксонного окончания, когда тонкое претерминальное волокно образует расширение. Это расширение (синаптическая бляшка) образует обширный контакт с иннервируемым субстратом и содержит внутри большой набор ультраструктурных компонентов (синаптические пузырьки, митохондрии, нейрофиламенты, гранулы гликогена).
Существует гипотеза (которую, в частности, разделяет и развивает нобелевский лаурят Ф. Крик) о том, что геометрия шипиков может меняться в зависимости от функционального состояния мозга. При этом узкая шейка шипика может расширяться, а сам шипик уплощается, в результате чего увеличивается эффективность аксо-шипикового контакта.
Если форма и размеры дендрических шипиков в коре мозга млекопитающих могут несколько варьировать, то наиболее постоянно в них наличие специфического шипикового аппарата. Он представляет собой комплекс взаимосвязанных канальцев (цистерн), расположенных, как правило, в головке шипика. Вероятно, это органелла связана с очень важными функциями, присущими филогенетически самым молодым мозговым образованиям, так как шипиковый аппарат встречается в основном в коре головного мозга, и только у высших животных.
Несмотря ни на что шипик является производным дендрита, в нем отсутствуют нейрофиламенты и дендритические трубочки, его цитоплазма содержит грубо или тонко гранулированный матрикс. Еще одной характерной чертой шипика в коре мозга является обязательное присутствие на них синаптических контактов с аксонными окончаниями. Цитоплазма шипика имеет специальные компоненты, которые отличают его от дендритных стволов. Можно отметить своеобразную триаду в цитоплазме шипика: субсинаптическая специализация активных зон - шипиковый аппарат - митохондрии. Учитывая многообразие сложных и важных функций, которые выполняют митохондрии, можно ожидать также сложных функциональных проявлений в «триадах» при синаптической передаче. Можно говорить о том, что цитоплазма дендритического шипика и шипиковый аппарат могут иметь непосредственное отношение к синаптической функции.
Дендритические шипики и концы дендритов также очень чувствительны к действию экстремальных факторов. При любом виде отравления (например, алкогольном, гипоксическом, тяжелыми металлами - свинцом, ртутью и т.д.) меняется количество выявленных шипиков на дендритах клеток коры больших полушарий. По всей вероятности, шипики при этом не исчезают, но у них нарушаются цитоплазматические компоненты, и они хуже импрегнируются солями тяжелых металлов. Так как шипики - один из структурных компонентов обеспечения межнейрональных контактов, то неполадки в них приводят к серьезным нарушениям функции мозга.
В некоторых случаях при кратковременном действии экстремального фактора может наступить на первый взгляд пара дорсальная ситуация, когда количество выявленных шипиков на дендритах клеток мозга не уменьшается, а увеличивается. Так, это наблюдается при экспериментальной ишемии мозга в начальный ее период. Параллельно с увеличением числа выявленных шипиков может улучшаться функциональное состояние мозга. В данном случае гипоксия является фактором, который способствует усилению метаболизма в нервной ткани, лучшей реализации резервов, не используемых в обычной обстановке, и быстрому сгоранию шлаков, накопленных в организме. Ультраструктурно это проявляется в более интенсивной проработке цитоплазмы шипиков, разрастании и увеличении цистерн шипикового аппарата. Вероятно, этот феномен положительного действия гипоксии наблюдается тогда, когда человек, испытывая большие физические нагрузки в условиях гипоксии, покоряет горные вершины. Эти трудности компенсируются затем более интенсивной продуктивной работой, как мозга, так и других органов.
Формирование дендритов
Дендриты и их межнейрональные связи формируются в процессе онтогенетического развития мозга. Причем дендриты, в частности апикальных, у молодых особей какое-то время остаются свободными для образования новых контактов. Участки дендрита, расположенные ближе к телу клетки, возможно, связаны с более прочными и простыми - натуральными условными рефлексами, а концы оставлены для образования новых связей, ассоциаций.
В зрелом возрасте на дендритах уже нет свободных от межнейрональных контактов участков, зато при старении прежде страдают именно концы дендритов и по насыщенности контактами
у старых особей они напоминают дендриты детского возраста. Это происходит как из-за того, что ослабляются транспортные белоксинтезирующие процессы в клетке, так и из-за нарушений кровоснабжения мозга. Может быть, именно здесь кроется та морфологическая основа для такого широко известного в неврологии и в обыденной жизни факта, когда старики трудно осваивают что-то новое, часто забывают текущие события и очень хорошо помнят о прошлом. То же самое наблюдается и при отравлению.
Как уже отмечалось, увеличение и усложнение дендритного древа в филогенезе необходимы не только для восприятия большого числа приходящих импульсов, но и для предварительной обработки.
Дендриты нейронов центральной нервной системы обладают синаптической функцией на всем протяжении, причем концевые участки нисколько не уступают в этом срединным. Если же речь идет о дистальных (концевых) участках апикальных дендритов пирамидных нейронов коры больших полушарий, то их доля в осуществлении межнейрональных взаимодействий еще более значительна, чем проксимальных. Там к большему числу концевых синаптических бляшек на самом стволе и разветвлениях апикального дендрита присоединяются еще контакты на дендритических шипиках.
Изучая эту проблему при помощи электронной микроскопии, исследователи также убедились в том, что концевые участки дендритов плотно покрыты синаптическими бляшками и, таким образом, принимают непосредственное участие в межнейрональных взаимодействиях. Электронная микроскопия также показала, что дендриты могут образовывать контакты между собой. Эти контакты могут быть или параллельными, которым большинство авторов приписывают электротонические свойства, или типичными ассиметричными синапсами с ясно выраженными органеллами, обеспечивающими химическую передачу. Такие дендро-дендритические контакты только еще начинают привлекать внимание исследователей. Итак, дендрит на всем своем протяжении выполняет синаптическую функцию. Каким же образом поверхность дендрита приспособлена для обеспечения контактов с аксонными окончаниями?
Поверхностная мембрана дендрита устроена так, чтобы максимальным образом использоваться для межнейрональных контактов. Дендрит весь изрыт углублениями, складками, карманами, имеет различного неровности рода микровыросты, шипы, грибовидные придатки и др. Все эти рельефы дендритных стволов соответствуют форме и размерам приходящих синаптических окончаний. Причем в различных отделах нервной системы и у разных животных рельеф дендритной поверхности имеет специфические особенности. Конечно же, самым замечательным выростом дендритической мембраны является дендритический шипик.
Дендриты очень чувствительны к действию различных экстремальных факторов. Нарушения в них приводят ко многим заболеваниям, например психическим расстройствам.
Рефлеторная дуга состоит из:
– рецепторов - воспринимающих раздражение.
– чувствительного (центростремительного, афферентного) нервного волокна, передающего возбуждение к центру
– нервного центра, где происходит переключение возбуждения с чувствительных нейронов на двигательные
– двигательного (центробежного, эфферентного) нервного волокна, несущего возбуждение от центральной нервной системы к рабочему органу
– эффектора- рабочего органа, который осуществляет эффект, реакцию в ответ на раздражение рецептора.
Рецепторы и рецептивные поля
Рецептор - клетки воспринимающие раздражение.
Рецептивное поле – это анатомическая область при раздражении которой вызывается данный рефлекс.
Рецептивные поля первично-чувствующих рецепторов организованы наиболее просто. Например, тактильное или ноцицептивное рецептивное поле кожной поверхности представляет собой разветвления одиночного чувствительного волокна.
Рецепторы, расположенные в различных участках рецептивного поля, имеют различную чувствительность к адекватному раздражению. В центре рецептивного поля обычно находится высокочувствительная зона, а ближе к периферии рецептивного поля чувствительность падает.
Рецептивные поля вторично-чувствующих рецепторов организованы аналогичным образом. Отличие состоит в том, что разветвления афферентного волокна оканчиваются не свободно, а имеют синаптические контакты с чувствительными клетками - рецепторами. Так организованы вкусовые, вестибулярные, акустические рецептивные поля.
Перекрытие рецептивных полей. Один и тот же участок чувствительной поверхности (например, кожи или сетчатки глаза) иннервируется несколькими чувствительными нервными волокнами, которые своими разветвлениями перекрывают рецептивные поля отдельных афферентных нервов.
Благодаря перекрытию рецептивных полей увеличивается общая сенсорная поверхность организма.
Классификация рефлексов.
По типу образования:
Условные (приобретенные)- отозваться на имя, слюна у собаки на свет.
Безусловные (врожденные)- мигательный глотательный, коленный.
По располож. рецепторов:
Экстероцептивные (кожные, зрительные, слуховые, обонятельные),
Интероцептивные (с рецепторов внутренних органов)
Проприоцептивные (с рецепторов мышц, сухожилий, суставов)
По эффекторам:
Соматические, или двигательные, (рефлексы скелетных мышц);
Вегетативные внутренних органов - пищеварительные, сердечно-сосудистые, выделительные, секреторные и др.
По биологическому происхождению:
Оборонительные, или защитные (ответ на тактильное болевое раздд.)
Пищеварительные(раздр. Рецепторов полости рта.)
Половые (гормоны в кровь)
Ориентировочные (поворот головы, тела)
Двигательные
Позотонические (поддерж. позы Тела)
По количеству синапсов:
Моносинаптические, дуги которых состоят из афферентного и эфферентного нейронов (например, коленный).
Полисинаптические, дуги которых содержат также 1 или несколько промежуточных нейронов и имеют 2 или несколько синаптических переключений. (сомат. и вегет. реф-сы).
Дисинаптические (2 синапса, 3 нейрона).
По характеру ответной реакции:
Моторные \ двигательные(мышечные сокращения)
Секреторные (выделение секреторной железы)
Сосудодвигательные (расширение и сужение сосудов)
Сердечные (изм. Работы мышци сердца.)
По длительности протекания:
фазные (быстрые) отдергивание руки
тонические (медленные) поддержание позы
По расположению нервного центра:
Спинальные (участвуют нейроны СМ) - одергивание Руки от горячего 2-4 сегменты, коленный рефлекс.
Рефлексы в головном мозге
Бульбарные (продолговатый мозг) - смыкание век при прикос. к роговице.
Мезенцифальные (средний м)- зрение ориентир.
Диэнцифальные (промежуточный мозг) – обоняние
Кортикальные (кора БП ГМ) – услов. реф.
Свойства нервных центров.
1. Односторонность распространения возбуждения .
Возбуждение передается с афферентного на эфферентный нейрон (причина: строение синапса).
Замедление передачи возбуждения.
Обусл. Наличием множества синапсов, также зависит от силы раздр.(суммация) и от физич-го сост. ЦНС(утомляемость).
3.Суммация сложение эффектов, ниже пороговых раздражителей.
Временная: реф. От пред. Имп-са еще не прошел, а след. Уже пришел.
Пространственная: смешение неск. Подпор. Им –сов обусл. Образов. Реф-са.
Центр облегчения и окклюзия.
Центр облегчение -возникает при действии оптимального раздражителя (max ответная реакция)- появл. Центр облегчения.
При действии min раздр. (сниж отв. Рекция) возник окклюзия.
Усвоение и трансформация ритма возбуждения.
Трансформация - изменение частоты нервного импульса при прохождении через нервный центр. Частота может повышаться или понижаться.
Усвоение (танец, режим дня)
Последствие
Запаздание окончания ответной реакции после прекращения действия раздражения. Связано с циркул-й нерв. Имп. По замкн. Цепям нейронов.
Кратковременное (доли секунды)
длительное (секунды)
Ритмическая активность нервных центров.
Увеличение или уменьшение частоты нервных импульсов связанных со свойствами синапса и интегративной длительностью нейронов.
8. Пластичность нервных центров.
Способность перестраивать функциональность свойства для более эффективной регуляции функций,осуществления новых, ранее не свойственных этому центру рефлексов или восстановления фунцкий. В основе пласт синпсов- изменение молл-й стру-ры.
Изменения возбудимости под действием химических веществ.
Высокая чуст-ть к дейст.различ-х ве-в.
Утомляемость нервных центров.
Связана с высокой утомляемостью синапсов. Сниж чувств. Рецепторов.
Общие принципы координационной деятельности ЦНС.
Торможение- особый нер. проц. проявл-ся в уменьшении или полном исчещновении отв. реакции.
Принцип конвергенции
Конвергенция - это схождение импульсов поступающих по различным афферентным путям в каком-либо одном центральном нейроне или нервном центре.
2 . Принцип конвергенции тесно связан с принципом общего конечного пути открытым Шерринктоном. Множество разнообразных раздражителей может вызвать возбуждение одного и того же мотонейрона и одну и ту же двигательную реакцию. Этот принцип обусловлен неодинаковым колличеством афферентных и эфферентных путей.
Принцип дивергенции
Это контактирование одного нейрона с множеством других.
Иррадиация и концентрация возбуждения.
Распространение процесса возбуждения на другие нервные центры, называется иррадиацией (избирательная - в одном направлении, генерализованная - обширная).
Иррадиация через некоторое время сменяется явлением концентрации возбуждения в том же исходном пукте ЦНС.
Процесс иррадиации игрет положительную (формирование новых условных рефлексов) и отрицательную (нарушение тонких взаимоотношений сложившихся между процессами возбуждения и торможения, что приводит к расстройству двигательной деятельности) роли.
Принцип реципрокности (вытормаживает)
Возбуждение одних клеток вызывает торможение других через вставочный нейрон.
Принцип доминанты
Ухтомский сформулировал принцип доминанты как рабочий принцип деятельности нервных центров. Термином доминанта обозначает господствующий очаг возбуждения ЦНС, который определяет текущую деятельность организма.
Принципы доминантного очага :
Повышенная возбудимость нервных центров;
Стойкость возбуждения возбуждения во времени;
Способность к суммации посторонних раздражителей;
Инерция (способность длительного сохранения возбуждения после окончания действия раздражения); способность вызывать сопряженные торможения.

