Smegenų kilmė Saveliev Sergejus Viačeslavovičius
§ 37. Roplių smegenų asociacinis centras
Atsižvelgiant į pastato bendrąjį planą nervų sistema, turėtume atskirai pasilikti prie naujų smegenų organizavimo ir veikimo principų, pirmą kartą pritaikytų ropliams. Archajiškų amniono nervų sistema tapo loginis vystymasis sėkmingo amfibijos dizaino pastatai. Tačiau varliagyvių smegenys praktiškai atliko sudėtingo refleksinio aparato funkciją, o jos intelektualinės galimybės liko nepanaudotos. Varliagyvių evoliuciją lėmė raumenys, dantys, linijiniai matmenys ir dauginimosi mastai.
Iš visų žinduolių žmonės yra labiausiai išsivysčiusi ir išsivysčiusios smegenys. Tačiau visų stuburinių gyvūnų smegenys susideda iš 3 pagrindinių struktūrinių vienetų: smegenų pusrutulių, smegenų kamieno ir smegenėlių. Tinklo formavimas yra pagrindinė informacijos mainų sritis, kuri yra smegenų energijos šaltinis. Šiame tinkle nervinių skaidulų kiekvienas neuronas gali būti prijungtas prie kitų kryžminio ryšio formatų, kurie nuolat gauna žinutes iš jutimo organų, kurie savo ruožtu skleidžia elektrinį aktyvumą per smegenų žievę.
Jis palaiko sąmonę. Jei susiūta juosta sumažina jų aktyvumą, žmogus gali užmigti arba patirti haliucinacijų, matyti, girdėti ir jausti nerealius pojūčius. Sensorinio nepritekliaus, kai žmogus kelias valandas uždaromas tamsiame, tyliame ir tuščiame kambaryje, rezultatas – tariama nuotaika, transas ar koma.
Vyko elementarus maisto išteklių vystymas, kur nebuvo nei vietos, nei biologinės būtinybės vystytis kompleksiniam elgesiui. Turėdami šio stuburinių evoliucijos laikotarpio pėdsakų, susiduriame su bandymu vystytis sąlyginiai refleksaiįvairiuose šiuolaikinių varliagyvių atstovuose. Itin žemas mokymosi gebėjimas ir ilgalaikės atminties stoka, norint kaupti individualią patirtį, rodo, kad senovės varliagyviai evoliucijos metu niekada nesusidūrė su sudėtingomis elgesio užduotimis.
Jutimo organų vystymosi ypatybės ir sudėtingo roplių elgesio požymiai yra pagrįsti struktūrinės smegenų organizacijos ypatumais. Roplių smegenys skiriasi nuo varliagyvių smegenų kiekiu ir dydžiu kokybiškai. Prieš atsirandant amnionams, pagal dominavimo principą buvo pasirenkamos elgesio strategijos arba atsakas į tam tikrą dirgiklį (žr. III-6 pav., f). Šis principas susideda iš to, kad daugelis pirminių vandens stuburinių ar varliagyvių neturi ryškaus didelio asociacinio smegenų centro (žr. III-6 pav., e). Elgesio forma pasirenkama palyginus maždaug lygiaverčių smegenų dalių, aptarnaujančių įvairius jutimo organus, veiklą. Vieno iš analizatorių smegenų analitinių centrų sužadinimo lygis vaidina lemiamą vaidmenį. Didžiausią sužadinimą smegenyse pasiekęs jutimo organo vaizdavimas tampa pagrindine sprendimų priėmimo sritimi. Pasirinkus vieną iš instinktyvių reakcijų, įvyksta jos elgesio įgyvendinimas. Šis procesas yra kontroliuojamas tuo pačiu paprastas palyginimas dominavimo. Jei reakcijos eigoje atsiranda naujas dirginimas, kuris keičia jutimo organų sužadinimų santykį, tai instinktyvaus proceso elgsenos realizavimas sustoja. Kiekviena konkreti situacija skiriasi nuo ankstesnės, tačiau dalyvauja tie patys pojūčiai. Jei didžiausias sužadinimas pasiekiamas toje pačioje jutimo sistemoje, tada elgesys išsaugomas, o jei kitoje – pasikeičia. Kadangi absoliučiai identiškų sąlygų natūraliame gyvenime praktiškai nebūna, net ir pačių primityviausių anamnijų elgesys bus be galo įvairus. Vadinasi, kiekvieno individo elgesys bus individualus su gana dideliu dinaminiu prisitaikymu.
Pirmieji asociacinio centro požymiai pasirodė varliagyvių smegenyse. Juose tokiu centru galėtų tapti vidurys arba tarpgalvis. Tam buvo visos priežastys. Diencephalone yra neuroendokrininiai centrai, kurie kontroliuoja seksualinį elgesį, migraciją ir anamnioninio organizmo energijos balansą. Suaktyvinus diencephaloninius centrus, paleidžiamos instinktyvios elgesio programos, kurios kontroliuoja kitų smegenų dalių darbą. Atrodytų, kad anamnijos, o vėliau ir vaisiaus vandenų, elgesio analitiniu centru galėtų tapti diencephalonas. Tačiau šiuo atveju elgesio reakcijų įgyvendinimo sistemą sudarytų ne tik nervų sistemos darbas. Kiekvieną kartą bet koks elgesio įvykis paskatintų neuroendokrininių centrų darbą. Hormoninis elgesio reguliavimas įgyvendinamas ilgai, o neurologinis – greitai. Sparčiai keičiantis elgesio formoms, kiltų konfliktas tarp inertiškų hormoninių ir dinaminių neuronų elgesio programų. Vabzdžių atveju šis konfliktas buvo išspręstas neurohormoninių centrų ir grynai instinktyvaus elgesio naudai.
Gana nestabili padėtis susidarė su pirminiais vandens stuburiniais gyvūnais ir varliagyviais. Viena vertus, hormoninio-instinktyvaus varliagyvių elgesio reguliavimo vaidmuo yra labai svarbus ir aiškiai dominuoja pasirenkant elgesio strategijas. Kita vertus, neuromorfologinis substratas yra pakankamai išvystytas, kad įgyvendinant šias strategijas būtų galima šiek tiek individualizuoti elgesį. Atsirado originali hormonų dominuojančio elgesio formų pasirinkimo iš standartinio instinktyvaus rinkinio sistema. Varliagyvių elgesio strategiją lemia neurohormoninė individo būklė. Įgyvendinant pasirinktą elgesio formą, jis prisitaiko prie konkrečių sąlygų, naudodamas dominavimo palyginimą, kuris buvo aprašytas aukščiau. Tokioje anamnio elgesio valdymo schemoje nėra vietos asociatyviniam centrui. Jo gali prireikti tik tada, kai būtų reikalinga greita adaptyvi elgesio individualizacija. Tokia situacija gali susidaryti tik tada, kai nuoseklus instinktyvių elgesio formų realizavimas tiesiogiai priklausys nuo nuolat besikeičiančios situacijos.
Tokiomis nestabiliomis sąlygomis aplinką pasirodė archajiški ropliai. Matyt, išaugo reikalavimai greitam elgesio individualizavimui ir atminčiai, o hormonų dominuojančio principo – rinktis iš standartinio instinktyvaus elgesio rinkinio – įgyvendinimas tapo neefektyvus. Atsirado visiškai naujas sprendimų priėmimo tipas, kuris buvo išsaugotas šiuolaikinių roplių, priklausančių tolimoms sisteminėms grupėms, smegenyse. Visus juos vienija viena iš esmės nauja smegenų savybė – ryškus asociatyvinis centras (žr. III-7 pav.).
Pagrindinis roplių asociacinis centras susiformavo vidurinių smegenų stoge (žr. III-5 pav., c; III-6; III-7, b). Jis atsirado dėl kelių jutimo organų, kurie buvo atstovaujami šioje smegenų dalyje. Pagrindinę vidurinių smegenų stogo dalį užima reprezentacija vizualinė sistema. regos nervai praėję pro chiasmatas, jie kertasi ir pakyla ant vidurinių smegenų stogo. Tinklainės ganglijinių ląstelių aksonai baigiasi ant vidurinių smegenų stogo neuronų, kurie yra suskirstyti į sluoksniuotas struktūras (žr. III-5 pav., c; III-7, b). Yra aiškus topografinis ryšys tarp tam tikros tinklainės srities ir vidurinės smegenų dalies. Tuo pačiu atvaizdo forma ir tarpusavio susitarimas jos elementai. Ilgą laiką priekinė vidurinių smegenų stogo dalis buvo laikoma išskirtinai smegenų centru. vizualinis analizatorius. Tačiau funkciniai-morfologiniai tyrimai parodė, kad taip toli gražu nėra.
Kartu su regos sistemos atvaizdavimu į vidurinių smegenų stogą patenka informacija apie somatinį (odos) jautrumą, motorinį analizatorių, vestibuliarinius ir klausos signalus (žr. III-6 pav., e). klausos analizatorius roplių, jis žymiai padidina savo atstovavimą šiame centre. Dėl to daugeliui roplių, esančių užpakalinėje vidurinės smegenų dalies stogo dalyje, atsiranda nepastebimų porinių išsikišimų - užpakalinių arba apatinių gumbų. Vidurinių smegenų stogas tampa ne vienalyčiu anatominiu dariniu, kaip anamnijoje, o keturkampiu. Jame sutelktas beveik visų pagrindinių nuotolinių ir kontaktinių analizatorių vaizdas. Netgi uoslės sistema yra tarpinių smegenų stoge. Išskyrus uoslės sistemą, beveik visos jutiminės projekcijos į roplių vidurinių smegenų stogą yra topologinės. Tai reiškia, kad informacija iš kiekvienos konkrečios kūno srities pateikiama griežtai tam tikra sritis vidurinių smegenų stogas. Išsaugomas kūno žemėlapio principas, kuris kryptingai perkeliamas į smegenis.
Taigi roplių vidurinių smegenų stoge koncentruojasi įvairi informacija apie jų pačių organizmo ir supančio pasaulio būklę, kuri derinama topologiniu principu. Apsvarstykite, kas nutinka vidurinių smegenų stoge, jei priekinė dešinė roplio galūnė tiesiog stovi ant neįprasto paviršiaus. Vertinant tokią situaciją vidurinių smegenų stoge, lyginamoji analizė somatinė, sensomotorinė, klausos ir vaizdinė informacija. Tai padaryti nesunku, nes visi signalai yra sutelkti viename centre, o dažnai ir vienas ant kito, kaip sluoksniuotame pyrage. Vidurinės smegenys veda sudėtinga analizė daug vieno reiškinio veiksnių, o tai leidžia pasirinkti tinkamiausią atsaką. Tam puikiai tinka stratifikuota vidurinio smegenų stogo struktūra.
Itin supaprastindami realią situaciją, galime teigti, kad vidurinių smegenų stoge vaizduojamos įvairios jutimo sistemos esantys sąlyginėse „grindyse“, išdėstytose horizontalioje plokštumoje. Kiekvieną aukštą užima savotiškas žemėlapis. Tai gali būti momentinis informacinis žemėlapis apie receptorių signalus iš kūno paviršiaus, vaizdas tinklainėje ar akustinis laukas. Visi šie žemėlapiai yra orientuoti savo „grindyse“ taip, kad atspindėtų maždaug tą pačią kryptį erdvėje. Garsinis signalas iš priekinės dešinės galūnės yra po jos vaizdiniu žemėlapiu ir virš somatinio signalo iš pėdos odos. Specializuotos „grindys“ tarpusavyje koreliuojamos naudojant vertikalias nuorodas, kurios leidžia greitai įvertinti konkrečią situaciją ir priimti teisingą sprendimą. Ši vidurinių smegenų stogo schema leidžia suprasti roplių smegenų refleksinį greitį. Matyt, būtent toks greitis tapo pagrindine archajiškų roplių evoliucinės sėkmės priežastimi.
Tobulo refleksinio sprendimų priėmimo centro atsiradimas ropliuose turėjo keletą svarbių pasekmių. Viena vertus, galimybė greitai pasirinkti sprendimus yra beprasmiška, jei bendras medžiagų apykaitos greitis išlieka tame pačiame lygyje. Vadinasi, vidurinių smegenų vystymąsi lydėjo medžiagų apykaitos padidėjimas. Kita vertus, padidėjus vidurinių smegenų stogo dydžiui, atsirado reikalingas ląstelių substratas atminties vystymuisi. Individuali gyvūno patirtis tapo pagrindu lyginti laike atskirtus įvykius. Šį įvykį sunku pervertinti. Pirmą kartą atsirado realūs pagrindai elgesio individualizavimui remiantis įvairių įvykių palyginimu. Reikėtų pažymėti, kad į šiuos neurobiologinius roplių smegenų pranašumus paprastai net neatsižvelgiama rekonstruojant ankstyvą amniono evoliuciją (Carroll, 1982).
Archajiški ropliai, vystydami refleksologinės analizės centrą, gavo materialų substratą įvairiems įvykiams prisiminti. Jiems tapo prieinamas individualios patirties atkūrimas, kuris yra mokymosi pagrindas. Varliagyviai nebegalėjo konkuruoti su savarankiškai besimokančiais archajiškais ropliais. Hormoniniai-instinktyvūs varliagyvių, žuvų ir bestuburių elgesio principai pavertė juos maistu ropliams, turintiems išsivysčiusias refleksines-asociacines vidurines smegenis.
Visi šie roplių smegenų struktūros pranašumai negalėjo atsirasti savaime. Tokiam giliam kokybiniam smegenų pertvarkymui turėjo susiklostyti itin atšiaurios ir nepaprastos sąlygos. Archajiški ropliai turėjo atsidurti savotiškoje aplinkoje, kuriai keliami labai aukšti smegenų ir individualios atminties analitinių savybių reikalavimai.
Iš knygos Aukštutinės Juros periodo roplių liekanų radiniai Saveljevskio skalūnų kasykloje autorius Iš knygos Aukštutinės Juros periodo roplių liekanų radiniai Saveljevskio skalūnų kasykloje autorius Žuravlevas Konstantinas IvanovičiusRoplių liekanų radiniai 1931 m., pačiais pirmaisiais skalūnų kasybos mėnesiais, kai kasyba dar buvo vykdoma atvirose duobėse, buvo gauti keli ichtiozaurų liekanų radiniai. Tarp šių radinių yra 13 uodegos slankstelių, rastų naftingųjų skalūnų sluoksnyje prie pagrindo.
autorius§ 36. Bendrasis roplių nervų sistemos sandaros planas Plačiai vystantis žemei, pasikeitė roplių smegenys. Tačiau jo morfologiniai pertvarkymai iš pirmo žvilgsnio nebuvo tokie esminiai kaip varliagyvių. Paviršinė anatominė analizė rodo, kad senovės
Iš knygos Smegenų kilmė autorius Saveljevas Sergejus Viačeslavovičius§ 38. Roplių smegenų atsiradimo sąlygos Asociatyvus smegenų centras negali atsirasti atsitiktinai. Smegenų asociacinio centro išlaikymo energijos sąnaudos ir elgesio strategijų keitimo išlaidos visada yra labai didelės (žr. I skyrių). Tam turi būti priežastis
Iš knygos Smegenų kilmė autorius Saveljevas Sergejus Viačeslavovičius§ 40. Adaptyvusis archajiškų roplių spinduliavimas Smulkūs plėšrūs ir visaėdžiai ropliai, išsivystę agresyvioje augalų liekanų aplinkoje, nebuvo didžiausi to meto gyvūnai. Karbono pabaigoje jų kūno dydis paprastai neviršydavo kelių dešimčių
Iš knygos Biologinė chemija autorius Lelevičius Vladimiras ValerjanovičiusFermento aktyvi vieta Fermento molekulės sritis, kuri specifiškai sąveikauja su substratu, vadinama aktyvia vieta. Aktyvioji vieta yra unikalus aminorūgščių likučių derinys fermento molekulėje, kuris užtikrina tiesioginį
autoriusKur yra Žemės ir Mėnulio sistemos masės centras? Žemės ir Mėnulio sistemos masės centras, vadinamasis baricentras, yra 4672 kilometrų atstumu nuo Žemės centro link Mėnulio, tai yra, maždaug 1700 kilometrų gylyje žemiau Žemės paviršiaus. Griežtai kalbant, pagal
Iš knygos naujausia knyga faktus. 1 tomas. Astronomija ir astrofizika. Geografija ir kiti žemės mokslai. Biologija ir medicina autorius Kondrašovas Anatolijus PavlovičiusKas yra malonumo centras ir kur jis yra kūne? Viena iš smegenų dalių yra pagumburis, kuris yra diencephalono dalis ir yra po regos gumburais (talamu). Pagumburis, kuriame yra autonominiai centrai
autorius Kondrašovas Anatolijus Pavlovičius Iš knygos „Naujausia faktų knyga“. 1 tomas [Astronomija ir astrofizika. Geografija ir kiti žemės mokslai. Biologija ir medicina] autorius Kondrašovas Anatolijus Pavlovičius Iš knygos Farmacijos ir maisto mafija pateikė Brower LouisFarmakologinės priežiūros centras „Paryžius, 1977 m. vasario 4 d. NCPC, įkurtas 1974 m. sausio mėn., vasario 2 d. Generalinėje asamblėjoje susirinko kartu su Nacionalinių gydytojų ir farmacininkų chartijų atstovais, prie kurių prisijungė grupė kontrolės centrų. apsvaigimo ir
Iš knygos Laiko laukuose autorius Čiževskis vokietis MichailovičiusVarliagyvių, roplių, paukščių ir žinduolių pagrindinių grupių klasifikacija ir trumpas aprašymas VARLIŲ ARBA VARLIŲ KLASĖ.
autorius Kandel Erikas Ričardas Iš knygos „Atminties beieškant“ [Naujo žmogaus psichikos mokslo atsiradimas] autorius Kandel Erikas Ričardas Iš knygos Psichopatai. Patikima istorija apie žmones be gailesčio, be sąžinės, be sąžinės graužaties pateikė Keel Kent A.Mendotos nepilnamečių reabilitacijos centras Dešimtojo dešimtmečio pradžioje Jungtines Valstijas užklupo tikra paauglių smurto epidemija. Nuo 1980 iki 1993 metų nepilnamečių nusikaltimų skaičius išaugo beveik dvigubai. Atrodė, kad niekas negali to sustabdyti
Iš knygos Skaitymas tarp DNR eilučių [Antrasis mūsų gyvenimo kodas, arba knyga, kurią turi perskaityti kiekvienas] autorius Shpork PeterNuo Berlyno iki būsimos revoliucijos centro Jis neįsižeidžia, kai jį painioja su studentu ar doktorantu. Ir tai 32 metų genetikui nutinka nuolat. Aleksandras Meisneris - pilkai mėlynos akys, tamsiai šviesūs nerūpestingai sušukuoti plaukai, trijų dienų ražiena - ne tik jauni,
Medžiagos pavadinimas: Zoologijos ir zoogeografijos pagrindai
bendrosios charakteristikos
Dėl roplių kaklo atsiradimo galva yra judriai sujungta su kūnu ir gali judėti ne tik, kaip ir varliagyvių, vertikalia, bet ir horizontalia kryptimi.
Kūnas pailgas (tik vėžlių platus), gana lankstus; daugumos rūšių uodega yra ilga, galinti įvairiai judėti; kojos stipriai išsivysčiusios (kai kurioms driežų rūšims ir visoms gyvatėms kojos sumažėjusios dėl perėjimo prie kito aktyvaus judėjimo būdo). Vidutinis kūno dydis yra daug didesnis nei varliagyvių, o kai kurios rūšys pasiekia milžiniškus dydžius.
Viršeliai. Odą dengia ragiškai stiprūs ir kartu lengvi ektoderminės kilmės žvyneliai, kurie saugo organizmą ne tik nuo mechaniniai pažeidimai bet ir nuo išdžiūvimo. Lydymosi metu keratinizuotas odos sluoksnis periodiškai išsilieja. Vėžliams ir krokodilams osifikacijos vystosi mezenchiminiame odos sluoksnyje, o kitiems ropliams jų nėra. Vėžlius nuo priešų gerai apsaugo kriauklės. Šiuolaikinių roplių oda, skirtingai nei varliagyvių, yra sausa, be liaukų.
Išorinių gaubtų spalva yra įvairi ir daugeliui rūšių turi apsauginę vertę, nes jos dėka gyvūnai yra sunkiai pastebimi aplinkos fone.
Nervų sistema. Dėl judresnio ir sudėtingesnio gyvenimo būdo roplių nervų sistema yra geriau išvystyta nei varliagyvių (95 pav.).
Ryžiai. 95. Driežo smegenys, vaizdas iš šono:
1 - pusrutulis priekinės smegenys; 2 - uoslės skiltis; 3 - diencephalonas; 4 - hipofizė; 5 - vidurinės smegenys; 6 - optinis chiasmas (chiasmas); 7 - smegenėlės; 8 - nugaros smegenys
Priekinės smegenys yra žymiai dideli dydžiai nei pastarieji (ypač dėl striatum padidėjimo), atsiranda pusrutulių paviršiuje nervų ląstelės- smegenų žievės užuomazga. Todėl ropliai greitai užmezga naujus ryšius su išorine aplinka nei jų protėviai. Didėja šios smegenų dalies, kaip viso organizmo gyvybinės veiklos koordinatoriaus, vaidmuo. Sudėtingėja tarpinių ir vidurinių smegenų struktūra, sustiprėja jų ryšys su priekinėmis smegenimis ir kitomis centrinės nervų sistemos dalimis. Smegenėlės dėl judėjimo metodų komplikacijos ir būtinybės išlaikyti kūno pusiausvyrą yra daug geriau išsivysčiusios nei varliagyvių. Padidėjo ir pailgosios smegenys, kurios sudaro ropliams būdingą vertikalų vingį. Nugaros smegenys yra ilgos; jos struktūra tapo sudėtingesnė. Tai buvo išreikšta ryškesniu išorinio sluoksnio (balto, ty takų) atskyrimu nuo vidinio (pilko, ty nervinių ląstelių) sluoksnio.
Iš roplių jutimo organų (išskyrus gyvates ir kai kurias kitas rūšis) akys yra geriau išsivysčiusios nei varliagyvių: jos judresnės. Jų prisitaikymas prie regėjimo įvairiais atstumais (daugelyje rūšių gana didelis) užtikrinamas ne tik, kaip žuvims ir varliagyviams, lęšiuko judėjimas, bet ir, kaip ir aukštesniųjų stuburinių, keičiant jo kreivumą. Be akių vokų, yra ir žadinanti membrana. Nemažai rūšių turi parietalinį organą, susietą su diencephalonu ir galintį suvokti šviesos dirgiklius. Kai kurie progresuojantys klausos organo pokyčiai: šiek tiek padidėjo apatinis maišelis; klausos kaulas geriau perduoda vibracijas ausies būgnelisį vidinę ausį; be ovalo lango atsirado antras langas - apvalus, kuris padidina labirintą supančios egzolimfos mobilumą; Būgninė membrana yra ne galvos paviršiuje, o nedidelėje įduboje. Kvapo organai yra gana gerai išsivystę, jų vidinis paviršius padidėjo. Daugumos rūšių skonio organai yra prastai išvystyti. Kalba bendraujant vaidina svarbų vaidmenį.
Skeletas. Jis beveik visiškai sukaulėjęs dėl stambiųjų pluoštų pakeitimo kaulinis audinys, būdingas varliagyviams, plonas pluoštinis, padidėjo jo stiprumas, pagerėjo atraminės ir apsauginės funkcijos (96 pav.).
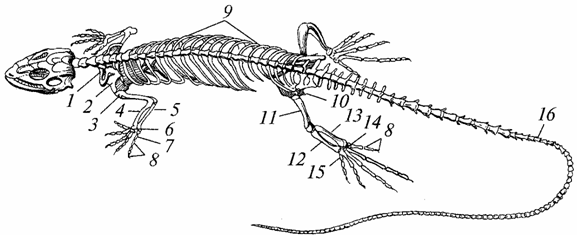
Ryžiai. 96. Driežo skeletas:
7 - raktikaulis; 2 - mentė; 3 - petys; 4 - spindulys; 5 - alkūnkaulis; b - riešas; 7 - metakarpas; 8 - pirštų falangos; 9 - šonkauliai; 10 - dubuo; 11 - šlaunys; 12 - didelis blauzdikaulis; 13 - šeivikaulis; 14 - tarsus; 15 - padikaulis; 16 - uodegos slanksteliai
Padidėjusi kaukolė; smegenų skyriaus pajėgumas tapo didesnis nei varliagyvių. Šoninis paviršius senovės roplių kaukolė buvo vientisa, vėliau joje atsirado laikinų duobių - viena ar dvi, kurios sumažino jos masę ir padidino tvirtinimo paviršių kramtymo raumenys. Apatiniai žandikauliai, kaip ir varliagyvių, yra pakabinti nuo kaukolės smegenų srities kvadratinio kaulo pagalba, taip pat susideda iš kelių kaulų (dantų, sąnarių ir kt.).
Pirmojo slankstelio kūnas (atlasas) išaugo danties pavidalu iki antrojo slankstelio kūno (epistrofija). Žiedo formos atlasas gavo galimybę suktis aplink epistrofinį dantį, dėl kurio tapo įmanoma pasukti galvą, ir tai didelę reikšmę maisto (daugiausia kitų gyvūnų) gavimui ir apsaugai nuo priešų. Atlasas yra sujungtas su kaukolės pakaušio sritimi vienos kondilo pagalba, o tai taip pat prisideda prie galvos mobilumo padidėjimo. Stuburas yra padalintas į penkias dalis: gimdos kaklelio, krūtinės, juosmens, kryžmens ir uodegos. Slanksteliai krūtinės ląstos yra sujungti su krūtinkauliu gerai išvystytais šonkauliais, dėl kurių susidaro krūtinė, kurios varliagyviams nėra. Sutrumpinti šonkauliai tęsiasi nuo juosmens slankstelių, nesiekia krūtinkaulio. Yra du kryžmens slanksteliai (varliagyviai turi vieną). Uodegos srityje yra daug slankstelių (išskyrus vėžlius).
Kojų kaulai yra geriau išsivystę ir patvaresni nei varliagyvių. Tarp riešo ir liemens kaulų yra judantys sąnariai. Todėl dilbis ir proksimalinė riešo eilė, taip pat blauzda ir proksimalinė riešo eilė veikia kaip vientisi dariniai. Ypač atkreiptinas dėmesys į galūnių diržų stiprinimą: pečių – dėl jungčių su krūtinkauliu ir krūtinės ląstos stuburu, dubens – dėl to, kad remiasi dviem kryžkaulio slanksteliai ir abi jo puselės sudaro dvi sąaugas – sėdmeninę ir gaktos.
Raumenų sistema. Jis buvo diferencijuojamas, susiformavo naujos raumenų grupės, ant kurių sukasi galva, kaklo judesiai, atsiradę ropliams, išsiplėtimas ir susiaurėjimas. krūtinė ir kt.. Žymiai padidėjo kojų raumenys, išsivystė pirštų lenkiamosios ir tiesiamieji raumenys. Padidėjo visos sistemos masė ir stiprumas.
Virškinimo sistema. Jis labiau diferencijuotas nei varliagyvių (97 pav.). Dantys daugiausia sutelkti į žandikaulius, jų skaičius yra didelis, tačiau jie yra to paties tipo ir daugiausia skirti grobiui sugauti. Seilių liaukos yra geriau išsivysčiusios nei varliagyvių (kai kuriose rūšyse šių liaukų išskyros yra nuodingos). Ropliuose prasideda kietojo gomurio formavimosi procesas, kuris ypač išvystytas krokodilams. Dėl kietojo gomurio susidarymo burnos ertmė yra atskirta nuo nosiaryklės ertmės, o tai palengvina tiek kvėpavimą, tiek maisto įsisavinimą. Ryklė, stemplė ir skrandis yra gerai išvystyti (ypač plėšrūnų, kurie minta stuburiniais gyvūnais). Kepenys (su tulžies pūsle) ir kasa yra tobulesnės nei varliagyvių. Plonoji žarna gerokai pailgėjusi, storoji žarna trumpa ir baigiasi kloaka. Šiuolaikiniai ropliai daugiausia minta gyvūnais, žolėdžių rūšių skaičius yra nereikšmingas.
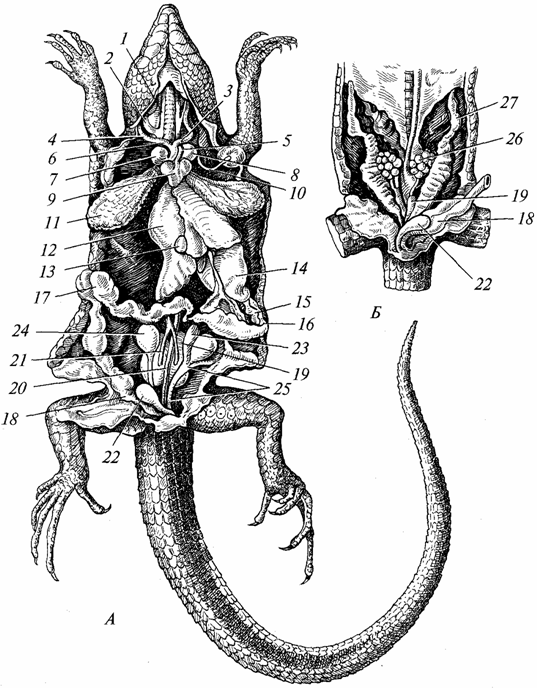
Ryžiai. 97. Vidaus organai driežai; A - vyras; B – moteris:
/ - išorinis ir 2 - vidinis jugulinės venos; 3 - kairioji ir 4 - dešinė miego arterijos; 5 - kairė ir 6 - dešinė aortos lankai; 7 - dešinysis ir 8 - kairysis atriumas; 9 - širdies skilvelis; 10 - kairioji poraktinė arterija; 11 - plaučiai; 12 - kepenys; 13 - tulžies pūslė; 14 - skrandis; /5 - kasa; 16 - dvylikapirštės žarnos; 17 - storoji žarna; 18 - tiesioji žarna; 19 - inkstai; 20 - nugaros aorta; 21 - eferentinės inkstų venos; 22 - šlapimo pūslė; 23 - sėklidės priedas; 24 - sėklidė; 25 - sėklų vamzdelis; 26 - kiaušidės; 27 - kiaušintakis
Kvėpavimo sistema. Roplių kvėpavimo organai yra plaučiai (žr. 97 pav.), nes jų oda netinkama dujų mainams; gleivinė burnos ertmė, kuris atlieka reikšmingą vaidmenį varliagyvių kvėpavime, šiai funkcijai atlikti naudoja kelios roplių rūšys. Dėl šios priežasties ir ypač dėl padidėjusio roplių aktyvumo, palyginti su varliagyviais, plaučių struktūra ir kvėpavimo takų sudėtingas. Plaučiai yra didesni organai ir yra giliau krūtinės ertmėje nei varliagyvių. Jų vidinis paviršius žymiai padidėjo, nes ant jo buvo sukurta daug skersinių su tankesniu tinklu. kraujagyslės. Oras patenka į plaučius, eidamas per choanus, ryklę, gerklas, ilgą trachėją, kurios spindį palaiko kremzliniai žiedai. Praeinant per trachėją, oras išvalomas nuo dulkių, jo temperatūra artėja prie kūno temperatūros. Varliagyviams trachėja buvo tik užuomazgoje. Užpakalinis trachėjos galas yra padalintas į du bronchus, kurių sienelėse taip pat yra kremzlinių žiedų. Įkvėpimas ir iškvėpimas yra efektyvesnis nei varliagyvių, nes roplių krūtinė išsivysto, jos tūris didėja įkvėpus ir mažėja iškvėpus. Toks pat mechanizmas buvo išsaugotas ir aukštesniems stuburiniams gyvūnams – paukščiams ir žinduoliams.
Kraujotakos sistema. Širdis, kaip ir varliagyvių, yra trijų kamerų, tačiau iš apačios skilvelyje atsiranda pertvaros gemalas. Krokodilams jis jau pilnas, jų širdis tampa keturių kamerų (du prieširdžiai ir du skilveliai). Vadinasi, pas roplius sustiprėja veninio ir arterinio kraujo srovių atskyrimas, tačiau visiškas abiejų srovių izoliavimas dar nėra baigtas net krokodiluose, kaip matyti iš toliau pateikto aprašymo (98 pav.).
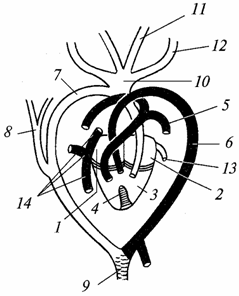
Ryžiai. 98. Driežo širdis ir pagrindinės kraujagyslės (šviesos - kraujagyslės su arteriniu krauju, šešėliai - su mišriu krauju; juodos - su veninėmis):
1 - dešiniojo prieširdžio; 2 - kairysis atriumas; 3 - skilvelis; 4 - nepilna pertvara, padalijanti skilvelį į kairę ir dešinę puses; 5 - plaučių arterija; 6 - kairioji aortos lanka; 7 - dešinysis aortos lankas; 8 - poraktinė arterija; 9 - nugaros aorta; 10 - bendras kamienas miego arterijos; 11 - kairioji vidinė miego arterija, 12 - kairioji išorinė miego arterija; 13 - plaučių vena; 14 - atgal tuščiosios venos ir priekinės tuščiosios venos (jugulinės) venos
Arterinis kraujas patenka į kairįjį prieširdį – jis ateina iš plaučių. Veninis kraujas patenka į dešinįjį prieširdį – jis surenkamas iš visų kūno dalių. Toliau kraujas iš abiejų prieširdžių patenka į skilvelį: dešinėje jo pusėje - veninis, kairėje - arterinis. Dėl pertvaros, nors ir nepilnos, kraujas skilvelyje susimaišo mažiau nei varliagyvių. Arterijos kūgio nėra, o pagrindiniai arterijų kamienai nukrypsta tiesiai iš širdies.
Nuo dešinioji pusė skilveliai išsiskiria plaučių arterijos(buvusi ketvirtoji šakotųjų arterijų pora), pernešanti veninį kraują į plaučius. Buvusi trečioji šakotųjų kraujagyslių pora atrofavosi. Antroji kraujagyslių pora susideda iš dviejų aortos lankų: dešinysis lankas prasideda nuo kairiosios skilvelio pusės ir teka arterinis kraujas, kairysis lankas prasideda nuo dešinės skilvelio pusės (bet arčiau kairiosios pusės) ir teka mišrus kraujas. . Nuo dešiniojo aortos lanko miego arterijos (buvusi pirmoji žiauninių kraujagyslių pora) tęsiasi iki galvos. Jie aprūpina smegenis, ypač jautrias deguonies trūkumui, arteriniu krauju. Abu aortos lankai susijungia ir sudaro aortą, kuri išsišakoja į daugybę arterijų. Taigi roplių kūnas daugiausia aprūpinamas mišriu krauju, tačiau jis yra prisotintas daugiau deguonies nei varliagyvių, o tai prisideda prie intensyvesnės medžiagų apykaitos.
išskyrimo sistema. Tai labai pasikeitė dėl padidėjusio roplių medžiagų apykaitos greičio, dėl kurio padaugėjo disimiliacijos produktų, ir jų visiško prisitaikymo prie antžeminio gyvenimo būdo, dėl ko kilo būtinybė ekonomiškiau naudoti patenkantį vandenį. kūnas. Todėl ropliai sukūrė sudėtingesnius šalinimo organus, vadinamus dubens arba antriniais inkstais (metayaephros). Visi šių inkstų kanalėliai neturi piltuvėlių ir prasideda Bowmano kapsulėmis, supančiomis Malpighian kūnus su padidėjusiu kapiliarų skaičiumi. Dubens inkstų kanalėliai yra labai ilgi ir tankiai susipynę su kapiliarais. Bowmano kapsulėse iš disimiliacijos produktų, ištirpusių vandenyje ir kai organinės medžiagos(pvz., gliukozės) susidaro pirminis šlapimas. Pastarieji, eidami per ilgus inkstų kanalėlius, per jų sieneles pinančius kapiliarus grąžina kraujui nemažą dalį vandens ir kitų organizmui reikalingų medžiagų. Po to susidaręs antrinis šlapimas išeina ne vilko kanalais, o naujai išsivysčiusiais šlapimtakiais į kloaką ir kaupiasi šlapimo pūslė. Pirminio ir antrinio šlapimo susidarymo procesai taip pat atliekami kamieniniuose inkstuose, tačiau į kraują grįžta mažesnis vandens kiekis. Pirminiai vandens stuburiniai gyvūnai, ypač varliagyviai, kurių oda plika, dažnai kenčia nuo vandens pertekliaus, o ne dėl jo trūkumo. Taip pat reikėtų pažymėti, kad roplių galutinis baltymų skilimo produktas yra šlapimo rūgštis, kurio pašalinimui iš organizmo su šlapimu išleidžiama mažai vandens. Varliagyviams ir daugeliui kitų pirminių vandens stuburinių gyvūnų skaidant baltymus susidaro karbamidas, kuriam pašalinti reikia daug vandens.
Roplių embrionai turi galvos inksto užuomazgas ir išsivysčiusį kamieninį inkstą, kuris veikia suaugusiems (kai kuriose driežų rūšyse) iki brendimo. Šie faktai patvirtina trijų tipų inkstų tęstinumą stuburinių gyvūnų evoliucijos metu.
Reprodukcija. Roplių patinų reprodukcinė sistema apima dvi apvalios formos sėklides, gulinčias juosmeninės stuburo dalies šonuose, du sėklidžių priedus, suformuotus iš priekinių kamieno inkstų dalių, du sėklinius latakus (Volfo kanalus) ir iš sienelės išsivysčiusius kopuliacinius organus. kloakos. Moteriškas dauginimosi sistema susideda iš dviejų kiaušidžių, kurios stipriai auga veisimosi sezono metu, ir dviejų kiaušintakių (Miulerio kanalų), kurie įteka į kloaką. Tręšimas yra tik vidinis. Sėklą į patelės kloaką įneša patinai, pasitelkę kopuliacinius organus. Subrendę kiaušinėliai, kuriuose yra daug trynio, patenka į kiaušintakių piltuvus. Moteriškos ir vyriškos lytinės ląstelės susijungia viršutinėje kiaušintakių dalyje, kur po lytinių santykių prasiskverbia spermatozoidai. Zigotos, judančios išilgai kiaušintakių, pastarųjų vidurinėje dalyje yra apsuptos baltyminiu apvalkalu, o nugaroje – apvalkalu.
Kai kuriose driežų ir gyvačių rūšyse stebimas partenogenetinis patelių dauginimasis. Šiose rūšyse patinų paprastai nėra.
Plėtra. Roplių vystymasis yra tiesioginis; ropliai neturi lervų. Apvaisintus kiaušinėlius ropliai dažniausiai deda į dirvą, o kai kurias rūšis – po pūvančiomis augalų liekanomis. Daugelyje rūšių zigotų vystymasis vyksta patelės lyties organuose, iš karto po išsivysčiusių kiaušinėlių išleidimo iš jų išsirita susiformavę gyvūnai, t.y. tokioms rūšims būdingas ovovivipariškumas. Kai kuriose rūšyse stebimas tikras gyvas gimimas: trynio maišelio kraujagyslės yra glaudžiai susijusios su kiaušintakio kraujagyslėmis, o embrionas gauna didelę dalį maistinių medžiagų iš motinos kūno. Kiaušialąstė dažniau stebima rūšyse, paplitusiose šiauriniuose ir kalnuotuose regionuose. Akivaizdu, kad ovoviviparity yra besivystančių embrionų apsauga nuo aušinimo ir kitų nepalankių sąlygų. Nėra nė vienos roplių rūšies, kurios kiaušinėliai vystosi vandenyje. Netgi diriguojant ropliams dauguma gyvenimas vandenyje (visi krokodilai, daug vėžlių, kai kurios gyvatės), kiaušialąstės ir vystymasis vyksta tik sausumoje. Jūros gyvatės, kurios niekada nepalieka vandens, yra gyvybingos.
Roplių kiaušiniai ir embrionai turi daugybę pritaikymų vystymuisi sausumoje. Pirma, kiaušiniai padengiami lukštais ir kitomis membranomis, kurios apsaugo juos nuo išdžiūvimo ir pažeidimų. Antra, embrionuose susidaro specialios embrioninės membranos (serozinės, amniono ir alantozės), kurios yra labai svarbios užtikrinant. embriono vystymasis gyvūnai sausoje žemėje. Serosa ir amnionas išsivysto iš tos pačios dvigubos embriono kūno raukšlės, kilusios iš ektodermos ir mezodermos. Ši raukšlė atsiranda iš dviejų priešingos pusės užuomazgų pavidalu, kurie auga vienas kito link ir užsidaro virš embriono. Išorinė raukšlės dalis virsta serozine (jos išorinis sluoksnis – ektoderminis, vidinis – mezoderminis), o vidinis – amnionu (jos išorinis sluoksnis – mezoderminis, vidinis – ektoderminis). Amniono suformuota ertmė užpildoma vandeniniu skysčiu, dėl kurio vystymasis vyksta drėgnoje aplinkoje, o embrionas apsaugotas nuo išdžiūvimo ir įvairaus drebėjimo, ypač pavojingo ore.
Kilmė. Pirmieji ropliai, atsiradę antroje karbono periodo pusėje, gyveno drėgnuose žemės biotopuose ir tikriausiai veisėsi bei vystėsi vandenyje. Tuo metu žemėje jau gyveno žemesni sausumos augalai, įvairūs bestuburiai, orą pradėjo užkariauti vabzdžiai. Ropliai, kurie iš vandens telkinių galėjo nukeliauti toliau nei varliagyviai, turėjo gausų maisto ir nesusidūrė su rimtais priešais. Karbono pabaigoje ir ne kartą per Permo klimatas tapo sausesnis, o tai sukėlė daugelio varliagyvių mirtį ir, atvirkščiai, prisidėjo prie laipsniško roplių vystymosi ir platesnio pasiskirstymo sausumoje. Po kurio laiko jie perėjo prie dauginimosi ir vystymosi už vandens telkinių ribų, pirmiausia drėgnesniuose sausumos biotopuose, o vėliau – sausesnėse vietose. Mezozojaus eroje jie išplito visame pasaulyje ir daugelyje įvairios grupės. Kartu su plėšrūnais „atsirado daug žolėdžių rūšių, atsirado skraidyti prisitaikę ropliai, jūrose gyvenantys ropliai. Jų kūno dydis buvo skirtingas, tačiau dauguma buvo stambūs gyvūnai, o kai kurie buvo milžiniški – iki 20 m ir daugiau ilgio. Klimatas beveik per visą mezozojaus epochą buvo šilta, labai palanki šiems gyvūnams, kurie neturi pakankamai aukšto lygio pastovi temperatūra kūno ir praranda savo aktyvumą, kai aplinka vėsta.
Mezozojaus eros pabaigoje, pasaulis vyko galingi kalnų kūrimo procesai, kuriuos lydėjo staigus egzistavimo sąlygų pasikeitimas (temperatūra, drėgmė ir kt.). Tai paskatino masinį mezozojaus roplių išnykimą, kurie negalėjo pakęsti kovos už egzistavimą tokiomis jiems sudėtingomis abiotinėmis aplinkos sąlygomis, kurias apsunkino konkurencija su žinduoliais ir paukščiais. Tuo pat metu kainozojaus epochoje atsirado ir daugėjo naujų roplių grupių – iš jų kilo driežai ir gyvatės. Iš senųjų grupių išliko tik viena labai primityvi pirminių driežų rūšis – nedidelis skaičius krokodilų ir vėžlių. Šiuolaikiniai ropliai daugiausia paplitę šiltuose kraštuose, ypač subtropinėse ir atogrąžų šalyse, vidutinio klimato šalyse jų nedaug, o tolimojoje šiaurėje jų nėra.

