5-10-2012, 19:14
Функции коры
Лобная доля. Основной функцией лобных долей является контроль произвольных движений, координация двигательных механизмов выразительной речи, а также обеспечение «творческого», или абстрактного, мышления.
Лобную долю можно разделить на пять областей:
- моторную,
- премоторную,
- лобную моторную зрительную,
- моторную область речи Брока
- и лобную ассоциативную области
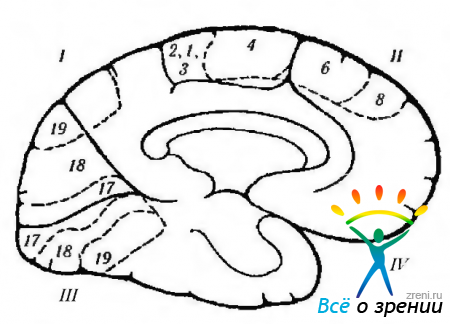
Рис. 4.1.12. Схематическое изображение функций коры большого мозга, локализованных на медиальной поверхности, и их нарушение при различных заболеваниях. сопровождающихся возбуждением или разрушением коры (по Crosby et al., 1962): 1 - теменная доля. Центральная извилина (4). Левая затылочная область: -алексия без аграфии. Задняя кора: -сложность перевода взгляда; -синдром Балинта (двухсторонний), 11 - лобная доля. Дополнительное двигательное поле (6): +поднятие противоположной руки, поворот глаз и головы к руке; + сокращение зрачка; + задержка речи. Лобное двигательное глазное поле (8). Поясная извилина: + расширение зрачка; + слезоотделение. Орбитальная поверхность. III - затылочная доля. Первичная зрительная кора. Контрлатеральное зрение: -правосторонняя гемианопсия. IV-височная доля: паралич движений глаз; ипсилатеральная гемиплегия; гидроцефалия; децеребральная регидность; педункулярный галлюциноз; контрлатеральная гемианопсия; синдром Паринауда. Под поверхностью: - распознание лица. Примечание. Цитоархитектонические поля соответствуют классификации Бродмана. Плюс обозначает результат электростимуляций коры, а минус - результат разрушения поля
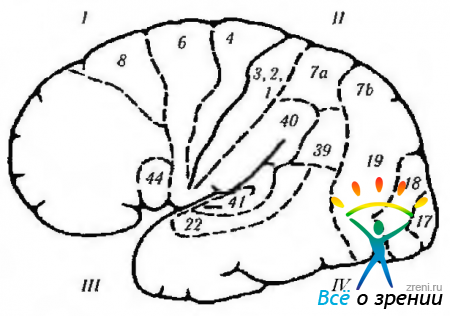
Рис. 4.1.13. Латеральная поверхность мозга (обозначения аналогичны приведенным на предыдущем рис.): 1 - лобная доля. Двигательное поле (4). Премоторное поле (6). Двигательное глазное поле (8): + контрлатеральная саккада; + закрытие противоположного глаза; + гомолатеральное содружественное отклонение глаза. Двигательное речевое поле (44): -двигательная (моторная) афазия. 11 - теменная доля: -(-простые галлюцинозы; -иллюзии; - метаморфопсия: -потеря внимания; -агнозия; - апраксия; -ослабление оптокинетического нистагма; - нижняя квадрантная гемианописия. Супрамаргинальная извилина (40): -амнезия; -жаргонная афазия. Угловая извилина (39): -аграфия; - акалькулия; -утеря способности различать левую и правую сторону; пальцевая агнозия. Соматическая чувствительность. III-височная доля: + зрительные галлюцинации; + deja vu; + нарушение обоняния; + припадки; + автоматизмы; -верхняя квадрианопсия; -потеря памяти; - синдром Клювер-Бюси. Первичная слуховая кора. Вторичная слуховая кора: -слуховая афазия. Planum temporale: - дислексия. IV-затылочная доля (17, 18, 19): + ощущение света; -(-ощущение цветов; -(-ощущение движения; + ощущение линейных изображений. Зрительные ассоциативные поля (18, 19): -(-контрлатеральные движения глаза типа плавного преследования; + вергентные движения; -синдром Антона
Первичная двигательная область (моторная область; 4-е поле по Бродману).
Моторная област ь находится впереди центральной борозды, отделяющей лобную долю от теменной доли. Состоит она из большого количества гигантских пирамидных клеток Беца.
В ней представлены движения, а не индивидуальные мышцы. Клетки, управляющие движениями, расположены таким образом, что движения нижней половины тела представлены в верхней части извилины, а верхней - в нижней. Аксоны нейронов спускаются через внутреннюю капсулу и ножки мозга в виде кортикобульбарных и кортикоспинальных путей.
Удаление двигательной области коры головного мозга ведет к параличу .
Премоторная область (поле б) лежит непосредственно кпереди первичной двигательной коры, занимая заднюю часть трех горизонтально ориентированных лобных извилин.
Формирует она волокна фронтопонтомозжечковых трактов, а также отдает многочисленные волокна базальным ганглиям.
Предполагают, что премоторная кора вовлечена в планирование двигательных действий. Об этом свидетельствует то, что лишь при размышлении о предполагаемом действии нейроны шестого поля возбуждаются. Стимуляция этих нейронов приводит к подъему руки противоположной стороны, а голова и глаза поворачиваются в направлении поднятой конечности. Зрачки расширяются, и нарушается речь (см. рис. 4.1.12, 4.1.13).
Разрушение этой области сопровождается псевдомозжечковой атаксией противоположной стороны . Нарушается также возможность копирования изученных движений (апраксия). При этом отдельные движения не повреждены, но больной не способен производить их в нужной последовательности.
Лобная моторная зрительная область (поле 8 по Бродману).
Эта область (поле 8 и частично 6 и 9) занимает заднюю часть медиальной лобной извилины (gyrus frontalis medius).
Лобная моторная зрительная область важна в произвольных движениях глаза . Некоторые участки определяют конвергенцию, дивергенцию и аккомодацию.
Необходимо отметить, что движение глаз, зрачка и век могут выявляться при электростимуляции почти всех областей полушарий мозга. Электростимуляция лобной моторной глазной области приводит к саккадическим движениям, в отличие от реакции при стимуляции полей 18, 19 и 22 (плавное слежение и вергентные движения). Стимуляция 8-го поля у обезьян сопровождается содружественным контрлатеральным отклонением глаз. При этом расширяется зрачок, а голова и туловище поворачиваются в противоположную сторону. Повреждение лобной доли вызывает обратимую потерю способности к контрлатеральным саккадическим движениям.
Эта область коры отличается многочисленными связями.
Афферентные волокна поступают в зрительную лобную кору от зрительного бугра, а также от верхне-височной, перистриарной и теменной (область 7) коры. Здесь обнаружены нейроны, контролирующие движения глаз.
Эфференты проецируются на базальные ганглии, зрительный бугор, претектальную область (включая ядро зрительного тракта), верхние бугорки четверохолмия и часть ретикулярной формации моста.
В свою очередь необходимо отметить, что лобные зрительные области проецируются на контрлатеральные лобные поля и на области коры с этой же стороны. Обеспечивают они зрительное восприятие (на 7-м поле теменной коры и перистриатной коре).
При проекции на средний мозг волокна прелобной коры пересекают переднюю часть внутренней капсулы и делятся на дорзальные и вентральные пути в ростральной части промежуточного мозга.
Дорзальный трансталамический путь пересекает дорзомедиальное и интраламилярное ядро зрительного бугра и среднюю часть подушки, образуя при этом небольшое количество синаптических связей. Синапсы обнаруживаются также в претектальных ядрах и в верхних бугорках четверохолмия.
Вентральный педункулотегментальный путь проходит в наиболее вентральной части ножки мозга, поставляя волокна к ядрам гипоталамуса, а глубже - к верхним бугоркам четверохолмия. При этом лобные зрительные области обладают топографической проекцией на верхние бугорки четверохолмия. Завершается путь в ретикулярной формации моста.
Третья префронтальная связка возникает в области диэнцефально-мезенцефалического соединения, расположенного около красного ядра. Этот путь проецируется на ядро медиального продольного пучка и промежуточное ядро Кахаля с этой же стороны, а также на ядро Даркшевича и растральную часть глазодвигательного ядра, с обеих сторон.
Различные участки лобной зрительной коры контролируют саккадические движения глаза различной амплитуды, а также направление саккад. Они обеспечивают так называемое «зрительное внимание», направляя глаза к «цели».
В настоящее время выясняется роль лобной доли в функционировании век . Произвольное закрытие век требует расслабления леватора век и сокращения круговой мышцы глаза. Существуют разногласия относительно локализации участков мозга, контролирующих волевые движения века. Эти участки могут располагаться в двигательной премоторной или лобной моторной зрительной областях. Стимуляция этой области ведет к контрлатеральному закрытию века. Односторонние повреждения недоминирующих лобных долей могут сопровождаться двусторонним нарушением закрытия века.
Моторная речевая область Брока (поля 44 и 45 по Бродману).
Эта область располагается в задне-нижней части лобной доли. Она вносит вклад в инициирование речи . Повреждение области Брока приводит к афазии, заключающейся в сложности соединения в определенной последовательности слов для обеспечения связанной речи (экспрессивная афазия). Больные при этом понимают как письменную, так и разговорную речь, но обычно не могут ничего произнести. Нарушена также способность писать, хотя рука может использоваться для выполнения других действий.
Лобная ассоциативная область . Эта важная область в передней половине лобной доли ответственна за инициирование всех типов поведенческо-интеллектуальных, висцеральных и эмоциональных функций. Изменения памяти, интеллекта и индивидуальности наступают при раздражении или массивном разрушении лобных долей. Это особенно выражено в тех случаях. когда поражено доминирующее полушарие.
Обонятельный тракт находится ниже глазничной поверхности лобной доли и является единственным сенсорным путем, достигающим коры без образования синапсов в зрительном бугре. Односторонняя опухоль глазничной поверхности лобной доли (менингиома обонятельной борозды или менингиома в области малого крыла клиновидной кости) может привести к сдавлению обонятельного тракта и зрительного нерва. В результате развивается аносмия (отсутствие обоняния), ипсилатеральная атрофия зрительного нерва и отек диска зрительного нерва с противоположной стороны (синдром Кеннеди Фостера).
Поражение структур лимбической системы в пределах лобной доли, особенно поясной извилины, может вызвать нарушения эмоциональной стороны жизнедеятельности, а также висцеральные признаки, включая беспричинное слезотечение и изменение реакции зрачка на световые стимулы.
Теменная доля (см. рис. 4.1.12, 4.1.13). В целом функция теменных долей связана с чувствительностью, памятью, относящейся к речи и обучению, а также с ориентацией организма в пространстве путем обработки зрительной информации.
В переднем отделе теменной доли находится извилина, расположенная параллельно прецентральной извилине или двигательной области коры. Это постцентральная извилина, или первичная соматосенсорная кора (поля 3, 1 и 2) (см. рис. 4.1.13). Оставшуюся часть теменной доли можно разделить на две дольки - верхнюю и нижнюю. Поля 5 и 7, расположенные на поверхности верхней теменной дольки, связаны с корреляцией соматосенсорной информации, дающей возможность сознательной оценки веса, свойств поверхности, размера и формы предмета.
Нижняя теменная долька (супрамаргинальная и угловая извилины) тесно связана с речевыми механизмами, поскольку разрушение этой области коры в левом полушарии ведет к нарушению восприятия речи.
Роль теменной доли в пространственной ориентации пока еще не изучена. Однако у больных с ее поражением отмечается нарушение этой функции.
Через теменную долю проходят приводящие волокна, соединяющие области 18 и 19 с лобными долями и стволом мозга. Эти пути включают верхний и нижний продольный пучки.
Предполагают, что теменная кора (поле 7) обеспечивает функцию «внимания» к зрительным целям .
Седьмое поле получает афференты от поясной извилины (g. cinguli), от претектальной области и верхних бугорков четверохолмия через ядра зрительного бугра. Обнаружены также афференты, идущие от лобной зрительной области и престриарной коры к коре теменной дольки.
Эфференты проецируются на претектальную область, верхние бугорки четверохолмия, серое вещество, расположенное вокруг сильвиевого водопровода, и на теменные доли двух полушарий мозга.
При движении глаз нейроны нижней теменной доли активизируются, что свидетельствует об их роли в контроле движений глаз, подчиняющихся воле. Активность продолжается на всем протяжении наблюдения за целью.
Клетки 7-го поля не определяют саккадические движения, поскольку их активность при саккадах не увеличивается.
Стимуляция угловой извилины (g. angularis) приводит у экспериментальных животных к содружественному отклонению глаз. Это поля 38 и 39, переходящие вокруг заднего края средней височной извилины (g. temporalis medius). Оперативное удаление этой области коры не приводит к нарушению движения глаз. Двухсторонние повреждения теменной коры могут привести к развитию моторной апраксии глаза.
Поражение теменной доли сопровождается также нарушением зрительных моторных механизмов, обеспечивающих оптокинетический нистагм, в течение которого глаза производят саккады для поиска и фиксации цели.
Другие моторные признаки изолированных повреждений теменной доли включают содружественное отклонение глаз в противоположную сторону от места повреждения при принудительном закрытии век, малую подвижность больного и плохой контроль (непонимание) конечностей противоположной стороны. Трудности в осмыслении характера действия конечностей ведут к конституциональным и глазодвигательным апраксиям. У больных с повреждениями теменной доли могут возникать трудности в процессе перемещения взгляда от одного зрительного объекта к другому, в пределах контрлатеральной половины поля зрения.
Другим признаком дисфункции теменной доли является неспособность фиксировать зрительный объект .
Кроме сенсорных функций, которые локализуются в задней губе центральной борозды Роландо, теменная доля является участком сложного процесса узнавания и осмысления. Диффузное повреждение теменных долей завершается агнозиями, зрительным невниманием, метаморфопсией, фотопсией, галлюцинациями, иллюзиями и алексией (см. рис. 4.1.12, 4.1.13).
Более подробные сведения о последствиях нарушения функции теменной доли приведены в неврологической литературе. Здесь мы приводим лишь некоторые синдромы.
Зрительная агнозия , которая часто развивается при нарушении кровообращения в ЦНС, характеризуется нарушением процесса узнавания. Агнозия может быть полной или частичной. Зрительная агнозия диагностируется в тех случаях, когда больной может идентифицировать объект только тактильно. Часто нарушено узнавание и осмысление зрительных изображений и цветового смысла изображения.
Двухсторонние повреждения теменной доли (чаще встречается при старческом слабоумии, размягчении височно-затылочной и теменной областей коры, ранениях задних отделов обоих полушарий, энцефалите) сопровождаются зрительной агнозией и зрительной атаксией (синдром Балинта).
Зрительная пространственная агнозия характеризуется возникновением трудностей при оценке размера объекта и расстояния до него. Отмечается также появление неправильного решения «синтетических» пространственных задач типа чтения карты. Симультантная агнозия сводится к тому, что у больных сохранена способность узнавания отдельных объектов, но отсутствует способность воспринимать группу объектов (изображений) как целое.
Астерогнозия характеризуется тем, что больной может анализировать объекты по их виду, но не способен охарактеризовать их тактильно. Это развивается вследствие нарушения сложного взаимодействия между тактильной и зрительной информацией в теменных долях коры.
Диффузные поражения теменных долей могут привести к возникновению иллюзий и метаморфопсий. Эти искажения включают микропсию, макропсию. Больные отмечают движение стационарных целей, неправильно оценивают расстояние до объектов. К специальным типам иллюзий относятся папинопсия (нарушение зрительного восприятия в виде сохранения или повторного возникновения зрительного образа после исчезновения предмета из поля зрения), полиопия галлюцинаторная и др.
Иллюзии развиваются в результате диффузных нарушений коры, а галлюцинации и фотопсии - при развитии опухоли или токсическом повреждении коры.
Особенности патологии теменной доли могут зависеть и от того, локализуется ли патологический очаг в доминирующе м (обычно левом) или недоминирующем полушариях. Доминирующая теменная доля ответственна за развитие нарушения речи типа слуховой сенсорной афазии, зрительной агнозии и аграфии.
При повреждении доминирующей угловой извилины (чаще в результате нарушения кровообращения в бассейне угловой артерии) развиваются нистагм, зрительно-агностические расстройства, аграфия, акалькулия (неспособность складывать или вычитать цифры), пальцевая агнозия, невозможность различать правую и левую стороны (синдром Герстманна; синдром угловой извилины). Возможно развитие гомонимной гемианопсии.
При повреждении недоминирующей угловой извилины нарушается взаимодействие больного с окружающей средой. В результате развивается зрительная дезориентация и топографическая агнозия.
Кпереди от угловой извилины и поперек надвисочной борозды находится надкраевая извилина . В доминирующем полушарии эта область является местом встречи зрительных и слуховых компонентов языка.
Повреждение задних теменных ассоциативных областей нередко завершается развитием апраксии (трудность выполнения «тонких» специализированных движений). Это, по-видимому, связано с недопониманием необходимой последовательности движений для выполнения той или иной функции (т. е. больному неясно, что и в какой последовательности должно быть выполнено). У больного отсутствует способность нарисовать простую диаграмму (конструктивная апраксия). Менее серьезной формой этой патологии является осязательная агнозия (неспособность распознать объект при пальпации). Иногда это заболевание называется астерогнозией. При этом нет никаких признаков потери осязательной или проприоцептивной чувствительности, а нарушена интеграция зрительной и соматосенсорной информации. Апраксия и астерогнозия чаще развиваются после повреждения левого полушария головного мозга. Астерогнозия обычно ограничена противоположной стороной тела, в то время как апраксия обычно двухсторонняя.
Зрительная и слуховая информация жестко связана с функцией языка. По этой причине повреждение полей 39 и 40, также как и части слуховой ассоциативной области, обычно приводит к афазии, известной как афазия Вернике (Wernicke). Если повреждение ограничено только угловой извилиной, теряется способность понимать письменный язык (алексия) и способность писать (аграфия). При этом больные понимают речь. Это состояние обычно развивается после травмы.
Интеграция зрительной и соматосенсорной информации важна для формирования «образа тела», т. е. понимания особенностей положения тела в пространстве. Обширные повреждения полей 7, 39 и 40 одного полушария могут привести к развитию «непонимания» или «пренебрежения» противоположной половины тела. Больные не в состоянии перевязать или разбинтовать собственные конечности. Они также не узнают собственные конечности.
Затылочная доля (поля 17, 18 и 19). Вдоль шпорной борозды (fissura calcarina) располагается первичная зрительная область (поле 17). К ней примыкает «оценочная» зрительная область (поле 18), которая, как полагают, преобразует сигналы, получаемые полем 17, в зрительные образы.
Поле 17 затылочной коры является зоной приема и обработки зрительной информации и обладает важными связями с двигательными зрительными участками коры. Именно на этом уровне зрительной коры оценивается значение увиденного. Раздражение этой области вызывает галлюцинации и образы типа сновидений.
Двигательные связи затылочной доли заслуживают внимания и потому, что они вовлечены в оптокинетический нистагм, рефлекс аккомодации и регуляции бинокулярного зрения.
Поля 18 а 19 взаимозависимы и имеют большее количество связей с остальной частью мозга. Предполагают, что область 18 больше заинтересована объединением зрительной информации. полученной в 17-м поле, в то время как поле 19 переводит эту информацию в более сложную мысленную и двигательную деятельность.
Последние анатомические и физиологические исследования выявили, что существуют, по крайней мере, 10 подобластей полей 18 и 19, нейроны которых обладают способностью реагировать на зрительные стимулы. Каждая из этих подобластей связана с 17-м полем. При этом между собой они не связаны. Повреждение этих подобластей не приводит к слепоте. Нарушаются некоторые зрительные функции, например анализ цвета, восприятие цельности объекта и др.
Зрительная ассоциативная кора , расположенная в некоторых областях височной доли, также вовлечена в обработку зрительной информации. В наибольшей степени это относится к полю 21 (в средней височной коре). Именно это поле обеспечивает узнавание образа, а ее повреждение завершается агнозией.
Выше приведенные свойства зрительной коры обеспечиваются и многочисленными горизонтальными и вертикальными связями между нейронами различных слоев зрительной коры. Именно благодаря их наличию отмечается исключительная пластичность зрительной коры при обработке зрительной информации различного типа.
Более подробно структурная организация зрительной коры и ее функции будут приведены в следующем разделе.
Височная доля (поля 41, 22).
Эта часть мозга связана с восприятием слуховых ощущений, участвует в речевой функции посредством слухового контроля речи, а также играет определенную роль в оценке пространства и в функции памяти. Давно установлено, что поперечная височная извилина (поле 41) является первичным слуховым центром. Небольшое поле (22). окружающее эту извилину, рассматривают как «психический», или «оценочный», слуховой центр. Раздражение большей части височной доли, особенно вдоль средней височной извилины, вызывает ощущения, которые больной описывает как слуховые. Больные с поражением «психического поля» височной доли иногда утрачивают способность оценки звуковых тонов. При этом больной не узнает мотивов, а музыкальные звуки воспринимаются им как беспорядочный шум.
Поле 22 левого полушария также связывают с речевой функцией, поскольку поражение этой области приводит к потере способности понимать значение слов. Существуют также данные, свидетельствующие о том, что височная доля связана с «вестибулярной» чувствительностью (чувство равновесия), так как стимуляция задних отделов верхней височной извилины у больного, находящегося в сознании, вызывает головокружение и ощущение вращения.
Глубокие поражения височной доли часто затрагивают самые нижние волокна зрительной лучистости, идущие от наружного коленчатого тела. При этом нарушены функции зрения . Чаще встречается верхняя квадрантная гемианопсия.
При поражении височной или психомоторной доли развивается эпилепсия, сопровождающаяся зрительными галлюцинациями , чувством deja vu и deja pense («уже виденное» - фр.), при которых зрительный опыт и мысли имеют странные дружественные отношения, появляются автоматизмы, сопутствующие амнезии, компульсивное поведение, гнев и беспокойство.
У больных с односторонним повреждением первичной слуховой коры отмечается незначительная потеря слуха. При нарушении слухового узнавания (слуховая агнозия) поражены ассоциативные слуховые поля. Больные, как правило, отмечают, что они могут слышать звуки, но не могут их идентифицировать.
На вентральной поверхности височной доли обнаруживается область, участвующая в узнавании лица. Повреждение этой области (чаще при нарушении кровообращения в области базилярной артерии мозга), что, как правило, распространяется и на смежную затылочную долю (поля 20, 21), обоих полушарий, сопровождается развитием прозопагнозии (неспособность узнавать лица). Вообще, это состояние не сопровождается другими неврологическими признаками. Больной может читать и называть объекты. В случаях, когда больной слышит голос знакомого человека, он может немедленно идентифицировать его.
Оставшаяся часть верхней височной извилины занята областью 22 (слуховая ассоциативная кора), которая получает большое количество афферентов от полей 41 я 42 к отдает волокна теменной и затылочной долям. Область 22 также вовлечена в функции языка. Повреждение верхней части 22-го поля приводит к возникновению трудностей в понимании языка.
Височные доли особенно важны в функции памяти . Например, стимуляция слуховой ассоциативной коры у больных во время нейрохирургических операций вызывает сложные воспоминания как слухового, так и зрительного характера. При обширных повреждениях височной доли возникает нарушение памяти.
Лимбическая доля (рис. 4.1.14).
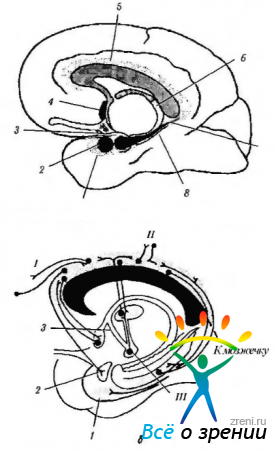
Рис. 4.1.14. Лимбическая система (по Brodsl, 1992): а - расположение структур лимбической системы (1 - энториальное поле; 2 - миндалина; 3 - крючок; 4 - септальное ядро; 5 - поясная извилина; б - свод; 7 - гиппокамп; 8 - парагиппокампальная извилина); б - связи поясной извилины (I - лобная ассоциативная кора; II - теменная и височная ассоциативная кора; III - сосцевидное тело). Поясная извилина связана с ассоциативными полями коры головного мозга и лимбической системой, обеспечивая взаимодействие между ними
Лимбическая система представляет комплекс образований конечного, промежуточного и среднего мозга. Фактически «лимбическая система» совпадает с ранее применявшимся понятием - «обонятельный мозг».
Если новая кора (неокортекс) управляет пространственно-временными взаимоотношениями организма с окружающей средой, а также отвечает за формально-логическое мышление и стереогностические способности, то лимбическая система обусловливает эмоциональные функции и побуждение к действию , а также процессы научения и памяти. Контролирует она и гомеостаз. Немаловажной ее функцией является организация поведенческих реакций индивидуума в ответ на воздействия внешней среды и изменение внутренней среды организма. Эти реакции направлены на сохранение особи и опосредуются через низшие центры промежуточного мозга.
Лимбическая доля имеет какое-то отношение и к функции обоняния, поскольку раздражение или повреждения области гиппокампа сопровождаются ощущением запахов.
Лимбическую долю образуют :
- подмозолистая область (area subcallosa),
- поясная извилина (gyrus cinguli),
- перешеек поясной извилины (isthmus gyri cinguli),
- парагиппокампальная извилина (gyrus parahippocampalis),
- гиппокамп (аммонов рог, зубчатая извилина и основание гиппокампа, или субикулум)
- и диагональная связка Брока.
К подкорковым структурам лимбической системы относятся миндалины (corpus amygdaloideum), септальные ядра и переднее таламическое ядро.
Афферентные и эфферентные связи структур лимбической системы как между собой, так и с другими отделами головного мозга чрезвычайно разнообразны. Наиболее выражены реципрокные связи ее с гипоталамусом.
Гипоталамусе и сосковидные (мамиллярные) тела соединяются с гиппокампом и септальной областью посредством свода, с миндалиной - посредством терминальной полоски и амигдалофугального пучка, а с обонятельным мозгом - посредством медиального пучка переднего мозга. Через гипоталамусе и сосцевидные тела лимбическая система соединена со средним мозгом.
Лимбическая система сообщается с новой корой в области лобной и височной долей. Височные области отвечают главным образом за передачу информации от зрительной, слуховой и соматической коры миндалине и гиппокампу. Лобные области служат основным отделом новой коры, регулирующим лимбические зоны.
Базальные ганглии представляют собой подкорковое скопление нейронов в виде группы ядер (рис. 4.1.15; 4.1.16).
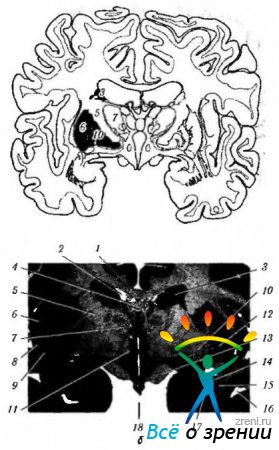
Рис. 4.1.15. Базальные ганглии: а - схематическое изображение расположения структур, относящихся к базальным ганглиям; б - фронтальный срез головного мозга, проходящий через структуры, относящиеся к базальным ганглиям (4 - мозолистое тело; 2- рог бокового желудочка; 3- тело хвостатого ядра; 4-свод; 5-внутренняя капсула; 6 - островок; 7 - зрительный бугор; 8- скорлупа; 9 - ограда; 10- бледный шар; 11 - подбугорная область; 12 - безымянная субстанция: 13 - зрительный тракт; 14 - задняя нижняя часть хвостатого ядра; 15 - миндалина: 16 - височный рог бокового желудочка; 17 - лобный рог бокового желудочка; 18 - третий желудочек)
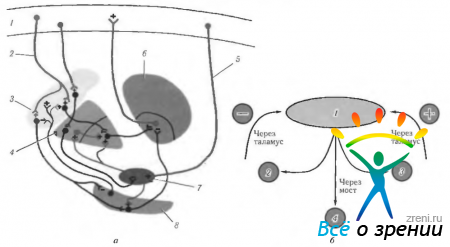
Рис. 4.1.16. Важнейшие афферентные, эфферентные и внутренние связи базальных ганглиев и их связь с мозжечком (разъяснение в тексте): а - (1-двигательная кора; 2 - эфферентные волокна от коры к полосатому телу; 3 - полосатое тело; 4 - бледный шар; 5 - афферентные волокна; 6 - зрительный бугор; 7 - субталамическое ядро; 8 - черная субстанция); б - (1 - кора мозга (двигательная); 2 - базальные ганглии; 3 - мозжечок; 4-двигательные и вставочные нейроны)
Контролируют они двигательную активность тела. Эта функция базальных ганглиев была установлена на основании клинических наблюдений. При их повреждении отмечается нарушение двигательной активности как в виде отсутствия возможности инициировать необходимые движения, так и отсутствия возможности подавлять движения.
При болезни Паркинсона, возникающей при повреждении базальных ганглиев, у больных лицо «невыразительное». Это связывают с нарушением двигательной активности мимических мышц и контроля движений глаза, в частности нарушения саккадических движений. Именно по этой причине мы более подробно рассмотрим нейроанатомию этой области.
Различают три основных скопления подкорковых ядер , которые называются полосатое тело (corpus striatum), ограда (claustrum) и миндалевидное тело (corpus amigdaloideum).
- Corpus striatum состоит из двух частей - хвостатого ядра (nucleus caudatus) и чечевицеобразного ядра (nucleus lentiformis).
Хвостатое ядро лежит выше и медиальнее чечевицеобразного ядра, отделяясь от последнего прослойкой белого вещества, называемой внутренней капсулой (capsula interna). Утолщенная передняя часть хвостатого ядра, его головка (caput nuclei caudati) образуют латеральную стенку переднего рога бокового желудочка, задний же утонченный отдел хвостатого ядра (corpus et cauda nuclei caudati) тянется назад по дну центральной части бокового желудочка. С медиальной стороны nucleus caudatus прилегает к зрительному бугру, отделяясь от него полоской белого вещества (stria terminalis). Спереди и снизу головка хвостатого ядра доходит до переднего перфорированного вещества (substantia perforata anterior). В этом месте головка соединяется с чечевицеобразным ядром (с частью, называемой putamen). Кроме этого широкого соединения обоих ядер с вентральной стороны, имеются еще тонкие полоски серого вещества, располагающиеся вперемешку с белыми пучками внутренней капсулы. Они послужили причиной названия «полосатое тело» (corpus striatum).
Чечевицеобразное ядро (nucleus lentiformis) залегает латерально от хвостатого ядра и зрительного бугра, отделенное от них внутренней капсулой. На горизонтальном разрезе полушария медиальная поверхность чечевицеобразного ядра, обращенная к внутренней капсуле, имеет форму утла с верхушкой, направленной к середине. Передняя сторона угла параллельна хвостатому ядру, а задняя - зрительному бугру. Латеральная поверхность немного выпукла и обращена к латеральной стороне полушария в области островка. Спереди и вентрально, как было уже указано, чечевицеобразное ядро сливается с головкой хвостатого ядра.
На фронтальном разрезе чечевицеобразное ядро имеет форму клина, верхушка которого обращена в медиальную сторону, а основание - латерально. Чечевицеобразное ядро двумя параллельными белыми прослойками (laminae medullares) разделяется на три членика. Латерально расположенный участок темно-серого цвета называется скорлупой (putamen), а два медиальных, более светлых, вместе носят название бледного шара (globus pallidus).
Отличаясь уже по своему макроскопическому виду, бледный шар (globus pallidus) имеет также и гистологическую структуру, которая существенно отличается от строения других частей полосатого тела.
Ввиду всех этих особенностей бледный шар выделяют в особую морфологическую единицу под названием pallidum, тогда как обозначение striatum оставляют только за putamen и nucleus caudatus. Вследствие этого термин «чечевицеобразное ядро» теряет свое прежнее значение и может употребляться только в чисто топографическом смысле. При этом вместо прежнего названия corpus striatum хвостатое и чечевицеобразное ядра именуют стриопаллидарной системой.
Стриопаллидарная система представляет собой главную часть экстрапирамидной системы, а кроме того, она является высшим регулирующим центром вегетативных функций в отношении теплорегуляции и углеводного обмена, доминирующим над подобными же вегетативными центрами в гипоталамусе.
- Ограда (claustrum) представляет собой тонкую пластинку серого вещества, заложенную в области островка, между ним и putamen. От последнего она отделяется прослойкой белого вещества, называемой наружной капсулой (capsula externa).
- Миндалевидное тело
(corpus amygdaloideum) расположено под putamen в переднем конце височной доли. Миндалевидное тело, по-видимому, относится к подкорковым обонятельным центрам и к лимбической системе. В нем оканчивается идущий из обонятельной доли и переднего перфорированного вещества (substantia perforata anterior) пучок волокон, отмеченный при описании зрительного бугра под названием stria terminates.
Роль миндалины у человека недостаточно изучена. В основном, роль миндалины выявлена на основании исследования неврологических больных. Благодаря этим исследованиям установлено, что механическое или электрическое раздражение миндалины у человека часто приводит к возникновению страха или других реакций эмоционального характера. Таким образом, миндалина у человека участвует в процессах эмоционального восприятия . Показано также, что при двухстороннем повреждении миндалин возникает потеря «эмоциональной» памяти, появление «испуганного» выражения лица, нарушение интеграции обонятельной и зрительной информации.
Хорошо известно, что человек обладает способностью установления настроения и эмоционального состояния другого индивидуума. В последнее время выявлено участие миндалины в этом сложном физиологическом процессе. При определении эмоционального состояния человека наш мозг улавливает и анализирует еле заметные изменения выражения лица, и в первую очередь направленность взора. Невральные механизмы, лежащие в основе этих процессов, точно не ясны. Тем не менее установлено, что «точная» установка взгляда, психофизиологически оцениваемая как наличие «внимания», определяется верхней частью височной коры и миндалиной . При исследовании больных аутизмом выявлено, что нередко у них выявляется аномалия миндалины или ее повреждение. Непосредственные механизмы, лежащие в основе социального суждения о других личностях на основании определения направленнности взгляда, остаются в значительной степени неизвестными.
К базальным ганглиям в последнее время относят также черную субстанцию (substantia nigra) и субталамические ядра. Причем в черной субстанции различают 2 части - ретикулярную (pars reticulata) и компактную (pars compacta).
Морфологическими и электрофизиологическими методами показано, что большая часть афферентных сигналов, приходящих к базальным ганглиям, поступает в полосатое тело (см. рис. 4.1.16). Эти сигналы исходят из многих источников, основными из которых являются:
- все области коры больших полушарий;
- внутрипластинчатые ядра зрительного бугра
- черная субстанция (по дофаминэргическому пути).
От внутреннего отдела бледного шара берет начало самый важный из всех эфферентных путей базальных ганглиев. Заканчивается этот путь преимущественно в зрительном бугре и крыше среднего мозга.
Таким образом, базальные ганглии играют роль промежуточного звена в цепи, связывающей двигательные области коры со всеми ее остальными областями. Основной их функцией является «планирование» двигательной активности .
В последние годы установлено, что базальные ганглии совместно с некоторыми корковыми областями также определяют некоторые познавательные функции, включая внимание, память . Понятие «внимание», в широком смысле слова, представляет собой «процесс выбора». Учитывая то, что процесс ориентации в пространстве и выбора цели интереса, участие в котором принимают саккадические движения глаз, связан с базальными ганглиями, становится ясна значительная роль базального комплекса в процессах ориентации посредством саккад. При этом особое место в контроле саккад глаза играет черная субстанция, в частности ее ретикулярная часть. Как анатомически, так и электрофизиологически доказано наличие ее связи с верхними бугорками четверохолмия. Основной функцией нейронов черной субстанции является торможение активности нейронов верхних бугорков четверохолмия, участвующих в генерации саккад.
В двигательной активности глаз принимает участие и хвостатое ядро, имеющее связи с черной субстанцией и наружными бугорками четверохолмия. Возбуждение хвостатого ядра приводит к саккадическим движениям глаза в противоположную от возбуждаемого ядра сторону.
Субталамическое ядро также участвует в движении глаз. На это ядро проецируются бледный шар, кора лобной области. От субталамического ядра отходят волокна к черной субстанции, наружному и внутреннему сегментам бледного шара.
Функция бледного шара в глазодвигательной активности менее ясна. Это связано с тем, что бледный шар имеет большое количество связей между различными ядрами базальных ганглиев и мало связей с нервными центрами, расположенными вне базальных ганглиев. Глазодвигательные нейроны были обнаружены лишь в дорзальной части бледного шара, т. е. в части, принимающей входы от хвостатого ядра.
Роль базальных ганглиев в движении глаз подтверждается многочисленными клиническими наблюдениями. При болезни Паркинсона, сопровождающейся дегенерацией базальных ганглиев и серого вещества среднего мозга и прерывающей фронтомезенцефалические пути, развивается прогрессивный надъядерный паралич. У таких больных скорость саккадических движений глаза уменьшена. Саккады, как правило, гипометрические, что связано с нарушением функции «памяти саккады». Часто движение глаз не может быть завершено в полном объеме. Вертикальные движения глаза повреждаются более значительно, чем горизонтальные. У больных также выявляется осевая ригидность и деменция. Близкие изменения выявлены и при болезни Хунтингтона.
Мозолистое тело (corpus callosum). Мозолистое тело представляет собой большой пучок волокон (состоит примерно из 250 млн волокон), соединяющих между собой оба полушария (рис. 4.1.17).
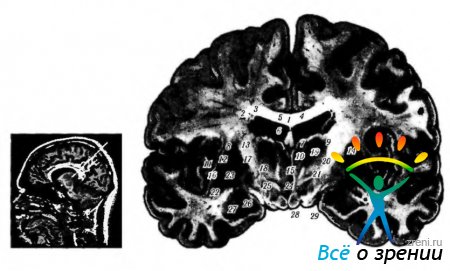
Рис. 4.1.17. Локализация ядер зрительного бугра: а - ядерный парамагнитный резонанс; б - фронтальный срез головного мозга (1 - тело мозолистого тела; 2 - тело хвостатого ядра; 3 - центральная часть бокового желудочка; 4 - прозрачная перегородка; 5 - сосудистое сплетение бокового желудочка; 6 - свод; 7- передняя группа ядер зрительного бугра; 8- наружная капсула; 9 - латеральная группа ядер зрительного бугра; 10- медиальные ядра зрительного бугра: - скорлупа; 12- латеральная медуллярная пластинка; 13 - внутренняя капсула; 14 - ретикулярное ядро зрительного бугра; 15- межталамическая спайка; 16 - латеральная часть бледного шара; 17 - медиальная медуллярная пластинка; 18 - мамиллоталамический пучок; 19 - HI поле Фореля; 20 - гопа incerta; 21 - Н2 поле Фореля; 22 - ограда; 23-медиальная часть бледного шара; 24 - третий желудочек; 25 - ядра гипоталамуса; 26-зрительный тракт; 27 - миндалевидное тело; 28-сосковидное тело; 29-основание ножек мозга)
Основной функцией этого пучка является передача информации из одного полушария в другое.
Основными функциями мозолистого тела являются :
- Корреляция изображений на правой и левой половинах поля зрения.
- Интеграция ощущений, поступающих от парных конечностей, необходимая для обучения двигательной координации.
- Интеграция процессов внимания и активации в полушариях.
У больных, у которых с терапевтической целью была произведена перерезка мозолистого тела, нарушений функций мозга в повседневной жизни не наблюдалось. Они выявлялись лишь с помощью специальных тестов. У других больных развиваются иногда довольно значительные нарушения поведения. Наиболее часто проявляется это «словесной слепотой», «словесной глухотой» и потерей координации движений правой и левой конечностей.
Продолжение в следующей статье: Анатомия головного мозга? Часть 3
И в глазах у всей столицы
Петушок спорхнул со спицы,
К колеснице полетел
И царю на темя сел,
Встрепенулся, клюнул в темя
И взвился... и в то же время
С колесницы пал Дадон —
Охнул раз, — и умер он.
Кажется, вы уже поняли, о чем пойдет речь в очередном материале цикла «Как устроен мозг». Мы уже рассказали о лобных долях, височных и теменных, теперь переходим к теменным долям. Они же Lobus parietalis на привычной медикам латыни.
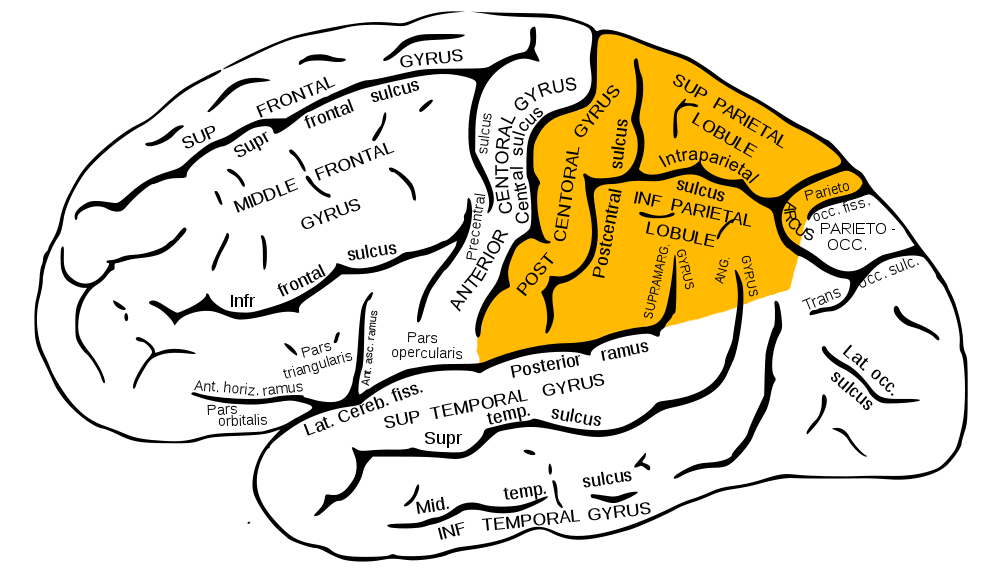
Теменные доли показаны жёлтым
Теменная доля находится чуть выше затылка и «состоит» из трех извилин: одна вертикальная - задняя центральная (самый старый отдел) и две горизонтальные - верхнетеменная (новый) и нижнетеменная (более новый).
Подобно строению лобных долей, на центральной передней извилине теменной «спроецированы» части тела человека: нижняя треть - лицо, средняя треть - рука и туловище, верхняя - нога. Не забываем, что доля «двоится» поэтому одна её половина отвечает за одну (противоположную) половину тела.
Структура теменных долей
Кроме этого, в верхней теменной извилине находятся центры, отвечающие за сложные виды глубокой чувствительности: мышечно-суставным, двумерно-пространственным чувством (определение цифры, буквы, фигуры, начерченные карандашом или другим тупым предметом на коже человека), чувством веса и объема движения, чувством распознавания предметов на ощупь.
В нижней теменной доле расположены центры праксиса, то есть ставшие «автоматическими» в процессе повторений и упражнений движения, которые вырабатываются в процессе обучения и постоянной практики, например, ходьба, еда, одевание и прочее.
Теменная доля участвует в переработке и осознанном восприятии соматосенсорной (от рецепторов мышц, кожи, суставов и внутренних органов) информации, которая влияет на произвольные движения.
Поражения верхней теменной дольки сопровождаются развитием нарушения способности познавать предметы путем их ощупывания с закрытыми глазами. Больные описывают отдельные качества предмета, но не могут синтезировать его образ.
При поражении нижней теменной дольки нарушается ощущение схемы тела. Человек не способен осознавать, где правая, а где левая сторона, не узнает собственных пальцев кисти. Другим видом расстройства является неосознание своего дефекта (больной уверяет, что он двигает парализованными конечностями). У таких больных может возникать псевдополимелия — ощущение лишней конечности или частей тела. Такие пациенты могут самостоятельно удалять «мешающую» конечность или способствовать её ампутации.
При поражении коры угловой извилины больной теряет ощущение пространственного восприятия окружающего мира, положения собственного тела и взаимосвязей его частей. Это сопровождается разнообразными психопатологическими симптомами: деперсонализацией, дереализацией. Они могут наблюдаться при условии полного сохранения сознания и критического мышления.
При поражениях теменной доли доминантного полушария у человека появляется дислексия - неспособность читать, различать право и лево, а также дискалькулия - неспособность к арифметике. Примечательно, что зачастую дискалькулия является самостоятельным недугом, а не следствием неврологических или психологических проблем. Кроме этих проблем возможна и апраксия - нарушение или невозможность выполнить какое-то целенаправленное действие (ну, например, взять стакан и выпить) при сохранении элементарных составляющих действия.
Анастасия Шешукова
Затылочная доля занимает задние отделы полушарий. На выпуклой поверхности полушария затылочная доля не имеет резких границ, отделяющих ее от теменной и височной долей, за исключением верхнего отдела теменно-затылочной борозды, которая, располагаясь на внутренней поверхности полушария, отделяет теменную долю от затылочной. Борозды и извилины верхнебоковой поверхности затылочной доли непостоянны и имеют вариабельное строение. На внутренней поверхности затылочной доли имеется шпорная борозда, которая отделяет клин (треугольной формы дольку затылочной доли) от язычковой извилины и затылочно-височной извилины.
Функция затылочной доли связана с восприятием и переработкой зрительной информации, организацией сложных процессов зрительного восприятия. При этом в области клина проецируется верхняя половина сетчатки глаза, воспринимающая свет от нижних полей зрения; в области язычковой извилины находится нижняя половина сетчатки глаза, воспринимающая свет от верхних полей зрения.
Островок
Островок, или так называемая закрытая долька, находится в глубине боковой борозды. От примыкающих соседних отделов островок отделен круговой бороздой. Поверхность островка разделена его продольной центральной бороздой на переднюю и заднюю части. В островке проецируется анализатор вкуса.
Лимбическая кора
На внутренней поверхности полушарий над мозолистым телом находится поясная извилина. Эта извилина перешейком позади мозолистого тела переходит в извилину около морского конька - парагиппокампову извилину. Поясная извилина вместе с парагиттпокамповой извилиной составляют сводчатую извилину.
Внутренняя и нижняя поверхности полушарий объединяются в так называемую лимбическую (краевую) кору вместе с миндалевидным ядром из группы подкорковых ядер, обонятельным трактом и луковицей, участками лобных, височных и теменных долей коры больших полушарий, а также с подбугорной областью и ретикулярной формацией ствола. Лимбическая кора объединяется в единую функциональную систему - лимбико-ретикулярный комплекс. Основной функцией этих отделов мозга является не столько обеспечение связи с внешним миром, сколько регуляция тонуса коры, влечений и аффективной жизни. Они регулируют сложные, многоплановые функции внутренних органов и поведенческие реакции. Лимбико-ретикулярный комплекс - важнейшая интегративная система организма. Лимбическая система имеет также важное значение в формировании мотиваций. Мотивация (или внутреннее побуждение) включает в себя сложнейшие инстинктивные и эмоциональные реакции (пищевые, оборонительные, половые). Лимбическая система принимает участие также в регуляции сна и бодрствования.
Лимбическая кора выполняет также важную функцию обоняния. Обоняние - восприятие находящихся в воздухе химических веществ. Обонятельный мозг человека обеспечивает обоняние, а также организацию сложных форм эмоциональных и поведенческих реакций. Обонятельный мозг является частью лимбической системы.
Обонятельный мозг состоит из двух отделов - периферического и центрального. Периферический отдел представлен обонятельным нервом, обонятельными луковицами, первичными обонятельными центрами. Центральный отдел включает извилину морского коня - гиппокамп, зубчатую и сводчатую извилины.
Рецепторный аппарат обоняния расположен в слизистой оболочке носа. По системе нервных проводников информация с рецепторов передается в корковый отдел обонятельного анализатора (рис. 8).
Рис. 8. Обонятельный анализатор (схема):
1 - обонятельный эпителий, биполярные обонятельные клетки; 2 - обонятельная луковица; 3 - обонятельный тракт; 4 - первичные обонятельные центры; 5 - зрительный бугор; 6 - корковый обонятельный центр; 7 - мозолистое тело
Корковый отдел обонятельного анализатора находится в поясной извилине, извилине морского коня и в крючке морского коня которые вместе составляют замкнутую кольцевидную область. Периферический отдел обонятельного анализатора связан с корковыми областями обоих полушарий.
Физиологический механизм восприятия запахов обонятельным анализатором окончательно не ясен. Существуют две основные гипотезы, с разных позиций объясняющие природу этого процесса. Согласно одной из гипотез, взаимодействие между молекулами пахучего вещества и хеморецепторами происходит по типу ключа и замка, т.е. типу молекулы соответствует специальный рецептор. Другая гипотеза базируется на предположении о том, что молекулы пахучего вещества имеют определенную волну колебания, на которую “настроены” обонятельные рецепторы. Молекулы, имеющие сходные колебания, должны иметь общую волну и соответственно давать близкие запахи.
Термин “обонятельный мозг” применительно к физиологии человека несколько условен и не раскрывает полностью его многогранной и универсальной функции. “Размещение” центрального звена обонятельного мозга в больших полушариях неслучайно и является результатом той огромной “информационной” роли, которую играло обоняние в процессе эволюции при адаптации к внешней среде и регуляции сложных поведенческих реакций. Добывание пищи, выбор особи противоположного пола, забота о потомстве, целостности территории, организация групповых сообществ внутри вида - все эти повседневные функции у многих животных выполняются при непосредственном участии тонко сконструированной системы обонятельной рецепции и основанной на этом способности ряда животных посылать во внешнюю среду тонкие дифференцированные специфические пахучие вещества - сигналы-информаторы.
Универсальные формы поведенческих реакций у животных проявляющиеся в повседневной заботе о месте обитания, о потомстве, создают впечатление о наделенности их разумом. Кажущийся интеллект - просто результат реакции на внешние стимулы. Однако сами эти стимулы и реакции на них прекрасно соответствуют биологическим потребностям животных.
В жизни людей обоняние потеряло то биологическое информационное значение, которое оно имело у животных. Обонятельная система человека предназначена как для выполнения узкой, “своей” функции, так и для своего рода “зарядки” эмоций. О силе воздействия запахов на эмоциональную сферу, о том, что они являются важнейшим “пищевым субстратом эмоций”, хорошо известно с давних времен истории человечества.
Острота обоняния человека может варьировать. Как правило, эти вариации незначительны, однако в отдельных случаях острота обоняния может быть очень высокой (дегустаторы парфюмерной промышленности).
Поскольку обонятельный анализатор играет важную роль в регуляции эмоций, его центральный отдел относят к лимбической системе, образно названной “общим знаменателем” для множества эмоциональных и висцеросоматических реакций организма.
Большие полушария головного мозга
представляют собой самый массивный отдел головного мозга. Они покрывают мозжечок и ствол мозга. Большие полушария составляют примерно 78 % общей массы мозга.
В процессе онтогенетического развития организма большие полушария головного мозга развиваются из конечного мозгового пузыря нервной трубки, поэтому данный отдел головного мозга называется также конечным мозгом.
Большие полушария головного мозга разделены по средней линии глубокой вертикальной щелью на правое и левое полушария. В глубине средней части оба полушария соединены между собой большой спайкой - мозолистым телом. В каждом полушарии различают доли: лобную , теменную , височную , затылочную .
Доли мозговых полушарий отделяются одна от другой глубокими бороздами. Наиболее важны три глубокие борозды: центральная (роландова), отделяющая лобную долю от теменной; боковая (сильвиева), отделяющая височную долю от теменной, и теменно-затылочная, отделяющая теменную долю от затылочной на внутренней поверхности полушария.
Каждое полушарие имеет верхнебоковую (выпуклую) - конвекситальную, нижнюю - базальную и внутреннюю - медиальную поверхность. Каждая доля полушария имеет мозговые извилины, отделенные друг от друга бороздами. Сверху полушарие покрыто корой - тонким слоем серого вещества, которое состоит из нервных клеток.
Кора головного мозга - наиболее молодое в эволюционном отношении образование центральной нервной системы. У человека она достигает наивысшего развития. Кора головного мозга имеет огромное значение в регуляции жизнедеятельности организма, в осуществлении сложных форм поведения и становлении нервно-психических функций.
Под корой находится белое вещество полушарий, оно состоит из отростков нервных клеток - проводников. Из-за образования мозговых извилин общая поверхность коры головного мозга значительно увеличивается. Общая площадь коры полушарий составляет 1200 см2, причем 2/3 ее поверхности находится в глубине борозд, а 1/3 - на видимой поверхности полушарий. Каждая доля мозга имеет различное функциональное значение.
Кора головного мозга состоит из 4 долей, отделенных друг от друга бороздами. Главные борозды, отделяющие лобные, теменные и височные доли – Роландова и Сильвиева.
Доли коры головного мозга :
лобная (frontal ),
теменная (parietal ),
затылочная (occipital ),
височная (temporal ),
В связи с долями говорят о локальных системах мозга.
ЛОКАЛЬНЫЕ СИСТЕМЫ МОЗГА:
Затылочные отделы мозга выполняют функции организации зрительного восприятия. Первичные зоны затылочной коры - элементарные функции зрения.
Вторичные отделы затылочной коры - оптико-гностические функции.
Височные отделы мозга "отвечают" за организацию слухового восприятия. Первичные зоны височной коры - элементарные функции слуха.
Вторичные зоны височной коры - акустико-гностические функции.
Сенсомоторные и премоторные отделы мозга - организация движений. 1. Постцентральные отделы мозга – афферентная организация движений. 2. Премоторные зоны коры - эфферентная организация (программирование) движения.
3. Моторные зоны коры – двигательный анализатор, выполнение двигательной программы. Лобные доли мозга (префронтальные отделы) выполняют регуляцию психической деятельности, т.е. регуляцию состояний активности, произвольных движений и действий, когнитивных процессов и эмоционально-личностной сферы, отвечают в целом за мышление, интеллектуальную деятельность. Теменные отделы мозга представлены первичными зонами (все виды кожно-кинестетической чувствительности), вторичными зонами (зрительно-пространственные представления, представления о схеме тела, соматогнозис, стереогнозис) и третичными зонами.
Лобная доля занимает передние отделы полушарий. От теменной доли она отделяется центральной бороздой, от височной - боковой бороздой. В лобной доле имеются четыре извилины: одна вертикальная - прецентральная и три горизонтальные - верхняя, средняя и нижняя лобные извилины.
Извилины отделены друг от друга бороздами. На нижней поверхности лобных долей различают прямую и орбитальную извилины. Прямая извилина залегает между внутренним краем полушария, обонятельной бороздой и наружным краем полушария.
В глубине обонятельной борозды лежат обонятельная луковица и обонятельный тракт. Лобная доля человека составляет 25-28% коры, средняя масса лобной доли 450 г.
Функция лобных долей связана с организацией произвольных движений, двигательных механизмов речи, регуляцией сложных форм поведения, процессов мышления. В извилинах лобной доли сконцентрировано несколько функционально важных центров. Передняя центральная извилина является «представительством» первичной двигательной зоны со строго определенной проекцией участков тела. Лицо «расположено» в нижней трети извилины, рука в средней трети, нога - в верхней трети. Туловище представлено в задних отделах верхней лобной извилины. Таким образом, человек спроецирован в передней центральной извилине вверх ногами и вниз.
Передняя центральная извилина вместе с прилегающими задними отделами лобных извилин выполняет очень важную в функциональном отношении роль. Она является центром произвольных движений. В глубине коры центральной извилины от так называемых пирамидных клеток -центрального двигательного нейрона - начинается основной двигательный путь - пирамидный, или кортикоспинальный, путь. Периферические отростки двигательных нейронов выходят из коры, собираются в единый мощный пучок, проходят центральное белое вещество полушарий и через внутреннюю капсулу входят в ствол мозга; в конце ствола мозга они частично перекрещиваются (переходя с одной стороны на другую) и затем спускаются в спинной мозг . Эти отростки заканчиваются в сером веществе спинного мозга. Там они вступают в контакт с периферическим двигательным нейроном и передают ему импульсы из центрального двигательного нейрона. По пирамидному пути передаются импульсы произвольного движения.
В задних отделах верхней лобной извилины располагается также экстрапирамидный центр коры, тесно связанный анатомически и функционально с образованиями так называемой экстрамирамидной системы. Экстрапирамидная система - двигательная система, помогающая осуществлению произвольного движения. Это система «обеспечения» произвольных движений. Будучи филогенетически более старой, чем пирамидная система, экстрапирамидная система у человека обеспечивает автоматическую регуляцию «заученных» двигательных актов, поддержание общего мышечного тонуса, «готовность» периферического двигательного аппарата к совершению движения, перераспределение мышечного тонуса при движениях. Кроме того, она участвует в поддержании нормальной позы.
В заднем отделе средней лобной извилины находится лобный глазодвигательный центр, осуществляющий контроль за содружественным, одновременным поворотом головы и глаз (центр поворота головы и глаз в противоположную сторону). Раздражение этого центра вызывает поворот головы и глаз в противоположную сторону. Функция этого центра имеет огромное значение в осуществлении так называемых ориентировочных рефлексов, имеющих очень важное значение для сохранения жизни животных.
В заднем отделе нижней лобной извилины находится моторный центр речи (центр Брока).
Лобный отдел коры больших полушарий принимает также активное участие в формировании мышления, организации целенаправленной деятельности, перспективном планировании.
Височная доля занимает нижнебоковую поверхность полушарий. От лобной и теменной долей височная доля отграничивается боковой бороздой.
На верхнебоковой поверхности височной доли имеются три извилины - верхняя , средняя и нижняя . Верхняя височная извилина находится между сильвиевой и верхней височной бороздами, средняя - между верхней и нижней височной, бороздами, нижняя - между нижней височной бороздой и поперечной мозговой щелью. На нижней поверхности височной доли различают нижнюю височную извилину, боковую затылочновисочную извилину, извилины гиппокампа (ноги морского коня).
Функция височной доли связана с восприятием слуховых, вкусовых, обонятельных ощущений, анализом и синтезом речевых звуков, механизмами памяти. Основной функциональный центр верхнебоковой поверхности височной доли находится в верхней височной извилине. Здесь располагается слуховой, или гностический, центр речи (центр Вернике).
В верхней височной извилине и на внутренней поверхности височной доли находится слуховая проекционная область коры. Обонятельная проекционная область находится в гиппокамповой извилине, особенно в ее переднем отделе (так называемом крючке). Рядом с обонятельными проекционными зонами находятся и вкусовые.
Височные доли играют важную роль в организации сложных психических процессов, в частности памяти.
Теменная доля занимает верхнебоковые поверхности полушария. От лобной теменная доля спереди и сбоку ограничивается центральной бороздой, от височной снизу - боковой бороздой, от затылочной - воображаемой линией, проходящей от верхнего края теменно-затылочной борозды до нижнего края полушария.
На верхнебоковой поверхности теменной доли имеются три извилины: одна вертикальная - задняя центральная и две горизонтальные - верхнетеменная и нижнетеменная. Часть нижнетеменной извилины, огибающей задний отдел латеральной борозды, называют надкраевой (супрамаргинальной), а часть, окружающую верхнюювисочную извилину , узловой (ангулярной) областью.
Теменная доля, как и лобная, составляет значительную часть полушарий головного мозга . В филогенетическом отношении в ней выделяют старый отдел - заднюю центральную извилину, новый - верхнетеменную извилину и более новый - нижнетеменную извилину. Функция теменной доли связана с восприятием и анализом чувствительных раздражений, пространственной ориентацией. В извилинах теменной доли сконцентрировано несколько функциональных центров.
В задней центральной извилине спроецированы центры чувствительности с проекцией тела, аналогичной таковой в передней центральной извилине. В нижней трети извилины спроецировано лицо, в средней трети - рука, туловище, в верхней трети - нога. В верхней теменной извилине находятся центры, ведающие сложными видами глубокой чувствительности: мышечно-суставным, двухмерно-пространственным чувством, чувством веса и объема движения, чувством распознавания предметов на ощупь.
Таким образом, в теменной доле локализуется корковый отдел чувствительного анализатора.
В нижней теменной доле расположены центры праксиса. Под праксисом понимаются ставшие автоматизированными в процессе повторений и упражнений целенаправленные движения, которые вырабатываются в процессе обучения и постоянной практики в течение индивидуальной жизни.
Ходьба, еда, одевание, механический элемент письма, различные виды трудовой деятельности (например, движения водителя по управлению автомобилем, косьба и пр.) являются праксисом.
Праксис - высшее проявление свойственной человеку двигательной функции. Он осуществляется в результате сочетанной деятельности различных территорий коры головного мозга .
Затылочная доля занимает задние отделы полушарий. На выпуклой поверхности полушария затылочная доля не имеет резких границ, отделяющих ее от теменной и височной долей, за исключением верхнего отдела теменно-затылочной борозды, которая, располагаясь на внутренней поверхности полушария, отделяет теменную долю от затылочной.
Борозды и извилины верхнебоковой поверхности затылочной доли непостоянны и имеют вариабельное строение.
На внутренней поверхности затылочной доли имеется шпорная борозда, которая отделяет клин (треугольной формы дольку затылочной доли) от язычковой извилины и затылочно-височной извилины.
Функция затылочной доли связана с восприятием и переработкой зрительной информации , организацией сложных процессов зрительного восприятия. При этом в области клина проецируется верхняя половина сетчатки глаза, воспринимающая свет от нижних полей зрения; в области язычковой извилины находится нижняя половина сетчатки глаза, воспринимающая свет от верхних полей зрения.
Островок , или так называемая закрытая долька , находится в глубине боковой борозды. От примыкающих соседних отделов островок отделен круговой бороздой.
Поверхность островка разделена его продольной центральной бороздой на переднюю и заднюю части. В островке проецируется анализатор вкуса .
Обонятельный анализатор
Нервные клетки , воспринимающие обонятельные раздражения, расположены в слизистой оболочке верхних отделов носовой полости. Отсюда аксоны этих клеток идут в полость черепа и поступают в обонятельные луковицы. От них нервные волокна направляются к височной доле (внутренняя поверхность) , где находятся нервные клетки обонятельного анализатора .
Вкусовой анализатор
Начинается этот анализатор в нервных окончаниях вкусовых сосочков языка, которые представлены в них вкусовыми луковицами. Отходящие от вкусовых луковиц нервные волокна идут в головной мозг и оканчиваются, как и нервы обонятельного анализатора, на внутренней поверхности височной доли .
Мозолистое тело - дугообразная тонкая пластинка, филогенетически молодая, соединяет срединные поверхности обоих полушарий . Удлиненная средняя часть мозолистого тела сзади переходит в утолщение, а спереди искривляется и дугообразно загибается вниз.
Мозолистое тело соединяет филогенетически наиболее молодые участки полушарий и играет важную роль в обмене информацией между ними.
МОЗГОВОЙ СТВОЛ, ИЛИ СТВОЛ ГОЛОВНОГО МОЗГА -
традиционно выделяющаяся система отделов головного мозга, представляющая собой протяжённое образование, продолжающее спинной мозг.
В ствол всегда включают продолговатый мозг,варолиев мост, а такжесредний мозг. Часто в него включаютмозжечок, иногда -промежуточный мозг.
Продолговатый мозг -
отдел головного мозга. Встречается также традиционное название bulbus (луковица, из-за формы этого отдела).
Продолговатый мозг входит в ствол головного мозга.
Снаружи на вентральной(лицевой) стороне расположены пирамиды (в них пролегает кортикоспинальный тракт - путь от коры к двигательным нейронамспинного мозга) и оливы (внутри них находятся ядра нижней оливы, связанные с поддержанием равновесия). Надорсальнойстороне: тонкий и клиновидный пучки, оканчивающиеся бугорками тонкого и клиновидного ядер (переключают информацию глубокой чувствительности нижней и верхней половин тела соответственно), нижняя половина ромбовидной ямки, являющейся дном четвертого желудочка, и отделяющие ее веревочные тела, или нижние ножки мозжечка.
Внутри расположены также ядра от VIII до XII (и одно из ядер VII) черепномозговых нервов, часть ретикулярной формации, медиальная петля и другие восходящие и нисходящие пути.
Имеет вид усеченного конуса.
Благодаря исследованиям таких учёных, как Р.Магнус и И. Ф. Клейн было установлено наличие в продолговатом мозге сложной системы рефлекторных центров, обеспечивающих определённое положение в теле за счёт статических и статико-кинетических рефлексов. Эти рефлексы, собственно, представляют собой механизмы перераспределения мышечного тонуса таким образом, что сохраняется удобная для животного поза (позно-тонические рефлексы) или возвращение в данную из неудобной (выпрямительные рефлексы), а также обеспечивается сохранение равновесия при ускорении (стато-кинетические рефлексы). Осуществление данных рефлексов происходит с участием таких формаций ствола как ретикулярная формация, красное ядро и вестибулярные ядра.
Ретикулярная формация - это формация, идущая от спинного мозга к таламусув ростральном (к коре) направлении. Кроме участия в обработке сенсорной информации, ретикулярная формация оказывает активизирующее воздействие на кору головного мозга, контролируя, таким образом, деятельность спинного мозга. Впервые механизм воздействия ретикулярной формации на мышечный тонус был установлен Р.Гранитом (R.Granit): он показал, что ретикулярная формация способна изменять активность γ-мотонейронов, в результате чего ихаксоны(γ-эфференты) вызывают сокращение мышечных веретён, и, как следствие, усиление афферентной импульсции от мышечных рецепторов. Эти импульсы, поступая в спинной мозг, вызывают возбуждение α-мотонейронов, что и является причиной тонуса мышц.
Установлено, что участие в выполнении данной функции ретикулярной формации принимают два скопления нейронов: нейроны ретикулярной формации моста и нейроны ретикулярной формации продолговатого мозга. Поведение нейронов ретикулярной формации продолговатого мозга схоже с поведением нейронов ретикулярной формации моста: они вызввают активацию α-мотонейронов мышц-сгибателей и, следовательно, тормозят активность α-мотонейронов мышц-разгибателей. Нейроны ретикулярной формации моста действуют ровно наоборот, возбуждают α-мотонейроны мышц-разгибателей и тормозят активность α-мотонейронов мышц-сгибателей. Реткулярная формация имеет связь с мозжечком (часть информации от него идёт к нейронам продолговатого мозга (от ядер пробковидного и шаровидного мозжечка), а от шатра - к нейронам моста) и с корой головного мозга, от которой получает информацию. Это позволяет утверждать, что ретикулярная формация является коллектором неспецеффического сенсорного потока, возможно участвующим в регуляции мышечной активности.
Важное функциональное значение имеет ретикулярная , или сетевидная формация ствола мозга, которая развивается в связи с возникновением системы блуждающего, вестибулярного и тройничного нервов .
Сетевидная формация состоит из различных по величине и форме нервных клеток, а также из густой сети нервных волокон, идущих в различных направлениях и располагающихся главным образом вблизи желудочковой системы. Ретикулярной формации придается основное значение в корково-подкорковых взаимоотношениях. Она располагается в средних этажах продолговатого мозга ,гипоталамусе , сером веществе покрышки среднего мозга , варолиевом мосту .
К сетевидной формации подходят многочисленные коллатерали от всех афферентных (чувствительных) систем. Через эти коллатерали любое раздражение с периферии, направляясь в определенные участки коры по специфическим путям нервной системы, достигает и сетчатой формации. Неспецифические восходящие системы (т. е. пути от ретикулярной формации) обеспечивают возбуждение коры больших полушарий, активацию ее деятельности.
Наряду с восходящими неспецифическими системами в стволе мозга проходят нисходящие неспецифические системы, которые воздействуют на спинальные рефлекторные механизмы.
Ретикулярная формация тесно связана с лимбической системой, а также с корой больших полушарий. Благодаря этому формируется функциональная связь между высшими отделами центральной нервной системы и стволом головного мозга . Эта система получила название лимбико-ретикулярного комплекса, или лимбико-ретикулярной оси. Этот сложный структурно-функциональный комплекс обеспечивает интеграцию важнейших функций, в осуществлении которых участвуют различные отделы головного мозга.
Известно, что бодрствующее состояние коры обеспечивается специфическими и неспецифическими системами. Реакция активации поддерживается постоянным поступлением импульсов с рецепторов слухового , зрительного , обонятельного, вкусового и кожно-кинестетического анализаторов. Эти раздражения передаются по специфическим афферентным путям в различные участки коры. От всех поступающих в зрительный бугор , а затем в кору больших полушарий афферентных путей отходят многочисленные коллатерали к ретикулярной формации, чем и обеспечивается ее восходящая активирующая деятельность.
В свою очередь ретикулярная формация получает импульсы из мозжечка , подкорковых ядер, лимбической системы , которые обеспечивают эмоционально-адаптивные поведенческие реакции, мотивационные формы поведения. Однако степень обеспечения неспецифической системой адаптивных безусловнорефлекторных реакций у человека и у животных различна. Если у животных подкорковые образования и лимбическая система имеют ведущее значение в выполнении жизненно важных потребностей организма для его выживания в окружающей среде, то у человека в связи с доминированием коры деятельность глубинных структур мозга (подкорковых образований, лимбической системы, ретикулярной формации) в большей степени, чем у животного, подчинена коре больших полушарий . Ретикулярной формации принадлежит важная роль в регуляции мышечного тонуса. Регуляция мышечного тонуса проводится по двум видам ретикулоспинальных путей. Быстро проводящий ретикулоспинальный путь регулирует быстрые движения; медленно проводящий ретикулоспинальный путь - медленные тонические движения.
Ретикулярная формация продолговатого мозга принимает участие в возникновении децеребрационной ригидности. При перерезке ствола мозга выше продолговатого мозга понижается активность нейронов, оказывающих тормозящее влияние на мотонейроны спинного мозга, что приводит к резкому повышению тонуса скелетной мускулатуры.
Функции продолговатого мозга
Защитные рефлексы (например кашель, чиханье).
Жизненно важные рефлексы (например дыхание).
Регулирование сосудистого тонуса.
Рефлекторные центры продолговатого мозга:
защитная (кашель, чиханье и тому подобное)
центры регуляции тонуса скелетных мышц для поддержания позы человека.
укорочение или удлинение времени спинального рефлекса
пищеварение
сердечная деятельность
Варолиев мост
Варолиев мост (от имени Констанзо Варолия), или мост - отдел головного мозга, является вместе с мозжечком частьюзаднего мозга. Принадлежитстволу мозга,
В базисе моста проходят нисходящие пути: кортикоспинальный пирамидный тракт, кортикобульбарный, кортикомостовый тракты.
Сре́дний мозг (лат.Mesencephalon) - отдел головного мозга, древний зрительный центр. Включен вствол головного мозга.
Функции среднего мозга
1. двигательная,
2.сенсорная (зрение, слух),
3. Регулирующая акты жевания и глотания,

