Mišićno-koštani sustav služi za držanje tijela u određenom položaju i kretanje u prostoru. Mišićno-koštani sustav sastoji se od koštanog sustava (skeleta), ligamenata, zglobova i skeletni mišić. Kosti, ligamenti i zglobovi pasivni su elementi organa za kretanje. Mišići su aktivni dio motoričkog sustava.
Evolucija somatomotornog živčanog sustava
U nastavku ćemo se baviti samo somatomotornim živčanim sustavom koji inervira skeletne mišiće. Animacija: molekularni mehanizam mišićne kontrakcije. Somatomotorni živčani sustav se mijenja, postupno se razvija tijekom evolucije. Razvoj se odvija paralelno s formiranjem skeletnih mišića i mehanizma kretanja, budući da strukture ovise jedna o drugoj. Tijekom evolucije o životinjama možemo govoriti samo ako funkcioniraju mišića pojavljuje u ljudskom tijelu. Primitivne, jednostavno konstruirane životinje s umreženim živčanim sustavom reagiraju na udar okoliš uz snažno kretanje njihove cijele mišićna masa, čija je svrha bila izbjeći štetnih utjecaja ili pristupiti njihovoj hrani.
Kostur služi kao oslonac i zaštita cijelom tijelu i pojedinačna tijela, a mnoge kosti su i moćne poluge, uz pomoć kojih se izvode različiti pokreti tijela i njegovih dijelova u prostoru. Mišići pokreću cijeli sustav koštanih poluga. Kostur čini osnovu tijela i određuje njegovu veličinu i oblik. dijelovi kostura kao što je lubanja, prsni koš i zdjelice kičmeni stup, služe kao mjesto skladištenja i zaštite vitalnih organa - mozga, pluća, srca, crijeva itd. Kostur je uključen u metabolizam, posebno u održavanju na određenoj razini mineralni sastav krv. Osim toga, brojne tvari koje čine kosti (kalcij, fosfor, limunska kiselina itd.), Ako je potrebno, lako ulaze u metaboličke procese. Organsku osnovu koštane tvari čine uglavnom bjelančevine, dok mineralnu čine soli kalcija i fosfora.
Njihov jednostavan živčani sustav i mišićni sustav više nije bilo dopušteno diferencirano kretanje. Tijekom evolucije razvijali su se sve savršeniji pokreti. One su postale moguće zahvaljujući razvoju, diferencijaciji živčani sustav i mehanizam kretanja te njihovo usklađivanje.
Mekušci koji žive bez fiksnog kostura prolaze kroz valovite peristaltičke kontrakcije svojih mišićnih prstenova. Segmente mišićnih prstenova kontroliraju ganglijske stanice koje su uzdužno povezane s živčana vlakna za sinkronizaciju pokreta.
Kada se povećaju, tvrde i glatke površine kostiju imaju poroznu strukturu. postojati različiti tipovi koštano tkivo, koji su najčešće različite dijelove jedna kost: kompaktni sloj i spužvasta tvar. Kosti kao što su kralješci, vrat bedrene kosti, epifiza radius, sastoje se uglavnom od spužvaste tvari. U spužvastoj tvari koštane su grede raspoređene u obliku zakrivljenih ploča povezanih poprečnim ili kosim prečkama. Duge cjevaste kosti udova sastoje se uglavnom od tvari u kojoj su koštane ploče smještene vrlo tijesno.
U početku difuzno lokaliziran mišićna vlakna kasnije formiraju međusobno povezane mišićne snopove; mišići. Evolucija fiksnog kostura omogućila je brzi razvoj, što je različito riješeno kod člankonožaca kao vanjskog kostura, kod kralježnjaka - kao unutarnjeg kostura. Dok su oba učinkovita u pomaganju kretanja tijela, potonje se rješenje pokazalo korisnijim jer je omogućilo neometan rast, omogućujući stvaranje većih, složenijih živih bića.
Živčani sustav člankonožaca je segmentiran. Aktivnost udova u medijalnom segmentu kontroliraju segmentni gangliji. Gangliji artropoda mogu raditi neovisno. Iako je ganglion u glavi veći od drugoga, on ne kontrolira aktivnost ostalih ganglija. Njegova veličina prilično opskrbljuje oči, antene i alate za usta koji su pohranjeni na glavi. Kad se artropodu odsiječe glava, životinja se također može kretati nekoliko dana dok ne umre od gladi bez usta. Interganglijske veze osiguravaju koordinaciju pokreta ekstremiteta.
Kosti kao i drugi unutarnji organi sastoje se od stanica. Postoje posebne stanice koje stalno uništavaju koštanu tvar (osteoklasti); stanice koje obnavljaju, obnavljaju kost (osteoblasti), te stanice odgovorne za stvaranje koštanog skeleta i mineralizaciju koštanog tkiva (osteociti).
Tijekom ljudskog života u kostima se neprestano odvijaju procesi restrukturiranja koštanog tkiva: u nekom odvojenom malom području koštanog tkiva, koštano tkivo se uništava, zatim se uklonjena stara kost zamjenjuje točno istom količinom nove kosti. . Kod zdravih ljudi procesi razaranja koštanog tkiva i stvaranja nove kosti kvantitativno su isti. Aktivnost ovih stanica kontroliraju mnoge biološki aktivne tvari, kao što su hormoni štitnjače i paratireoidnih žlijezda, hormoni kore nadbubrežne žlijezde, vitamin D3 i, konačno, spolni hormoni (estrogeni i progesteron). Rast i razvoj koštanog tkiva događa se do 16-25 godina. Nakon postizanja maksimalne razine koštane mase, u dobi od 30-40 godina, počinje njezin neznatan gubitak, koji iznosi 0,2-0,5% godišnje.
U usporedbi s njegovim složena funkcija, iznenađujuće malo živčanih stanica nalazi se u gangliju. Cijeli mišić uda obično je opskrbljen s četiri neurona. Veze između neurona osiguravaju uzajamnu inervaciju između fleksora i ekstenzora te za koordinaciju pokreta s drugim udovima.
Prvi kralježnjaci su životinje koje žive u vodi; Riba. U početku su se njihovi skeletni mišići sastojali samo od aksijalnih mišića, koje je apsorbirala vertebralna kiselina i koji su bili izvedeni iz miotoma. Mišići omogućuju samo savijanje i okretanje tijela. Prva homogena aksijalna mišićna masa kasnije se segmentira; tako su se razvili pojedini mišići. Tada su mišići bili rastegnuti iu zidu tijela. Kao sljedeći rezultat evolucije, peraje, a zatim, nakon što su se neki odvažili na kopno, raspoređeni udovi.
U dobi od 30-40 godina gubitak koštane mase je 0,5% godišnje. A nakon početka menopauze kod žena se godišnje gubi 3-5% koštane mase.
U ljudskom tijelu je također izoliran meki kostur (kostur) koji sudjeluje u držanju organa u blizini kostiju. Meki kostur uključuje fascije, ligamente, vezivno tkivne kapsule organa i druge strukture. Većina mišića je pričvršćena za kosti. Mišići uključuju kosti kostura u pokretu i obavljaju posao. Mnogi mišići koji okružuju tjelesne šupljine štite unutarnje organe.
Mišići koji stvaraju te strukture izbijaju iz stijenke tijela. Razvoj udova bio je skokovit korak u evoluciji sustava za kretanje kralježnjaka. Udovi omogućuju učinkovitije kretanje živih bića, pa su se njihovi životni uvjeti, njihova prilagodba okolini znatno poboljšali. Tijekom evolucije njihova struktura i veličina stalno su se poboljšavale. Udovi koji su znatno veći od veličine tijela omogućavali su dvonožni život. Kao rezultat toga, mogli su bolje navigirati s višim osjetilnim organima u glavi, a njihove ruke, oslobođene tereta kretanja, mogle su se koristiti u druge svrhe.
Stanje kostiju ovisi o teretu koji nose. Dobro razvijeno mišićno tkivo jača zglobove i doprinosi normalnom razvoju i funkciji kostiju. I mišići i kosti gube na masi ako je opterećenje na njih premalo. Stoga, kako bi se mišićno-koštani sustav dugo održao u zdravom stanju, potrebno je stalno trenirati, izvoditi razne psihička vježba. Svaka osoba nakon 30 godina osuđena je na tjelesni odgoj.
To i odgovarajuće preuređenje malih zglobova ruku potaknulo je korištenje alata, što je također ubrzalo razvoj mozga, koji je bio najvažniji organ antropoidne prilagodljivosti. Bilateralnost je posebno naglasila razvoj motoričkog sustava i ujedno pridonijela, budući da uspravan položaj tijela zahtjeva puno više motoričke koordinacije nego stajanje na četiri noge.
Širenje udova također je promijenilo mišiće. U puno većem dostupnom prostoru dopušteno je i umnožavanje i produžavanje udova. Pomažu u povećanju snage mišića i izvedbe manjih pokreta. Razvoj mehanizma kretanja dao je nove mogućnosti kralježnjacima. Njihovo kretanje postaje savršenije, brže, preciznije i prikladnije. Neophodno razvijene neuralne strukture koje kontroliraju funkciju skeletnih mišića. Kretanje kretanja bilo je olakšano istodobnim razvojem osjetilnih organa.
Ovo je posebno važno za žene, čije su kosti manje gustoće nego kod muškaraca. Osim toga, žene su sklonije gubitku mišićne mase u starijoj dobi.
Koštana masa, stupanj njezine formiranosti i njezin gubitak mogu se kontrolirati dobra prehrana. Kalcij i vitamin D jačaju kosti i sprječavaju njihovo pucanje, lomljenje i druge ozljede. Potrebna dnevna doza kalcija je otprilike 1200-1500 mg za odrasle osobe. Žene, kao i muškarci, dobiju 50% svoje koštane mase do dobi od 20 godina. U kombinaciji s redovitom tjelovježbom, kalcij potiče zdravu, jaku koštanu masu. Između 20. i 30. godine koštana masa lagano raste, a nakon 30. godine počinje proces njezina gubitka.
Uz njihovu pomoć čovjek može dobiti sve potpunije i točnije podatke iz okoline. Kako bi iskoristili te mogućnosti, neuroni koji podržavaju motoričku koordinaciju su se umnožili u mozgu, a kako bi ostali aktivni, formirali su sve složenije i intenzivnije veze s osjetilnim organima. Aksijalne mišiće u početku su kontrolirali isključivo spinalni motorni neuroni. Nakon odvajanja pojedinih mišića kontrolu nad pojedinim mišićima preuzeo je mozak koji se razvija kod kralješnjaka i ima središnju kontrolnu ulogu.
U vezi s biološkom funkcijom vašeg tijela, posebnu pozornost zdrava prehrana treba dati ženama. Trudnoća i dojenje zahtijevaju dodatni kalcij. Ako krv žene ne sadrži potreban iznos ovog minerala, kosti postaju dodatni izvor kalcija, što u konačnici dovodi do njihovog slabljenja. pušenje i prekomjerna upotreba alkoholna pića također doprinose gubitku koštane mase. Hormon estrogen regulira ispiranje kalcija iz kostiju i tako stvara normalne uvjete za dugotrajan rast kostiju. Međutim, u razdoblju menopauze i postmenopauze žensko tijelo prestaje proizvoditi estrogen koji je glavni uzrok ubrzanog gubitka koštane mase. Redovita tjelovježba i povećana doza dnevnog unosa kalcija spriječit će ovaj proces u postmenstrualnom razdoblju. Osim uzimanja kalcija kao dodatka prehrani, preporuča se u svakodnevnu prehranu uključiti hranu koja sadrži ovaj element.
Prvi autocentar kod kralježnjaka bio je lokomotorni dio smješten u Mesencephalonu. Iako je evolucija evoluirala do novih područja mozga specijaliziranih za motoričku kontrolu, retikulospastični trakt postoji i kod ljudi. Osim kontrole mišićnog tonusa, uglavnom je uključen u izvođenje ritmičkih pokreta.
Tijekom evolucije, mozak kralježnjaka u početku dominira dijelom modernog moždanog debla. Kako su moždane funkcije postajale složenije, bilo je potrebno više živčanih stanica za njihovu kontrolu. Budući da u tom trenutku u mozgu nije bilo dovoljno mjesta, novi neuroni formirali su novu koloniju ispred do tada smještenog mozga. U sljedećem koraku formiraju se tri jezgre odgovorne za vid: dvije iza i jedna iznad srednjeg mozga. Prethodni kralješnjaci imali su tri oka. Informacije dvaju bočnih očiju obradile su dvije bazalokativne lateralne genikulate tjelešca, koje prelaze gornje parijetalne oči kroz gornju jezgru.
Na program Fizičke aktivnosti treba uključiti i trening s utezima. Važan faktor, određivanje zdravlja mišićno-koštanog sustava, je održavanje optimalne težine.
Posebnu pozornost treba obratiti na noge. Noge imaju vrlo važnu ulogu u mišićno-koštanom sustavu, kao i u zdravlju cijelog organizma. Pravilna njega iza nogu uključuje, uz druge higijenske postupke, pažljiv odabir cipela. Udobne i pravilno odabrane cipele izvrsna su prevencija mnogih bolesti stopala.
Bočne oči i njihove jezgre su se također sačuvale kod čovjeka kao vidni organi, dok se istovremeno tjemeno oko i njegova jezgra postupno povukla, izgubila vid i poput epitalamusa tonula u dubinu. Sljedeća funkcija u dienecephalonu bila je somatosenzorna. U gmazova se liječenje somatosenzorne funkcije nastavilo razvijati; razvijene dorzalne i prednje jezgre talamusa. U isto vrijeme, anteroventralna jezgra bila je prvi koordinacijski centar za mezencefalon.
Posljednje područje talamusa koje se razvilo bio je Pulvinar Talami, koji je upravljao koordinacijom između osjetilnih organa. Kod nesisavaca preuzeo je cijelu ulogu tri osjetilna režnja mozga. Bazalni gangliji, koji koordiniraju sustav kretanja, već su pronađeni kod vodozemaca. Brzo su evoluirali, ali samo kod sisavaca.
"Mišićno-koštani sustav" i drugi članci iz rubrike Bolesti mišićno-koštanog sustava
Funkcionalna struktura voljnog kretanja. Iz navedenog proizlazi da su različite komponente uključene u osiguranje bilo kakvog kretanja, pa je jedno od glavnih pitanja kako osigurati jednokratnu naredbu koju prima izvršni aparat. Bez obzira na strategiju i taktiku pojedinog pokreta, glavna zadaća sustava koji osigurava program je koordinacija svih komponenti tima.
Naslage uranskog telencefalona, koje su se proširile s bočnih stijenki dienske ćelije, već se stvaraju kod vodozemaca. Prve telencefalne strukture odgovaraju biljkama bazalnih ganglija i limbičkom sustavu sisavaca. Njihovi glavni zadaci bili su njuh i koordinacija određenih, posebice seksualnih ponašanja. Cerebralni korteks je "izum" sisavaca. Prema gore navedenim osnovama nalaze se takve nove funkcije čiji neuroni nisu našli dovoljno mjesta u diencecephalonu.
Kako dienecephalon više nije mogao rasti, novi su neuroni nastali iz stijenke neuroepitelne cijevi koja je izrasla iz njezine stijenke. Postupno su se razvile lateralne klijetke, a iz njihovih stijenki režnjevi encefalona. Prvi sisavci bili su mali štapićasti glodavci koji su u početku skakali okolo, a zatim su postali vrećicasti. U doba dinosaura sklonili su se u špilje u podzemnoj željeznici. U skladu sa svojim načinom života, u mračnim špiljama kretali su se prvenstveno pomoću mirisa.
CNS ima niz genetski fiksiranih programa (na primjer, lokomotorni program hodanja temeljen na aktivnosti spinalni - koji se odnosi na kralježnicu.");" onmouseout="nd();" href="javascript:void(0);">spinalna generator). Takvi jednostavni programi kombiniraju se u složenije sustave poput održavanja uspravnog stava. Ova povezanost nastaje kao rezultat učenja, koje se postiže sudjelovanjem prednjeg korteksa. hemisfere.
Najsloženija i filogenetski najmlađa je sposobnost formiranja niza pokreta i predviđanja njegove provedbe. Rješenje ovog problema povezano je s frontalnim asocijativnim sustavom koji takve sekvence pokreta pamti i pohranjuje u memoriju. Najviši odraz tog kodiranja u čovjeku je verbalizacija, odnosno verbalna pratnja osnovnih pojmova kretanja.
Opća pravilnost sustava upravljanja kretanjem je korištenje povratne veze. To uključuje ne samo proprioceptivnu povratnu informaciju od pokreta koji je započeo, već također Aktivacija - uzbuđenje ili povećana aktivnost, prijelaz iz stanja mirovanja u aktivno stanje. onmouseout="nd();" href="javascript:void(0);">aktivacija sustavi nagrađivanja ili kažnjavanja. Osim toga, uključena je i interna povratna informacija, tj. informacije o aktivnosti osnovnih razina motoričkog sustava ili eferentna kopija same motoričke naredbe. Ova vrsta povratne sprege nužna je za razvoj novih motoričkih koordinacija. Za pokrete različite složenosti i brzine, povratna sprega može se zatvoriti na različitim razinama. Stoga obje vrste upravljanja - programiranje i praćenje - mogu koegzistirati u istom sustavu upravljanja kretanjem.
Zaključno, preporučljivo je citirati izjavu izvanrednog fiziologa N.A. Bernsteina da pokreti "...nisu prostorni, već semantički, a motoričke komponente lanaca razine akcije diktirane su i odabrane prema semantičkoj biti predmeta i onoga što treba učiniti s njim."
Za preživljavanje dvije najvažnije funkcije su miris i reprodukcija. Stoga su ove dvije funkcije riješene u dvije nove regije u razvoju od dienecephalon-a. njihovi ostaci, limbički sustav i mirisni korteks, koji obavljaju iste funkcije, također se nalaze kod ljudi u medijalnom središnjem korteksu. Ove su dvije funkcije usko povezane morfološki i funkcionalno kod svakog kralježnjaka, čak i kod ljudi sa znatno smanjenim mirisom. Industrija parfema temelji se na ovom stavu.
Hemisfere corpus callosuma ne postoje u mozgu najstarijih preživjelih sisavaca, kloakosa i ptica. Sljedeći korak u evoluciji bio je značajan razvoj somatosenzornog sustava. Ovi noćni glodavci, slični današnjim štakorima i miševima, još uvijek su živjeli u mraku pod zemljom. Sa svojim su se mogli brzo i pouzdano kretati mračnim podzemnim kanalima duga kosa na licu, jer su ih mogli koristiti kao radar, preko vrlo osjetljivih receptora pritiska povezanih. Za ovu preciznu mehaničku "analizu slike" trebala im je nova populacija neurona.
10.4. Elektrofiziološki korelati organizacije pokreta
Elektrofiziološke metode se koriste za proučavanje različitih aspekata motorna aktivnost, a posebno one koje su nedostupne neposrednom promatranju. Vrijedne informacije o fiziološkim mehanizmima organizacije pokreta daju metode za procjenu međudjelovanja zona moždane kore, analiza lokalnog EEG-a i potencijala vezanih uz kretanje te snimanje neuronske aktivnosti.
Proučavanje međuzonskih veza moždanih biopotencijala omogućuje praćenje dinamike interakcije pojedinih kortikalnih zona u različitim fazama izvođenja pokreta, pri učenju novih motoričkih vještina, da se identificiraju specifičnosti međuzonske interakcije tijekom različiti tipovi pokreta.
Ovi su neuroni migrirali u novoformirani teleencefalon i formirali prvi pravi režanj - tjemeni režanj. Taj je režanj također ostao somatosenzorni centar u čovjeka. Kod oposuma i sličnih vrsta, moždana kora se uglavnom koristi za liječenje kože. Kod starih sisavaca signali su išli od mišićnih vretena do novorazvijenog malog mozga, a odatle kroz jezgre malog mozga do talamusa.
Kao sljedeća faza evolucije, mozgovi zecolikih glodavaca, koji su se već probijali do površine zemlje, suočili su se s novim izazovom: morali su slušati iz daljine i pronaći potencijalnog napadačkog neprijatelja samo kako bi pobjegli u vrijeme. Novo akustično središte prešlo je iz donjeg kolikulusa u temporalni režanj, koji se razvija iz stijenke neuroepitelne cijevi, koja izrasta iz dienozifalona i ljulja se prema dolje zbog nedostatka prostora. dijelovi lateralna klijetka, koji sadrži dvije vaskularne stijenke pleksusa, stanični medij i donji dio kukuruza, kao i parijetalni i temporalni režanj, koji se razvijaju iz njegove stijenke, najstariji su dijelovi moždane kore.
Prostorna sinkronizacija (PS), tj. Sinkrona dinamika električnih oscilacija snimljenih iz različitih točaka cerebralnog korteksa odražava stanje moždanih struktura, što olakšava širenje ekscitacije i stvara uvjete za međuzonsku interakciju. Metodu registracije PS razvio je izvrsni ruski fiziolog M.N. Livanov.
Istraživanja ritmičkih komponenti EEG-a pojedinih zona i njihovih prostorno-vremenskih odnosa kod ljudi tijekom izvođenja voljnih pokreta dala su prava prilika pristupiti analizi središnjih mehanizama funkcionalnih interakcija koje se razvijaju na razini sustava tijekom motoričke aktivnosti. Korelacijska analiza EEG-a snimljenog tijekom izvođenja ritmičkih pokreta pokazala je da kod ljudi u kortikalnoj organizaciji pokreta sudjeluju ne samo centri motoričkog korteksa, već i frontalne i donje parijetalne zone.
Učenje voljnih pokreta i njihovo uvježbavanje uzrokuje preraspodjelu intercentralnih korelacija kortikalnih biopotencijala. Na početku treninga ukupni broj centara uključenih u aktivnost zglobova naglo se povećava, a sinkroni odnos ritmičkih komponenti EEG-a motoričkih zona s prednjim i stražnjim Asocijativne zone korteksa - zone koje primaju informacije od receptora koji percipiraju iritaciju različitih modaliteta, te od svih projekcijskih zona. ");" onmouseout="nd();" href="javascript:void(0);">asocijativne područja. Kako se pokret svladava, ukupna razina PS značajno opada, a naprotiv, povećavaju se veze između motoričkih zona i donjih parijetalnih. 
Važno je napomenuti da se u procesu učenja ritmički sastav biopotencijala različitih kortikalnih zona preuređuje: spori ritmovi počinju se bilježiti u EEG-u, koji se po frekvenciji podudaraju s ritmom izvođenja pokreta. Ovi ritmovi u ljudskom EEG-u nazivaju se "označeni". Iste označene oscilacije pronađene su u djece predškolska dob kada rade ritmičke pokrete na ergografu.
Sustavna istraživanja ljudskog EEG-a tijekom provedbe cikličke (povremeno ponavljajuće) i acikličke motoričke aktivnosti otkrila su značajne promjene u dinamici električne aktivnosti cerebralnog korteksa. U EEG-u dolazi do povećanja lokalne i udaljene sinkronizacije biopotencijala, što se izražava u povećanju snage periodičnih komponenti, u promjenama frekvencijskog spektra auto- i kros-korelograma, u određenom usklađivanju maksimumi frekvencijskih spektara i funkcija Koherencija - stupanj sinkronizacije EEG frekvencijskih pokazatelja između različitih dijelova cerebralnog korteksa.");" onmouseout="nd();" href="javascript:void(0);">koherentnost na istoj frekvenciji.
PS i vrijeme reakcije. Vrijeme reakcije jedan je od najjednostavnijih motoričkih pokazatelja. Stoga je posebno zanimljiva činjenica da čak i jednostavna motorička reakcija može imati različite fiziološke korelate ovisno o produljenju ili smanjenju njezina trajanja. Dakle, uspoređujući sliku međucentralnih korelacijskih odnosa spektralnih komponenti EEG-a mozga s vremenom jednostavne motoričke reakcije, pokazalo se da je preuređenje prostorno-vremenskih odnosa EEG-a asocijativnih zona povezano s odgovorom vremena na dati podražaj. Kod brzih reakcija u zdrave osobe najčešće su se javljale visoke korelacije biopotencijala u oba donja parijetalna područja (nešto više s lijevom hemisferom mozga). Ako se vrijeme reakcije poveća, to je popraćeno sinkronizacijom biopotencijala u frontalne regije korteks i donja parijetalna regija lijeve hemisfere bila je isključena iz interakcije. Osim toga, pronađen je odnos između veličina faznih pomaka Alfa ritam - glavni ritam elektroencefalograma u stanju relativnog mirovanja, s frekvencijom u rasponu od 8 - 14 Hz i prosječnom amplitudom od 30 - 70 μV. ");" onmouseout="nd();" href="javascript:void(0);">ritam alfa registriran u frontalnom, precentralnom i okcipitalna područja mozga i brzinu jednostavne motoričke reakcije.
Važno je napomenuti da se povećanje sinkronizacije biopotencijala događa kod osobe već u predradnom stanju u procesu koncentracije prije motoričke akcije, kao i tijekom mentalnog izvođenja pokreta.
PS i specifičnosti pokreta. Uz nespecifično povećanje PS biopotencijala, zabilježeno je njegovo izraženo selektivno povećanje između kortikalnih zona izravno uključenih u organizaciju određenog motoričkog akta. Na primjer, najveća sličnost u električnoj aktivnosti utvrđena je: pri kretanju ruku - između frontalne regije i motoričke reprezentacije mišića gornji udovi; pri kretanju nogu – između frontalne regije i motoričke reprezentacije mišića donjih ekstremiteta. Preciznim radnjama koje zahtijevaju finu prostornu orijentaciju i vizualnu kontrolu (gađanje, mačevanje, košarka), poboljšavaju se interakcije između vizualnih i motoričkih područja.
Složena dinamika PS biopotencijala različitih dijelova mozga kod sportaša otkrivena je tijekom izvođenja razne vježbe i prikazana je ovisnost povećanja interakcije ritmičkih komponenti EEG-a o načinu motoričke aktivnosti, o kvalifikacijama sportaša, o sposobnosti osobe da riješi taktičke probleme, o složenosti situacije. Stoga su kod visokokvalificiranih sportaša međucentralne interakcije mnogo intenzivnije i jasnije lokalizirane. Također se pokazalo da složeniji motorički zadaci zahtijevaju više visoka razina prostorna sinkronizacija EEG ritmova, a vrijeme rješavanja taktičkih zadataka korelira s brzinom porasta intercentralnih interakcija. U ovom slučaju motorički odgovor slijedi nakon postizanja maksimalne sinkroniziranosti biopotencijala u moždanoj kori.
Zajedno, studije PS biopotencijala mozga kod ljudi omogućile su da se utvrdi da prilikom izvođenja jednostavnih i složenih motoričkih radnji različiti centri mozga stupaju u interakciju, tvoreći složene sustave međusobno povezanih zona s žarištima aktivnosti ne samo u projekciji, već ali i u asocijativnim područjima, osobito frontalnom i donjem tjemenom. Ove međucentralne interakcije su dinamične i mijenjaju se u vremenu i prostoru kako se motorički čin izvodi.
10.5. Kompleks moždanih potencijala povezanih s pokretima
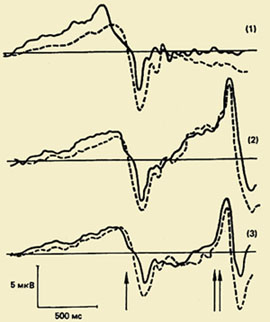 Jedan od važnih smjerova u proučavanju psihofiziologije motoričkog akta je proučavanje kompleksa oscilacija moždanih potencijala povezanih s pokretima (PMSD). Značaj ovog fenomena za razumijevanje fizioloških mehanizama organizacija pokreta je vrlo velika, jer proučavanje PMSD-a omogućuje otkrivanje skrivenog slijeda procesa koji se odvijaju u moždanoj kori tijekom pripreme i izvođenja pokreta, a kronometrija procesa obrade informacija je skup metoda za mjerenje trajanje pojedinih faza u procesu obrade informacija na temelju mjerenja fiziološki pokazatelji, posebno latentna razdoblja komponenti evociranih potencijala i potencijala povezanih s događajima.");" onmouseout="nd();" href="javascript:void(0);"> sat ovi procesi, tj. postaviti svoje vremenske granice.
Jedan od važnih smjerova u proučavanju psihofiziologije motoričkog akta je proučavanje kompleksa oscilacija moždanih potencijala povezanih s pokretima (PMSD). Značaj ovog fenomena za razumijevanje fizioloških mehanizama organizacija pokreta je vrlo velika, jer proučavanje PMSD-a omogućuje otkrivanje skrivenog slijeda procesa koji se odvijaju u moždanoj kori tijekom pripreme i izvođenja pokreta, a kronometrija procesa obrade informacija je skup metoda za mjerenje trajanje pojedinih faza u procesu obrade informacija na temelju mjerenja fiziološki pokazatelji, posebno latentna razdoblja komponenti evociranih potencijala i potencijala povezanih s događajima.");" onmouseout="nd();" href="javascript:void(0);"> sat ovi procesi, tj. postaviti svoje vremenske granice.
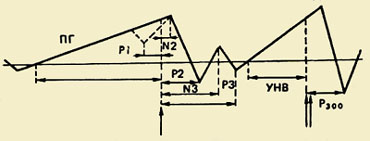
Komponentni sastav PMSD-a. Prvi put je ovaj kompleks, koji odražava procese pripreme, izvođenja i evaluacije pokreta, registriran 60-ih godina. Pokazalo se da pokretu prethodi spora negativna oscilacija - potencijal spremnosti (RP). Počinje se razvijati 1,5 - 0,5 s prije početka kretanja. Ova se komponenta registrira uglavnom u središnjim i fronto-centralnim odvodima obiju hemisfera. 500-300 ms prije početka kretanja, PG postaje asimetričan - njegova najveća amplituda se opaža u precentralnom području, Kontralateralno - odnosi se na suprotnu stranu tijela.");" onmouseout="nd();" href="javascript:void(0);"> kontralateralni pokret. Otprilike polovica odraslih ispitanika, na pozadini ove spore negativne oscilacije, neposredno prije početka kretanja, bilježi malu amplitudu pozitivne komponente. Naziva se "premotorna pozitivnost" (PMP). Sljedeća brzo rastuća negativna oscilacija po amplitudi, tzv. motorički potencijal (MP), počinje se razvijati 150 ms prije početka pokreta i dostiže svoju maksimalnu amplitudu u području motoričke reprezentacije pokretnog ekstremiteta u kori velikog mozga. Ovaj kompleks potencijala završava pozitivnom komponentom otprilike 200 ms nakon početka kretanja.
Funkcionalni značaj komponenti. Opće je prihvaćeno da potencijal spremnosti (RP) nastaje u motoričkom korteksu i povezan je s procesima planiranja i pripreme pokreta. Spada u klasu sporih negativnih oscilacija moždanog potencijala, čija se pojava objašnjava aktivacijom neuronskih elemenata odgovarajućih područja korteksa.
Hipoteze u vezi funkcionalna vrijednost PMP su različiti.
Ova se oscilacija smatra i odrazom isporuke središnje naredbe iz korteksa mišićima, i rezultatom opuštanja korteksa nakon završetka određene faze organizacije pokreta, te kao odraz procesa potiskivanja povezanih pokreta drugog uda, i kao povratna informacija od aferenata mišića. Trenutno neki autori vjeruju da su PMP samo odraz početka motoričkog potencijala.
Prilikom registracije MP u majmuna identificirane su dvije podkomponente u MP. Prva podkomponenta je u korelaciji s aktivacijom motoričkog korteksa povezanog s inicijacijom pokreta (sinaptička aktivnost piramidalni neuroni), a drugi - s aktivacijom polja 2,3 i 4 Brodmannova polja - odvojena područja moždane kore, koja se razlikuju po staničnoj strukturi (citoarhitektonici) i funkcijama. Na primjer, polja 17,18,19 su vidna područja cerebralnog korteksa, koja imaju različite strukture i funkcije u pružanju vizualne percepcije.");" onmouseout="nd();" href="javascript:void(0);">od Brodmana. Registriranje MP kod osobe s epilepsijom omogućilo je razlikovanje tri komponente u njemu. Prva komponenta nazvana je inicijacijski potencijal. Ima visoku amplitudu i javlja se nakon početka pokreta u precentralnom kontralateralnom korteksu. Drugi, koji se javlja nakon početka miograma i više je lokaliziran u kontralateralnom somatosenzornom polju, može biti povezan i s inicijacijom pokreta i senzornom povratnom spregom. Treća komponenta odražava impuls koji dolazi od aferentnih mišića do korteksa.
Pozitivni potencijal koji slijedi nakon MP smatra se odrazom obrnute aferentacije koja dolazi od perifernih receptora, uzlazne aktivnosti iz motoričkih centara, operacije usporedbe između motoričkog programa i neuronske slike njegovog izvršenja ili kortikalnih procesa. opuštanje nakon pokreta.
Val iščekivanja. Uz PMSD opisan je još jedan elektrofiziološki fenomen koji je bitno blizak potencijalu spremnosti. Riječ je o o negativnoj fluktuaciji potencijala zabilježenoj u prednjim dijelovima cerebralnog korteksa u razdoblju između djelovanja upozoravajućeg i okidačkog (koji zahtijeva reakciju) signala. Ova fluktuacija ima nekoliko naziva: val očekivanja, E-val, uvjetno negativno odstupanje (CNV). E-val se javlja 500 ms nakon signala upozorenja, njegovo trajanje raste s povećanjem intervala između prvog i drugog podražaja. Amplituda E-vala raste izravno proporcionalno brzini motoričkog odgovora na početni podražaj. Pojačava se s napetošću pažnje i povećanjem voljnog napora, što ukazuje na povezanost ovog elektrofiziološkog fenomena s mehanizmima voljne regulacije motoričke aktivnosti i ponašanja općenito.
10.6. neuralna aktivnost
Funkcionalni kortikalni stupovi. U motoričkoj zoni ljudskog korteksa postoje takozvani ogromni piramidalni Beza stanice – piramidalne stanice kore velikog mozga. onmouseout="nd();" href="javascript:void(0);">Betzove ćelije, koji su organizirani u različite stupce. Piramidalne stanice koje obavljaju slične funkcije smještene su jedna do druge, inače bi bilo teško objasniti točnu somatotopsku organizaciju korteksa. Takvi motorički stupovi sposobni su pobuditi ili inhibirati skupinu funkcionalno homogenih Motorni neuron (motorni neuron) – živčana stanica čiji akson inervira mišićna vlakna. onmouseout="nd();" href="javascript:void(0);">motorni neuroni.
Registriranje aktivnosti pojedinačnih piramidalnih stanica pomoću implantiranih mikroelektroda kod životinja koje izvode različite pokrete omogućilo je utvrđivanje temeljno važne činjenice. Kortikalni neuroni koji reguliraju aktivnost bilo kojeg mišića nisu koncentrirani unutar samo jednog stupca. Motorni stup je uglavnom funkcionalna asocijacija neurona koji reguliraju aktivnost nekoliko mišića koji djeluju na određeni zglob. Dakle, u stupcima piramidalnih neurona motornog korteksa pokreti su zastupljeni ne toliko koliko mišići.
Neuralni kodovi motoričkih programa. Kodiranje informacija u neuronu provodi se učestalošću njegovih pražnjenja. Analiza impulsne aktivnosti neurona tijekom razvoja različitih motoričkih programa kod životinja pokazala je da neuroni sudjeluju u njihovoj izgradnji. različitih odjela motornog sustava, dok obavlja određene funkcije. Prema nekim idejama, uključivanje motoričkih programa događa se zbog aktivacije takozvanih komandnih neurona. Zapovjedni neuroni su pak pod kontrolom viših kortikalnih centara. Inhibicija je živčani proces suprotan ekscitaciji; očituje se u slabljenju ili prestanku aktivnosti specifične za određeni sustav tijela. onmouseout="nd();" href="javascript:void(0);">Uspori naredbeni neuron dovodi do zaustavljanja programa kojim upravlja, ekscitacija, naprotiv, do aktivacije živčanog kruga i aktualizacije motoričkog programa.
Uključenost komandnih neurona u cjelovitu aktivnost mozga određena je trenutnom motivacijom i specifičnim motoričkim programom usmjerenim na zadovoljenje te motivacije. Motorički program, da bi bile adaptivne prirode, moraju uzeti u obzir sve signalno značajne komponente vanjskog okruženja, u odnosu na koje se vrši svrhovito kretanje, tj. temeljen na principu multisenzora Konvergencija je spajanje aksona skupine neurona, koje nastaje zbog stvaranja sinapsi na istom postsinaptičkom neuronu. onmouseout="nd();" href="javascript:void(0);">konvergencija.
Rječnik pojmova
- kretanje
- motorički neuron
- artikulacija
- aktiviranje
- koherentnost
- Brodmannova polja
- kronometrija procesa obrade informacija
Pitanja za samoispitivanje
- Koja je razlika između statične i dinamičke slike tijela?
- Koje strukture mozga igraju odlučujuću ulogu u organizaciji voljnog kretanja?
- Kako se razlikuju funkcije piramidnog i ekstrapiramidnog sustava?
- Što potencijali mozga povezani s kretanjem daju za razumijevanje moždanih mehanizama organizacije kretanja?
Bibliografija
- Batuev A.S. Funkcije motoričkog analizatora. L.: Nauka, 1970.
- Batuev A.S. viši živčana aktivnost. Moskva: Viša škola, 1991.
- Bernstein N.A. Ogledi o fiziologiji pokreta i fiziologiji aktivnosti. Moskva: Nauka, 1966.
- Bloom F., Lizerson A., Hofstadter L. Mozak, um i ponašanje. M.: Mir, 1988.
- Dudel J., Ruegg I., Schmidt R., Janig V. Ljudska fiziologija. T. 1 / Ed. R. Schmidt i G. Thevs. M.: Mir, 1985.
- Opći tečaj fiziologije čovjeka i životinja / Ed. A.A. Nozdračev. Moskva: Viša škola, 1991.
- Sologub E.B. Električna aktivnost ljudskog mozga u procesu motoričke aktivnosti. L.: Nauka, 1973.
- Khizman T.P. Kretanje djeteta i električna aktivnost mozga. Moskva: Pedagogija, 1973.
- Evarts E. Moždani mehanizmi koji kontroliraju kretanje // Brain. M.: Mir, 1982.
Teme seminarskih radova i eseja
- Učenje N.A. Bernstein o strukturi pokreta.
- Psihofiziološke tipologije pokreta.
- Pokreti ljudske ruke i njihovi neurofiziološki mehanizmi.
- Voljni pokreti i njihovi neurofiziološki mehanizmi.
- Povijesne faze istraživanja moždanih potencijala povezanih s pokretima.
- Uloga neuralne aktivnosti u izgradnji pokreta.

