14.1. OPĆE ODREDBE
Krajnji mozak (telencephalon), ili veliki mozak (cerebrum), koji se nalazi u supratentorijalnom prostoru lubanjske šupljine sastoji se od dvije velike
hemisfere (gemispherium cerebralis),odvojene dubokim uzdužnim prorezom (fissura longitudinalis cerebri), u koji je uronjen srp veliki mozak (falx cerebri) predstavljajući duplikaciju čvrstog moždane ovojnice. Velike hemisfere mozga čine 78% njegove mase. Svaki od hemisfere Ima režnjevi: frontalni, parijetalni, temporalni, okcipitalni i limbički. Obuhvaćaju strukture diencefalona te moždanog debla i malog mozga koji se nalaze ispod cerebelarnog plašta (subtentorijalno).
Iako ljudski mozak kao cjelina može obavljati mnoge zadatke i koncentrirati se na specifičnije od drugih, istina je da je ljudski mentalni aparat specijaliziraniji, odnosno da su dijelovi svake hemisfere usmjereni na određene zadatke.U slučaju žena , postoji integralnija uporaba obiju hemisfera više aktivnosti, što se naziva multitasking.
Segmentacija informacija
U muškom mozgu informacije su podijeljene u skupine ili segmente koji nisu nužno povezani. Kod žena je sve povezano kao sustav. Zbog toga su žene holističke. Zbog ovih razlika između mozgova muškaraca i žena, muškarci imaju tendenciju da se potpuno usredotoče na aktivnosti koje obavljaju, a da im pritom ne odvlače pažnju drugi aspekti. S druge strane, žene svaki događaj na svijetu promatraju kao cjeloviti crtež u kojem je svaki dio u interakciji.
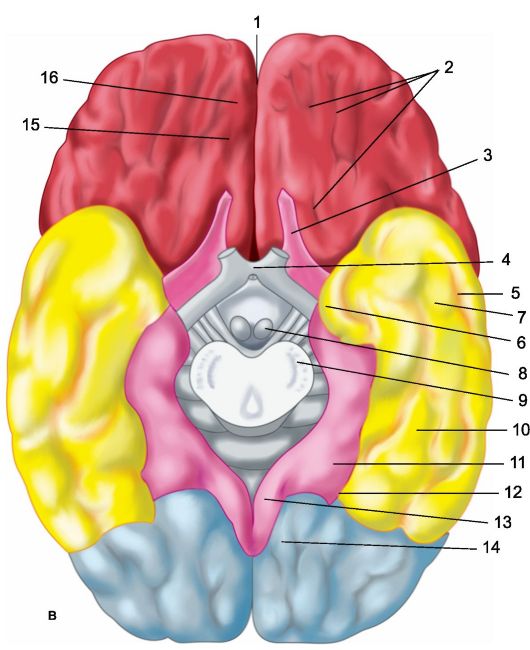
Svaka od moždanih hemisfera ima tri površine: gornji bočni, ili konveksilni (slika 14.1a), - konveksan, okrenut prema kostima lubanjskog svoda; unutarnji (Sl. 14.1b), uz veliki falciformni nastavak, i donji, ili bazalni (Sl. 14.1c), ponavljajući reljef baze lubanje (prednje i srednje jame) i cerebelarnog tena. U svakoj hemisferi razlikuju se tri ruba: gornji, donji unutarnji i donji vanjski, te tri pola: prednji (frontalni), stražnji (okcipitalni) i bočni (temporalni).
Osjećaji muškarca i žene
Također u osjećajima i percepciji svijeta postoje razlike između muškarca i žene. Kod čovjeka je vid dominantna funkcija, način na koji većina informacija ulazi u živčani sustav. Kod žena svi sustavi rade zajedno kao kod slijepaca, imaju jako razvijena osjetila sluha, njuha i dodira, ali su im dodani vid.
Veličina mozga također se razlikuje između muškaraca i žena i obično je malo veća. To ne znači da su pametniji, jer znanstvenici smatraju da je svrha ove ekspanzije povezana s velikim volumenom muških mišića, a ne s mentalnim procesima. Muškarci imaju 20 puta više testosterona u tijelu nego žene. Zbog toga se njegov živčani sustav očituje u agresivnijim i dominantnijim odnosima te je seks fizički poželjniji. Žene su delikatnije i njihovi umovi traže seksualno zadovoljstvo usredotočeno na senzualnost i ljubav.
Šupljina svake hemisfera je lateralna komora mozga dok je lijeva lateralna klijetka prepoznata kao prva, desna - druga. Lateralni ventrikul ima središnji dio smješten duboko u parijetalnom režnju (lobus parietalis) i tri roga izlaze iz njega: prednji rog prodire u frontalni režanj (lobus frontalis), donji - do temporalnog (lobus temporalis), stražnji - u okcipitalnom (lobus occipitalis). Svaki od lateralnih ventrikula komunicira s trećom moždanom komorom preko interventrikularnog rupa Monroe.
Hemisfere mozga i kognitivne sposobnosti
I na kraju ćemo reći da kod ljudi lijeva hemisfera mozga ima veću dominaciju. Kod žena postoji ravnoteža u korištenju obje hemisfere, lijeve i desne. To je razlog zašto u kognitivnim vještinama možemo primijetiti da muškarci imaju veliku tendenciju da budu dobri u matematici i računanju, dok žene imaju sposobnost jezika, aktivnosti koja zahtijeva korištenje nekoliko područja unutar sebe.
Oni nam pokazuju kako, u evolucijskom procesu, funkcionira živčani sustav oba su spola bila specijalizirana kako bi osigurala budućnost vrste. Nikada nemojte zaboraviti da su razlike povezane s fizičkim i mentalnim strukturama te da ni jedno ni drugo ne treba podcjenjivati ljudsku vrijednost svakog od žanrova. Različiti smo, ali se nadopunjujemo.
Središnji dijelovi medijalne površine obiju hemisfera međusobno su povezani cerebralnim komisurama, od kojih je najmasivniji corpus callosum, i strukturama diencefalona.
Telencefalon se, kao i drugi dijelovi mozga, sastoji od sivih i bijela tvar. Siva tvar nalazi se u dubini svake hemisfere, tvoreći tamo subkortikalne čvorove, te duž periferije slobodnih površina hemisfere, gdje čini koru velikog mozga.
U knjizi Nedostatak pažnje kod djece i adolescenata, jedna od stvari koju autorica Amanda Cespedes spominje je važnost poznavanja mozga i njegovih funkcija kao strategije za razumijevanje i odgovaranje na potrebe djece i adolescenata. Zapravo, učenjem se stječu nova znanja. Da bi se to postiglo, postoje neke osnovne funkcije mozga koje dolaze u igru od kognitivnih do emocionalnih. Konkretno, studija ljudski mozak u djetinjstvu se temelji na postojanju neuronske mreže u kojem se informacije obrađuju na isti način kao i računalo pri obradi podataka.
Osnovna pitanja vezana uz građu, funkcije bazalnih ganglija i varijante klinička slika pri njihovom porazu razmatraju se u poglavljima 5, 6. Moždana kora je otprilike
Riža. 14.1.Hemisfere mozga.
To znači da djetetov mozak nije ograničen samo na primanje informacija, au tom procesu stotine tisuća neurona postaju dostupni za učenje i informacijski proces koji se odvija u dvije hemisfere mozga: desnoj i lijevoj. U knjizi The Disability of ADHD in Children and Adolescents autorice Amande Cespedes, autorica se dublje bavi ovom temom, ističući karakteristike svake hemisfere i dajući konkretni primjeri situacije ili radnje u kojima se očituje funkcioniranje obiju hemisfera kod djece i učitelja, Zašto?
Budući da se samo dubokim razumijevanjem mozga mogu oblikovati strategije učenja koje odgovaraju specifičnim potrebama. Sazrijeva prije rođenja i jača svoje funkcionalne mreže tijekom prvih pet godina života. U ovoj hemisferi postoji šest talenata: vizualni, prostorna logika, melodijski ritam, kinesico, interpersonalni i naturalistički. Desna polutka nije reguliran objektivnom analizom vanjske stvarnosti i stoga dopušta postojanje subjektivnih stvarnosti pogodnih za maštu.
a - gornja bočna površina lijeve polutke: 1 - središnji sulkus; 2 - orbitalni dio donjeg frontalnog gyrusa; I - frontalni režanj; 3 - precentralni gyrus; 4 - precentralna brazda; 5 - gornji frontalni girus; 6 - srednji frontalni gyrus; 7 - tegmentalni dio inferiornog frontalnog gyrusa; 8 - donji frontalni gyrus; 9 - bočna brazda; II - parijetalni režanj: 10 - postcentralni gyrus; 11 - postcentralna brazda; 12 - intraparietalni žlijeb; 13 - supramarginalni gyrus; 14 - kutni gyrus; III - temporalni režanj: 15 - gornji temporalni gyrus; 16 - gornji temporalni sulcus; 17 - srednji temporalni gyrus; 18 - srednji temporalni sulcus; 19 - donji temporalni gyrus; IV - okcipitalni režanj: b - medijalna površina desne hemisfere: 1 - paracentralni režanj, 2 - precuneus; 3 - parieto-okcipitalni sulkus; 4 - klin, 5 - jezični gyrus; 6 - bočni okcipitotemporalni girus; 7 - parahipokampalni girus; 8 - kuka; 9 - svod; 10 - corpus callosum; 11 - gornji frontalni girus; 12 - cingularni gyrus; c - donja površina moždanih hemisfera: 1 - uzdužna interhemisferna pukotina; 2 - orbitalne brazde; 3 - mirisni živac; 4 - optički chiasm; 5 - srednji temporalni sulkus; 6 - kuka; 7 - donji temporalni gyrus; 8 - mastoidno tijelo; 9 - baza moždanog debla; 10 - bočni okcipitotemporalni girus; 11 - parahipokampalni girus; 12 - kolateralni utor; 13 - cingularni gyrus; 14 - jezični gyrus; 15 - mirisni utor; 16 - izravni gyrus.
Kognitivni stil ovog područja mozga je heterogenog tipa, što znači da se kreće od početne ideje do krajnje točke koja je nejasno povezana s ovom početnom idejom. Intuicija i holističko procesiranje vrlo su važni u ovoj hemisferi. Razumijevanje funkcioniranja ove strane mozga korisno je za kreiranje pedagoške prakse temeljene na tome, pa autorica detaljno i jednostavno opisuje aktivnosti koje se pojavljuju na ovoj strani djetetova mozga i strategije koje učitelj primjenjuje kako bi tu istu iskoristio. hemisfera.
Sazrijeva nakon šest mjeseci života, a glavno sazrijevanje završava u pubertetu. Talenti koji se temelje na ovoj strani mozga su: lingvistička, matematička simbolička logika i interpersonalna ili refleksivna. Lijeva hemisfera dovodi dijete u kontakt s pojmovnim znanjem, no takvo je učenje krhko i podložno zaboravu. Može se reći da njihova konsolidacija favorizira podršku desne hemisfere. Kao iu desnoj hemisferi, Amanda navodi radnje koje se pojavljuju na ovoj strani mozga djeteta i učitelja.
3 puta veća od površine hemisfera vidljive tijekom vanjskog pregleda. To je zbog činjenice da je površina moždanih hemisfera naborana, ima brojne depresije - brazde (sulci cerebri) a nalazi se između njih vijuge (gyri cerebri). Cerebralni korteks pokriva cijelu površinu vijuga i brazdi (stoga je njegovo drugo ime palij - plašt), dok ponekad prodire do velike dubine u supstancu mozga.
Naučite mijenjati brojeve kako biste formirali brojeve za izvođenje operacija i rješavanje problema. Razumijevanje, kaže autor, dizajna mozga u cjelini i njegovog izvršnog mehanizma znači imati uvjete za razumijevanje djeteta ili adolescenta i intervenirati u pedagoškim strategijama koje su prilagođene potrebama svakog učenika. Međutim, koliko znate o mozgu i kako to znanje primijeniti za stvaranje alata koji jamče ne samo učenje, već i izgradnju društvenog, školskog i obiteljskog života?
Što su režnjevi u hemisferama velikog mozga?
Činjenica je da su razgovori koje roditelji vode s djecom ključni u procesu učenja. Prema mnogim stručnjacima, to im pomaže u razvoju jezika, društvenih vještina i drugih stvari ključnih za razumijevanje svijeta oko sebe. U skladu s tim došla su nova otkrića koja naglašavaju važnost komuniciranja s ciljevima.
Ozbiljnost i položaj brazdi i vijuga moždanih hemisfera do određene su mjere varijabilni, međutim, glavni se formiraju u procesu ontogeneze i konstantni su, karakteristični za svaki normalno razvijeni mozak.
14.2. VELIKI KRETOVI I HVATIŠTA HEMISFERA MOZGA
Gornja lateralna (konveksilna) površina hemisfera (Slika 14.1a). Najveći i najdublji bočno brazda (sulcus lateralis),ili sylvian brazda, - odvaja frontalni i prednji dio parijetalnog režnja od onog ispod temporalni režanj. Frontalni i parijetalni režanj su odvojeni središnja, ili Rolandova, brazda(sulcus centralis), koji presijeca gornji rub hemisfere i ide dolje i naprijed duž svoje konveksilne površine, malo kraće od lateralne brazde. Parijetalni režanj je odvojen od okcipitalnog režnja koji se nalazi iza njega parijetalno-okcipitalnim i poprečnim okcipitalnim žljebovima koji prolaze duž medijalne površine hemisfere.
Otočić, limbički korteks
Prema riječima stručnjaka, neka istraživanja temeljena na promatranju obitelji pokazala su da razgovor o brojevima tijekom svakodnevnih aktivnosti i igara predviđa daljnje matematičko znanje djece. Osim toga, dokazi također navode da je ovo rano učenje dugoročno doprinosi razumijevanju matematike i jezika. Sada, iako su djeca rođena da uče brojeve, potrebna im je podrška kada pričamo o povezivanju imena brojeva sa točan broj naučiti kako prikazati različite brojeve objekata.
U frontalnom režnju ispred središnjeg girusa i paralelno s njim nalazi se precentral (gyrus precentralis), ili prednji središnji, gyrus, koji je sprijeda omeđen precentralnim sulkusom (sulcus precentralis). Gornji i donji frontalni žlijebovi polaze sprijeda od precentralnog sulkusa, dijeleći konveksitalnu površinu prednjih dijelova frontalnog režnja u tri frontalna vijuga - gornji, srednji i donji (gyri frontales superior, media et inferior).
Podrška za učenje brojeva
Kada roditelji vode ove razgovore sa svojom djecom, mogu saznati. Ovo razumijevanje broja znakova je stvarno važno kada počnu raditi s puno većim i složenijim količinama tijekom školski proces. Međutim, važno je da djeca znaju redoslijed brojeva, međutim, roditelji bi trebali vježbati sa svojom djecom mnogo više nego što se smatra. Ako se to ne učini, djetetu će biti teško razumjeti što nazivi brojeva znače i kako ih koristiti. Stoga stručnjaci nude neke ideje koje će ih podržati prije nego što se susretnu s matematikom u školi.
Prednji dio konveksilne površine parijetalnog režnja nalazi se iza središnjeg postcentralnog sulkusa (gyrus postcentralis), ili stražnji središnji, gyrus. Iza nje graniči postcentralni sulkus, od kojeg se unatrag proteže intraparijetalni sulkus. (sulcus intraparietalis), odvajajući gornji i donji parijetalni lobulus (lobuli parietales superior et inferior). U donjem parijetalnom režnju, pak, razlikuje se supramarginalni girus (gyrus supramarginalis), okružuje stražnji dio lateralnog (Sylvijevog) žlijeba i kutni girus (girus angularis), graniči sa stražnjim dijelom gornje temporalne vijuge.
Više od samog čitanja niz brojeva, broji i raspravlja o nizu vidljivih predmeta. Broji i klasificira broj objekata u skupu. To će omogućiti vašoj djeci da se povežu s postupkom brojanja u skladu s veličinom seta.
Koristite nazive brojeva koji su malo veći od brojeva koje vaše dijete već razumije. Pogledajte dječje pokrete brojevima. Mnogo puta ispravno reagiraju gestama, ali ne i riječima. Vaše geste su također korisne. Kad brojite brojeve s djecom, pokažite na svaki predmet koji je dio brojanja. Prstima pokažite količinu o kojoj govorite.
Na konveksitalnoj površini okcipitalnog režnja mozga, brazde su plitke i mogu značajno varirati, zbog čega je priroda vijuga između njih također promjenjiva.
Konveksitalnu površinu temporalnog režnja dijele gornji i donji temporalni sulkus, koji su gotovo paralelni s lateralnim (Silvijevim) sulkusom, dijeleći konveksitalnu površinu temporalnog režnja na gornji, srednji i donji temporalni vijug. (gyri temporales superior, media et inferior). Gornji temporalni girus tvori donju usnicu bočne (Sylvijeve) brazde. Na svojoj površini okrenutoj
Iskoristite količinu Svakidašnjica. Vježbajte brojanje dok čitate knjige. Zamolite djecu da odaberu određeni broj predmeta tijekom svojih dnevnih rutina. Usporedite broj tijekom igre. Razgovarajte sa svojom djecom o tome kako koristite brojeve u svakodnevnom životu. Na primjer, analizirajte kako ih koristite da biste znali što kupiti, planirali koliko ćete pojesti ili postavili stol.
Ako podržavamo proces čitanja, zašto to ne učiniti i s brojevima? Centar za razvoj djeteta na Sveučilištu Harvard nekoliko je puta govorio o važnosti prijemčive i obzirne interakcije između odraslih i djece. Ovo je, kažu istraživači centra, temeljno za izgradnju čvrstih temelja mozga za učenje i budući razvoj. Ova komunikacija ili interakcija naziva se "usluži i vrati" i ključna je za izgradnju temelja učenja i osiguravanje optimalne dobrobiti.
strana bočne brazde, postoji nekoliko poprečnih malih brazdi, ističući mali poprečni girus na njemu (Geschlov girus), što se može vidjeti samo širenjem rubova bočne brazde.
Prednji dio bočne (Silvijeve) brazde je udubina sa širokim dnom, koja tvori tzv. otok (otok) ili insula (lubus insularis). Gornji rub bočne brazde koja pokriva ovaj otok naziva se guma (operkulum).
Struktura temporalnog režnja
Sada, da bi se vježbala ova "igra" koja uključuje obje strane, postoje radnje ili koraci koji se mogu poduzeti, što je to? Promatrajte kada komunicirate s djecom. Gledate li nešto posebno ili pravite buku? Imate li poseban izraz lica? Mičete li rukama ili nogama? Te su stvari, kažu u centru, usluge i važno ih je pratiti tijekom dana u različitim situacijama jer tako mogu upoznati vještine, interese i potrebe djece. Tako ćete ih motivirati da istražuju i ojačat ćete vezu koju imaju.
Unutarnja (medijalna) površina hemisfere (Slika 14.1b). Središnji dio unutarnje površine hemisfere usko je povezan sa strukturama diencefalona, od kojih je omeđen strukturama koje se odnose na veliki mozak. svod (fornix) I Corpus callosum (Corpus callosum). Potonji je izvana obrubljen brazdom corpus callosuma (sulcus corporis callosi), počevši od njegove prednje strane – kljuna (govornica) a završava na svom zadebljanom stražnjem kraju (splenij). Ovdje sulcus corpus callosum prelazi u duboki hipokampalni sulcus (sulcus hippocampi), koji prodire duboko u supstancu hemisfere, pritiskajući je u šupljinu donjeg roga. lateralna klijetka, što rezultira stvaranjem takozvanog amonijevog roga.
Zagrljaji, nježne riječi, igra, izrazi lica i druge aktivnosti mogu biti način da uzvratite usluge koje djeca pružaju. Na taj način im se pruža podrška i motivira ih za daljnje služenje, što je, kako kažu, u središtu svojevrsna nagrada za dječju znatiželju i interes. To što od odrasle osobe ne dobivaju ništa zauzvrat može biti izvor stresa za njih. S druge strane, kada dobiju odgovor, možda znaju da su njihove misli i osjećaji, osim brige, važni.
Kada uzvraćate uslugu, važno je dati ime onome što djeca vide, osjećaju ili rade. Na taj ćete način stvoriti važne jezične veze na razini mozga, čak i prije nego što počnete razumjeti riječi i govoriti. Na primjer, ako dijete pokazuje na svoja stopala, možete izgovoriti izraze poput "Da, to su stopala." Zašto je to važno? Jer na taj način ćete mu pomoći da razumije svijet oko sebe i da oni shvate što mogu očekivati od njega. Imenovanje stvari također je način da im date riječi koje mogu koristiti i pokažete im da vam je stalo.
Donekle odlazeći od sulkusa corpus callosum i hipokampalnog sulkusa, nalaze se corpus callosum, subparietalni i nazalni sulci, koji se nastavljaju jedan na drugi. Ti utori izvana ograničavaju lučni dio medijalne površine hemisfere velikog mozga, poznat kao limbički režanj(lobus limbicus). Postoje dvije vijuge u limbičkom režnju. Gornji dio limbičkog režnja je gornji limbički (gornji rubni), ili pojas, gyrus (girus cinguli), donji dio tvori donji limbički girus, ili girus morskog konjica (girus hippocampi), ili parahipokampalnog girusa (girus parahypocampalis), ispred koje se nalazi kuka (uncus).
Oko limbičkog režnja mozga nalaze se tvorevine unutarnje površine frontalnog, parijetalnog, okcipitalnog i temporalni režanj th. Veći dio unutarnje površine frontalnog režnja zauzima medijalna strana gornjeg frontalnog girusa. Na granici između frontalnog i parijetalnog režnja nalazi se moždana hemisfera paracentralnog lobula (lobulis paracentralis), koji je, takoreći, nastavak prednjeg i stražnjeg središnjeg vijuga na medijalnoj površini hemisfere. Na granici između parijetalnog i okcipitalnog režnja jasno je vidljiva parijetalno-okcipitalna brazda. (sulcus parietooccipitalis). Od dna polazi natrag ostružna brazda (sulcus calcarinus). Između tih dubokih brazda nalazi se trokutasti girus, poznat kao klin. (cuneus). Ispred klina je četverokutna vijuga, povezana s tjemeni režanj mozak – prekuneus.
Donja površina hemisfere (Slika 14.1c). Donja površina hemisfere velikog mozga sastoji se od formacija frontalnog, temporalnog i okcipitalni režanj th. u susjedstvu središnja linija frontalni režanj je rectus gyrus (girus rectus). Izvana je ograničen olfaktornim žlijebom (sulcus olfactorius), na koje se odozdo naslanjaju tvorevine njušnog analizatora: olfaktorni bulbus i olfaktorni trakt. Bočno od njega, do bočne (Sylvijeve) brazde, koja se proteže do donje površine frontalnog režnja, nalaze se male orbitalne vijuge (gyri orbitalis). Bočne dijelove donje površine hemisfere iza lateralnog sulkusa zauzima inferiorni temporalni girus. Medijalno od njega je lateralna temporo-okcipitalna vijuga. (gyrus occipitotemporalis lateralis), ili fuziformni žlijeb. Prije-
njezini odjeli sa iznutra granice na gyrusu hipokampusa, a stražnji - na lingvalnom (gyrus lingualis) ili medijalni temporoccipitalni girus (gyrus occipitotemporalis medialis). Potonji, svojim stražnjim krajem, priliježe uz utor ostruge. Prednji dijelovi fusiformne i lingvalne vijuge pripadaju temporalnom režnju, a stražnji dijelovi okcipitalnom režnju mozga.
14.3. BIJELA TVAR VELIKIH HEMISFERA
Bijelu tvar hemisfera velikog mozga čine živčana vlakna, pretežno mijelin, čineći putove koji osiguravaju veze između neurona korteksa i klastera neurona koji tvore talamus, subkortikalne čvorove, jezgre. Glavni dio bijele tvari hemisfera velikog mozga nalazi se u njegovoj dubini poluovalno središte ili blistava kruna (zračeća korona), koji se uglavnom sastoji od aferentnih i eferentnih projekcija putovi koji povezuju cerebralni korteks sa subkortikalnim čvorovima, jezgrama i retikularnom supstancom diencefalona i moždanog debla, sa segmentima leđna moždina. Posebno su kompaktno smješteni između talamusa i subkortikalnih čvorova, gdje tvore unutarnju kapsulu opisanu u 3. poglavlju.
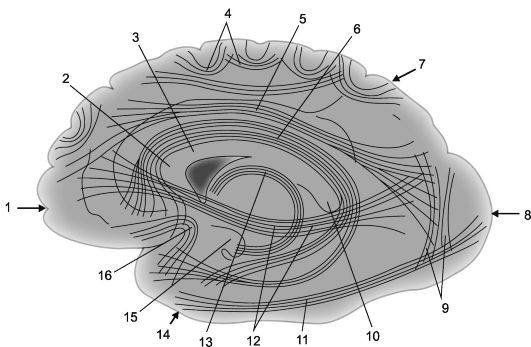
Živčana vlakna koja povezuju dijelove kore jedne hemisfere nazivaju se asocijativni. Što su ta vlakna i veze koje tvore kraća, to su površnija; duže asocijativne veze, smještene dublje, povezuju relativno udaljene dijelove kore velikog mozga (sl. 14.2 i 14.3).
Vlakna koja povezuju moždane hemisfere i stoga imaju zajedničku transverzalnu orijentaciju nazivaju se komisura, ili spavanje. Komisuralna vlakna povezuju identične dijelove moždanih hemisfera, stvarajući mogućnost kombiniranja njihovih funkcija. Formiraju se tri šiljka veliki mozak: najmasovniji od njih - Corpus callosum (Corpus callosum), osim toga, komisuralna vlakna čine prednja komisura, smješten ispod kljuna corpus callosuma (rostrum corporis collosum) i povezujući obje olfaktorne regije, kao i komisura svoda (commissura fornicis), ili hipokampalna komisura koju tvore vlakna koja povezuju strukture amonovih rogova obiju hemisfera.
U prednjem dijelu corpus callosum nalaze se vlakna koja povezuju frontalne režnjeve, zatim vlakna koja povezuju parijetalne i temporalne režnjeve, stražnji dio corpus callosum povezuje okcipitalne režnjeve mozga. Prednja komisura i komisura forniksa uglavnom spajaju dijelove drevne i stare kore obiju hemisfera; prednja komisura, osim toga, osigurava vezu između njihovih srednjih i donjih temporalnih vijuga.
14.4. Njušni sustav
U procesu filogeneze, razvoj velikog mozga povezan je s formiranjem olfaktornog sustava, čije funkcije pridonose očuvanju vitalnosti životinja i nisu od male važnosti za ljudski život.
Riža. 14.2.Asocijativne kortikalno-kortikalne veze u moždanim hemisferama [prema V.P. Vorobjov].
1 - frontalni režanj; 2 - koljeno corpus callosum; 3 - corpus callosum; 4 - lučna vlakna; 5 - gornja uzdužna greda; 6 - cingularni gyrus; 7 - parijetalni režanj, 8 - okcipitalni režanj; 9 - okomiti snopovi Wernickea; 10 - valjak corpus callosum;
11 - donja uzdužna greda; 12 - subkauzalni snop (frontalno-okcipitalni donji snop); 13 - svod; 14 - temporalni režanj; 15 - kuka gyrusa hipokampusa; 16 - snopovi kuka (fasciculus uncinatus).
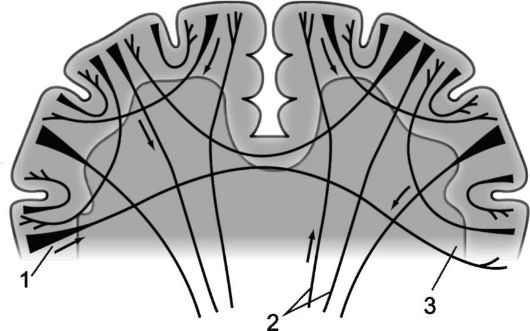
Riža. 14.3.Mijeloarhitektonika hemisfera velikog mozga.
1 - projekcijska vlakna; 2 - komisuralna vlakna; 3 - asocijativna vlakna.
14.4.1. Građa olfaktornog sustava
U sluznici su smještena tijela prvih neurona njušnog sustava nos, uglavnom gornji dio nosne pregrade i gornji nosni hodnik. Njušne stanice su bipolarne. Njihovi dendriti izlaze na površinu sluznice i tu završavaju specifičnim receptorima, te aksoni su grupirani u tzv mirisne niti (filiolfactorii),čiji je broj sa svake strane oko dvadeset. Takav snop njušnih niti i čini I kranijalni ili olfaktorni živac(Slika 14.4). Ove niti prolaze u prednju (olfaktornu, mirisnu) lubanjsku jamu kroz etmoidnu kost i završavaju kod stanice koje se nalaze ovdje mirisne lukovice. Olfaktorni bulbusi i proksimalni olfaktorni putevi zapravo su posljedica izbočina tvari velikog mozga nastalih u procesu ontogeneze i predstavljaju njemu srodne strukture.
Olfaktorne lukovice sadrže stanice koje su tijela drugih neurona. olfaktorni put, čiji aksoni nastaju olfaktorni traktovi (trakti olfactorii), smješten ispod olfaktornih utora, bočno od izravnih vijuga smještenih na bazalnoj površini frontalni režnjevi. Njušni putevi su usmjereni prema natrag na subkortikalne olfaktorne centre. Približavajući se prednjoj perforiranoj ploči, vlakna olfaktornog trakta podijeljena su u medijalne i lateralne snopove, tvoreći olfaktorni trokut sa svake strane. Kasnije su ova vlakna prikladna tijelima trećih neurona olfaktornog analizatora, koji se nalaze
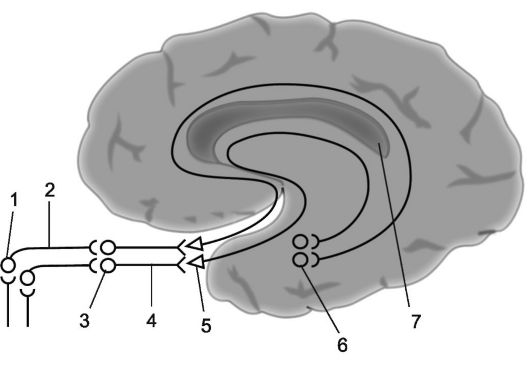
Riža. 14.4.Olfaktorni analizator.
1 - mirisne stanice; 2 - olfaktorne niti (ukupno čine olfaktorne živce); 3 - mirisne žarulje; 4 - olfaktorni putevi; 5 - mirisni trokuti; 6 - parahipokampalni girus; 7 - zona projekcije olfaktornog analizatora (pojednostavljeni dijagram).
u perialmondastom i subkalosalnom području, u jezgrama prozirnog septuma, smještenog ispred prednje komisure. Prednja komisura povezuje obje mirisne regije i također osigurava njihovu vezu s limbičkim sustavom mozga. Dio aksona trećih neurona olfaktornog analizatora, prolazeći kroz prednju komisuru mozga, križa se.
Aksoni trećih neurona olfaktorni analizator, smješten u subkortikalnim olfaktornim centrima, napreduje prema filogenetski stara kora mediobazalna površina temporalnog režnja (do piriformnog i parahipokampalnog girusa i do kuke), gdje se nalazi projekcijska olfaktorna zona, ili kortikalni kraj olfaktornog analizatora (polje 28, po Brodmannu).
Olfaktivni sustav je tako jedini osjetilni sustav u kojem specifični impulsi zaobilaze talamus na putu od receptora do korteksa. Međutim, Njušni sustav ima posebno izražene veze s limbičkim strukturama mozga, a informacije primljene preko njega imaju značajan utjecaj na stanje emocionalne sfere i funkcije autonomnog živčanog sustava. Mirisi mogu biti ugodni i neugodni, utječu na apetit, raspoloženje, mogu izazvati razne vegetativne reakcije, posebice mučninu, povraćanje.
14.4.2. Ispitivanje osjeta mirisa i značenja njegovih poremećaja za topikalnu dijagnostiku
Pri ispitivanju stanja njuha potrebno je ustanoviti osjeća li bolesnik mirise, jesu li ti osjeti obostrano jednaki, razlikuje li bolesnik prirodu osjetnih mirisa, ima li olfaktorne halucinacije - paroksizmalne osjete mirisa. koji su odsutni u okoliš.
Za proučavanje osjeta mirisa koriste se mirisne tvari čiji miris nije oštar (oštar miris može izazvati iritaciju receptora trigeminalnog živca koji se nalazi u nosnoj sluznici) i poznat je pacijentu (inače je teško prepoznati izopačenost mirisa). Njuh se provjerava sa svake strane posebno, dok druga nosnica mora biti zatvorena. Možete koristiti posebno pripremljene setove slabih otopina mirisnih tvari (metvica, katran, kamfor itd.), U praktičnom radu također se mogu koristiti improvizirana sredstva ( raženi kruh, sapun, banana itd.).
Smanjeni osjet mirisa - hiposmija, nedostatak mirisa - anosmija, pojačan osjet mirisa - hiperosmija, izopačenost mirisa disosmija, osjet mirisa u odsutnosti podražaja - parozmija, subjektivni osjećaj loš miris, koji zapravo postoji i uzrokovan je organskom patologijom u nazofarinksu - kakosmiya, mirisi koji zapravo ne postoje, koje bolesnik osjeća paroksizmalno - olfaktorne halucinacije - češće su olfaktorna aura epilepsije temporalnog režnja, koja može biti posljedica raznih razloga, posebice tumora temporalnog režnja.
Obostrana hipozmija ili anosmija najčešće je posljedica oštećenja nosne sluznice uslijed akutnog katara, gripe, alergijskog rinitisa, atrofije sluznice
nosa zbog kroničnog rinitisa i dugotrajnu upotrebu vazokonstriktorske kapi za nos. Kronični rinitis s atrofijom nosne sluznice (atrofični rinitis), Sjögrenova bolest osuđuje osobu na trajnu anosmiju. Bilateralna hiposmija može biti uzrokovana hipotireozom, dijabetes melitusom, hipogonadizmom, zatajenjem bubrega, produljenim kontaktom s teškim metalima, formaldehidom itd.
Međutim, jednostrana hiposmija ili anosmija često je posljedica intrakranijalnog tumora, češće meningeoma prednje lubanjske (olfaktorne) jame, što čini do 10% intrakranijalnih meningeoma, kao i neke glijalne tumore frontalnog režnja. Poremećaji njuha nastaju kao posljedica kompresije olfaktornog trakta na strani patološkog žarišta i mogu biti jedini žarišni simptom bolesti određeno vrijeme. Tumori se mogu vizualizirati CT ili MRI skeniranjem. Kako se meningiomi olfaktorne jame povećavaju, u pravilu se razvijaju mentalni poremećaji karakterističan za frontalni sindrom (vidi poglavlje 15).
Jednostrano oštećenje dijelova olfaktornog analizatora koji se nalaze iznad njegovih supkortikalnih središta, zbog nepotpune presijecanja putova na razini prednje cerebralne komisure, obično ne dovodi do značajnog smanjenja osjeta mirisa. Iritacija patološkim procesom korteksa mediobazalnih dijelova temporalnog režnja, prvenstveno parahipokampalnog girusa i njegove kuke, može izazvati paroksizmalnu pojavu. mirisne halucinacije. Bolesnik odjednom bez ikakvog razloga počne osjećati miris, često neugodne prirode (miris spaljenog, pokvarenog, pokvarenog, spaljenog itd.). Olfaktorne halucinacije u prisutnosti epileptogenog žarišta u mediobazalnim regijama temporalnog režnja mozga može biti manifestacija aure epileptičkog napadaja. Poraz proksimalnog dijela, posebno kortikalnog kraja olfaktornog analizatora, može uzrokovati umjerenu bilateralnu (više na suprotnoj strani) hiposmiju i oslabljenu sposobnost prepoznavanja i razlikovanja mirisa (olfaktorna agnozija). Posljednji oblik olfaktornog poremećaja, koji se manifestira u starijoj dobi, najvjerojatnije je povezan s kršenjem funkcije korteksa zbog atrofičnih procesa u njegovoj projekcijskoj olfaktornoj zoni.
14.5. LIMBIČKO-RETIKULARNI KOMPLEKS
Godine 1878. P. Broca(Broca P., 1824.-1880.) pod nazivom "veliki rubni ili limbički režanj" (od lat. limbus - rub) ujedinio je hipokampus i cingulatni girus, međusobno povezani pomoću isthmusa cingulate gyrusa, koji se nalazi iznad grebena corpus callosum.
Godine 1937. D. Papets(Papez J.), na temelju eksperimentalnih podataka, iznio je argumentiranu primjedbu na prethodno postojeći koncept uključenosti mediobazalnih struktura moždanih hemisfera uglavnom u pružanje mirisa. On sugerirao da je glavni dio mediobazalnih dijelova cerebralne hemisfere, tada nazvan olfaktorni mozak (rhinencephalon), kojemu pripada limbički režanj, morfološka osnova živčani mehanizam afektivno ponašanje, te ih objedinili pod imenom"emocionalni krug" koji uključuje hipotalamus,
prednje jezgre talamusa, cingularni girus, hipokampus i njihove veze. Od tada te strukture fiziolozi također nazivaju oko Papeza.
koncept "visceralni mozak" predložio je P.D. McLean (1949), označavajući tako složenu anatomsko-fiziološku asocijaciju, koja se od 1952. godine naziva "limbički sustav". Kasnije se pokazalo da je limbički sustav uključen u obavljanje različitih funkcija, a sada se većina, uključujući cingulatni i hipokampalni (parahipokampalni) girus, obično spaja u limbičku regiju, koja ima brojne veze sa strukturama retikularna formacija, nadoknađujući je limbičko-retikularni kompleks, osiguravajući širok raspon fizioloških i psiholoških procesa.
Trenutno do limbički režanj uobičajeno je pripisati elemente starog korteksa (arhiokorteks), koji pokrivaju zupčani girus i hipokampalni girus; drevni korteks (paleokorteks) prednjeg hipokampusa; kao i srednji, ili srednji, korteks (mezokorteks) cingularnog girusa. Termin "limbički sustav" uključuje komponente limbičkog režnja i srodne strukture - entorinalne (zauzimaju najviše parahipokampalnog girusa) i septalne regije, kao i kompleksa amigdale i mastoidnog tijela (Duus P., 1995.).
Mastoidno tijelo povezuje strukture ovog sustava sa srednjim mozgom i sa retikularna formacija. Impulsi koji potječu iz limbičkog sustava mogu se prenijeti kroz prednju jezgru talamusa do cingularnog girusa i do neokorteksa duž putova koje formiraju asocijativna vlakna. Impulsi koji potječu iz hipotalamusa mogu doći do orbitofrontalnog korteksa i medijalne dorzalne jezgre talamusa.
Brojne izravne i povratne veze osiguravaju međusobnu povezanost i međuovisnost limbičkih struktura i mnogih tvorevina diencefalona i oralnih dijelova moždanog debla (nespecifične jezgre talamusa, hipotalamusa, putamena, frenuluma, retikularne formacije moždanog debla), kao i sa subkortikalnim jezgre (pallidus, putamen, caudatus nucleus ) i s neokorteksom moždanih hemisfera, prvenstveno s korteksom temporalnog i frontalnog režnja.
Unatoč filogenetskim, morfološkim i citoarhitektonskim razlikama, mnoge od navedenih struktura (limbička regija, središnje i medijalne strukture talamusa, hipotalamus, retikularna formacija moždanog debla) obično se ubrajaju u tzv. limbičko-retikularni kompleks, koja djeluje kao zona integracije mnogih funkcija, osiguravajući organizaciju polimodalnih, cjelovitih reakcija organizma na različite utjecaje, što je posebno izraženo u stresnim situacijama.
Strukture limbičko-retikularnog kompleksa imaju veliki broj ulazi i izlazi kroz koje začarani krugovi od brojnih aferentnih i eferentnih veza koje osiguravaju kombinirano funkcioniranje formacija uključenih u ovaj kompleks i njihovu interakciju sa svim dijelovima mozga, uključujući moždanu koru.
U strukturama limbičko-retikularnog kompleksa postoji konvergencija osjetljivih impulsa koji se javljaju u intero- i eksteroreceptorima, uključujući receptorska polja osjetilnih organa. Na temelju toga, u limbičko-retikularnom kompleksu, primarna sinteza informacija o stanju unutarnje okoline tijela, kao io čimbenicima vanjske okoline koji utječu na tijelo, te se formiraju elementarne potrebe, biološke motivacije i popratne emocije.
Limbičko-retikularni kompleks određuje stanje emocionalne sfere, sudjeluje u regulaciji vegetativno-visceralnih odnosa usmjerenih na održavanje relativne konstantnosti unutarnjeg okoliša (homeostaze), kao i opskrbu energijom i korelaciju motoričkih akata. O njegovu stanju ovisi stupanj svijesti, mogućnost automatiziranih pokreta, aktivnost motoričkih i mentalnih funkcija, govor, pažnja, sposobnost orijentacije, pamćenje, promjena budnosti i spavanja.
Oštećenje struktura limbičko-retikularnog kompleksa može biti popraćeno različitim klinički simptomi: izražene promjene u emocionalnoj sferi trajne i paroksizmalne prirode, anoreksija ili bulimija, seksualni poremećaji, oštećenje pamćenja, posebno znakovi Korsakoffovog sindroma, kod kojih bolesnik gubi sposobnost pamćenja aktualnih događaja (zadržava aktualne događaje u sjećanju ne dulje od 2 minute), autonomno-endokrini poremećaji, poremećaji spavanja, psihosenzorni poremećaji u obliku iluzija i halucinacija, promjene svijesti, kliničke manifestacije akinetičkog mutizma, epileptični napadaji.
Do danas je proveden veliki broj studija o proučavanju morfologije, anatomskih odnosa, funkcije limbičke regije i drugih struktura uključenih u limbičko-retikularni kompleks, međutim, fiziologija i značajke kliničke slike njegove lezije danas još uvelike treba razjasniti. Većina podataka o njegovoj funkciji, posebno funkcije parahipokampalne regije, dobivenih u pokusima na životinjama metode iritacije, ekstirpacije ili stereotaksije. Dobiven na ovaj način rezultati zahtijevaju oprez pri ekstrapolaciji na ljude. Od posebne su važnosti klinička promatranja bolesnika s lezijama mediobazalnih dijelova moždane hemisfere.
U 50-60-im godinama XX stoljeća. tijekom razvoja psihokirurgije pojavili su se izvještaji o liječenju bolesnika s neizlječivim mentalni poremećaji i kroničnog bolnog sindroma bilateralnom cingulotomijom (disekcija cingularnog girusa), a obično je zabilježena regresija anksioznosti, opsesivnih stanja, psihomotorne agitacije, bolni sindromi, što je prepoznato kao dokaz uključenosti cingularnog gyrusa u formiranje emocija i boli. Istodobno, bicingulotomija je dovela do dubokih poremećaja osobnosti, do dezorijentacije, smanjenja kritičnosti vlastitog stanja i euforije.
Analiza 80 verificiranih kliničkih slučajeva lezija hipokampusa na temelju Neurokirurškog instituta Ruske akademije medicinskih znanosti dana je u monografiji N.N. Bragina (1974). Autor dolazi do zaključka da temporalni mediobazalni sindrom uključuje viscerovegetativne, motoričke i mentalne poremećaje, koji se obično manifestiraju u kompleksu. Sva raznolikost kliničkih manifestacija N.N. Bragin svodi na dvije glavne multifaktorijalne varijante patologije s prevlašću "iritativnih" i "inhibicijskih" fenomena.
Prvi od njih uključuje emocionalni poremećaji popraćeno motoričkom anksioznošću (povećana razdražljivost, govorljivost, nemir, osjećaj unutarnje tjeskobe), paroksizma straha, vitalne tjeskobe, raznih viscerovegetativnih poremećaja (promjene u pulsu, disanju, gastrointestinalni poremećaji, groznica, pojačano znojenje itd.). U ovih bolesnika, na pozadini stalnog motoričkog nemira, često su se javljali napadi motoričke ekscitacije.
niya. EEG ove skupine bolesnika karakteriziran je blagim cerebralnim promjenama prema integraciji (ubrzani i zaoštreni alfa ritam, difuzne beta oscilacije). Ponovljeni aferentni podražaji izazvali su jasne EEG odgovore, koji, za razliku od normalnih, nisu blijedili kako su podražaji opetovano prezentirani.
Drugu ("inhibitornu") varijantu mediobazalnog sindroma karakteriziraju emocionalni poremećaji u obliku depresije s motoričkom retardacijom (depresivno raspoloženje, osiromašenje i usporavanje tempa). mentalni procesi, promjene motiliteta, podsjećaju na tip akinetičko-rigidnog sindroma. Viscerovegetativni paroksizmi zabilježeni u prvoj skupini manje su karakteristični. EEG bolesnika u ovoj skupini karakteriziraju cerebralne promjene, koje se očituju u prevladavanju sporih oblika aktivnosti (nepravilan, spor alfa ritam, skupine theta oscilacija, difuzni delta valovi). Oštar pad EEG reaktivnosti privukao je pozornost.
Između ove dvije ekstremne opcije postojale su srednje s prijelaznim i mješovitim kombinacijama. pojedinačne simptome. Dakle, neke od njih karakteriziraju relativno slabi znakovi agitirane depresije s pojačanim motorna aktivnost i umor, s prevlašću senestopatskih osjeta, sumnjičavost, dosežući kod nekih pacijenata do paranoidnih stanja, hipohondrijski delirij. Druga srednja skupina odlikovala se ekstremnim intenzitetom depresivnih simptoma na pozadini pacijentove ukočenosti.
Ovi podaci omogućuju nam govoriti o dvostrukom (aktivirajućem i inhibitornom) utjecaju hipokampusa i drugih struktura limbičke regije na reakcije ponašanja, emocije, mentalni status i bioelektričnu aktivnost korteksa. Trenutno složeno klinički sindromi ovaj tip ne treba smatrati primarnim žarišnim. Umjesto toga, treba ih razmatrati u svjetlu ideja o višerazinskom sustavu organizacije moždane aktivnosti.
S.B. Buklina (1997) navodi podatke iz ankete 41 bolesnika s arteriovenskim malformacijama u području cingulate gyrus. Prije operacije 38 bolesnika imalo je u prvom planu poremećaje pamćenja, a pet ih je imalo znakove Korsakoffovog sindroma, kod tri bolesnika Korsakoffov sindrom nastao je nakon operacije, dok je težina porasta poremećaja pamćenja korelirala sa stupnjem destrukcije cingulate gyrusa. samom, kao i zahvaćanjem u patološki proces susjednih struktura corpus callosuma, dok amnezijski sindrom nije ovisio o strani lokalizacije malformacije i njezinoj lokalizaciji duž duljine cingulate gyrusa.
Glavna obilježja identificiranih amnestičkih sindroma bili su poremećaji u reprodukciji slušno-govornih podražaja, poremećaji selektivnosti tragova u obliku inkluzija i kontaminacija te nemogućnost zadržavanja značenja u prijenosu priče. Kod većine bolesnika smanjena je kritičnost procjene njihovog stanja. Autor je primijetio sličnost ovih poremećaja s amnestičkim defektima u bolesnika s frontalnim lezijama, što se može objasniti prisutnošću veza između cingulate gyrusa i frontalnog režnja.
Više rašireni patološki procesi u limbičkoj regiji uzrokuju izražene poremećaje vegetativno-visceralnih funkcija.
Corpus callosum(Corpus callosum)- najveća komisura između moždanih hemisfera. Njegovi prednji dijelovi, posebno koljeno corpus callosum
tijelo (genu corporis callosi), spojite frontalne režnjeve, srednje dijelove - deblo corpus callosum (truncus corporis callosi)- osiguravaju komunikaciju između temporalnih i parijetalnih dijelova hemisfera, stražnjih odjela, posebno grebena corpus callosum (splenium corporis callosi), spojite okcipitalne režnjeve.
Lezije corpus callosuma obično su popraćene poremećajima mentalnog stanja bolesnika. Uništavanje njegovog prednjeg dijela dovodi do razvoja "frontalne psihe" (aspontanost, kršenje akcijskog plana, ponašanje, kritičnost, karakteristično za frontalni kalozni sindrom - akinezija, amimija, aspontanost, astazija-abazija, apraksija, refleksi hvatanja, demencija). Prekid veza između parijetalnih režnjeva dovodi do perverzije razumijevanje "planovi tijela" I pojava apraksije uglavnom u lijevoj ruci. Disocijacija temporalnih režnjeva može manifestirati kršenje percepcije vanjskog okruženja, gubitak ispravne orijentacije u njemu (amnestički poremećaji, konfabulacije, sindrom već viđenog i tako dalje.). Patološka žarišta u stražnjim dijelovima corpus callosum često karakteriziraju znakovi vizualne agnozije.
14.6. ARHITEKTONIKA MOŽDANE KORE
Struktura kore velikog mozga je heterogena. Manje složene strukture, rano se pojavljuju u procesu filogeneze prastara kora (arhiokorteks) i stara kora (paleokorteks), srodni uglavnom do limbičkog režnja mozak. Veći dio kore velikog mozga (95,6%), zbog kasnijeg filogenetskog oblikovanja, tzv. nova kora (neokorteks) i ima mnogo složeniju višeslojnu strukturu, ali i heterogenu u svojim različitim zonama.
Zbog arhitektonika korteksa je u određenoj vezi s njegovom funkcijom, mnogo je pažnje posvećeno njegovom proučavanju. Jedan od utemeljitelja doktrine citoarhitektonike korteksa bio je V.A. Betz (1834-1894), prvi je opisao velike piramidalne stanice 1874. motorička zona korteks (Betzove stanice) i odredio principe podjele moždane kore na glavna područja. U budućnosti, veliki doprinos razvoju teorije o strukturi korteksa dali su mnogi istraživači - A. Campbell (A. Cambell), E. Smith (E. Smith), K. Brodmann (K. Brodmann ), Oscar Vogt i Cecilia Vogt (O. Vogt , S. Vogt). Velike zasluge u proučavanju arhitektonike korteksa pripadaju osoblju Instituta za mozak Akademije medicinskih znanosti (S.A. Sarkisov, N.I. Filimonov, E.P. Kononova i dr.).
Glavni tip strukture nove kore (Slika 14.5), s kojim se uspoređuju svi njegovi odsječci – kora koja se sastoji od 6 slojeva (homotipska kora, prema Brodmanu).
Sloj I - molekularni, ili zonalni, najpovršniji, siromašan stanicama, njegova vlakna imaju smjer, uglavnom paralelan s površinom korteksa.
II sloj - vanjski zrnati. Sastoji se od velikog broja gusto raspoređenih sitnih zrnaca nervne ćelije.
III sloj - male i srednje piramide, najšire. Sastoji se od piramidalnih stanica, čije veličine nisu iste, što omogućuje podjelu ovog sloja na podslojeve u većini kortikalnih polja.
IV sloj - unutarnji zrnati. Sastoji se od gusto poredanih malih stanica-zrna okruglog i uglatog oblika. Ovaj sloj je najvarijabilniji
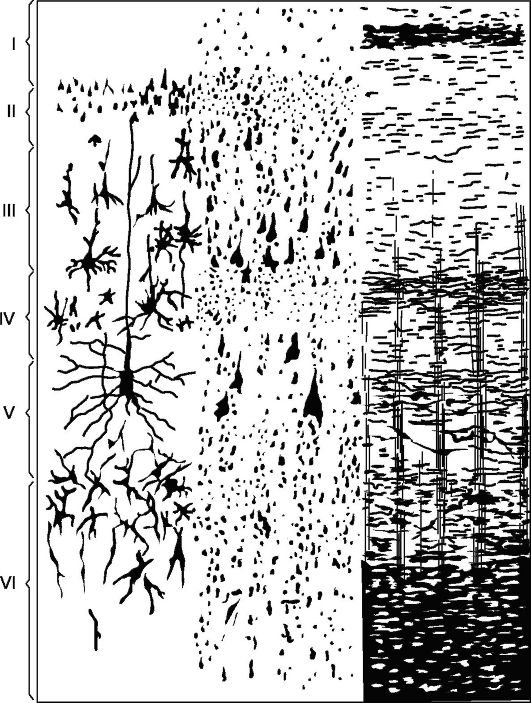
Riža. 14.5.Citoarhitektonika i mijeloarhitektonika motorne zone kore velikog mozga.
Lijevo: I - molekularni sloj; II - vanjski zrnati sloj; III - sloj malih i srednjih piramida; IV - unutarnji zrnati sloj; V - sloj velikih piramida; VI - sloj polimorfnih stanica; desno - elementi mijeloarhitektonike.
u nekim poljima (npr. polje 17) podijeljen je u podslojeve, na nekim mjestima naglo postaje tanji i čak potpuno nestaje.
V sloj - velike piramide, ili ganglijske. Sadrži velike piramidalne stanice. U nekim područjima mozga sloj je podijeljen na podslojeve, u motoričkoj zoni sastoji se od tri podsloja, od kojih se u sredini nalaze Betzove divovske piramidalne stanice, koje dosežu promjer od 120 mikrona.
VI sloj - polimorfne stanice, ili multiformne. Sastoji se uglavnom od trokutastih stanica vretenastog oblika.
Građa moždane kore ima veliki broj varijacija zbog promjene debljine pojedinih slojeva, stanjivanja ili nestajanja ili,
naprotiv, zgušnjavanje i podjela na podslojeve nekih od njih (heterotipne zone, po Brodmanu).
Kora svake cerebralne hemisfere podijeljena je u nekoliko regija: okcipitalnu, gornju i donju parijetalnu, postcentralnu, središnju vijugu, precentralnu, frontalnu, temporalnu, limbičku, inzularnu. Svaki od njih u skladu sa karakteristikama podijeljeno u nekoliko polja, osim toga, svako polje ima svoju konvencionalnu oznaku reda (Sl. 14.6).
Proučavanje arhitektonike moždane kore, uz fiziološka, uključujući elektrofiziološka istraživanja i klinička opažanja, uvelike je pridonijelo rješavanju problema raspodjele funkcija u korteksu.
14.7. PROJEKCIJSKA I ASOCIJACIJSKA POLJA CORTUSA
U procesu razvoja doktrine o ulozi cerebralnog korteksa i njegovih pojedinih dijelova u obavljanju određenih funkcija, postojala su različita, ponekad suprotna, gledišta. Dakle, postojalo je mišljenje o strogo lokalnoj zastupljenosti u kori velikog mozga svih ljudskih sposobnosti i funkcija, do najsloženijih, mentalnih. (lokalizacijski, psihomorfologizam). Njemu se suprotstavilo drugo mišljenje o apsolutnoj funkcionalnoj jednakosti svih dijelova moždane kore. (ekvipotencijalizam).
Važan doprinos teoriji lokalizacije funkcija u cerebralnom korteksu dao je I.P. Pavlov (1848-1936). Izdvojio je projekcijske zone korteksa (kortikalni krajevi analizatora određene vrste osjetljivost) i asocijativne zone smještene između njih, proučavali su procese inhibicije i ekscitacije u mozgu, njihov utjecaj na funkcionalno stanje moždana kora. Podjela kore na projekcijske i asocijativne zone doprinosi razumijevanju organizacije rada moždane kore i opravdava se u rješavanju praktičnih zadataka, posebice u lokalnoj dijagnostici.
projekcijske zone pružaju uglavnom jednostavne specifične fiziološke radnje, prvenstveno percepciju osjeta određenog modaliteta. Projekcijski putovi koji im se približavaju povezuju te zone s receptorskim teritorijima na periferiji koji su s njima u funkcionalnoj korespondenciji. Primjeri projekcijskih kortikalnih zona su područje stražnjeg središnjeg girusa već opisano u prethodnim poglavljima (zona opći tipovi osjetljivost) ili područje trnnog žlijeba koji se nalazi na medijalnoj strani okcipitalnog režnja (projekcijska vidna zona).
Zone udruživanja korteks nema izravne veze s periferijom. Nalaze se između projekcijskih zona i imaju brojne asocijativne veze s tim projekcijskim zonama i s drugim asocijativnim zonama. Funkcija asocijacijskih zona je provođenje viša analiza te sinteza mnogih elementarnih i složenijih komponenti. Ovdje, u biti, postoji razumijevanje informacija koje ulaze u mozak, formiranje ideja i koncepata.
G.I. Polyakov je 1969. godine, na temelju usporedbe arhitektonike kore velikog mozga čovjeka i nekih životinja, utvrdio da asocij.
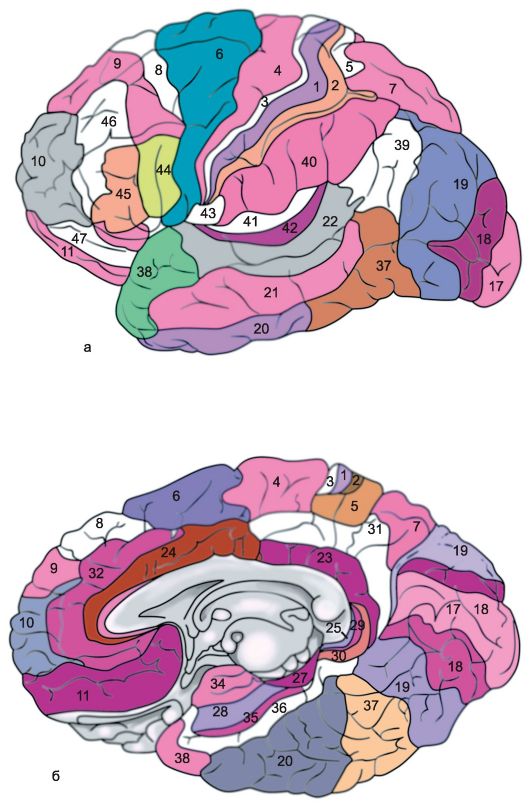
Riža. 14.6.Arhitektonska polja kore velikog mozga [prema Brodmanu]. a - vanjska površina; b - medijalna površina.
zone u ljudskom cerebralnom korteksu su 50%, u korteksu viših (humanoidnih) majmuna - 20%, u nižim majmunima ta je brojka 10% (slika 14.7). Među asocijacijskim područjima korteksa ljudskog mozga, isti je autor predložio izolaciju sekundarna i tercijarna polja. Sekundarna asocijativna polja su uz projekcijska. Provode analizu i sintezu elementarnih osjeta koji još uvijek zadržavaju specifičnu orijentaciju.
Tercijarna asocijativna polja nalaze se uglavnom između sekundarnih i preklapaju zone susjednih teritorija. Oni su prvenstveno povezani s analitičkom aktivnošću korteksa, osiguravajući najviše mentalne funkcije svojstvene čovjeku u njihovim najsloženijim intelektualnim i govornim manifestacijama. Funkcionalna zrelost tercijarnog as-
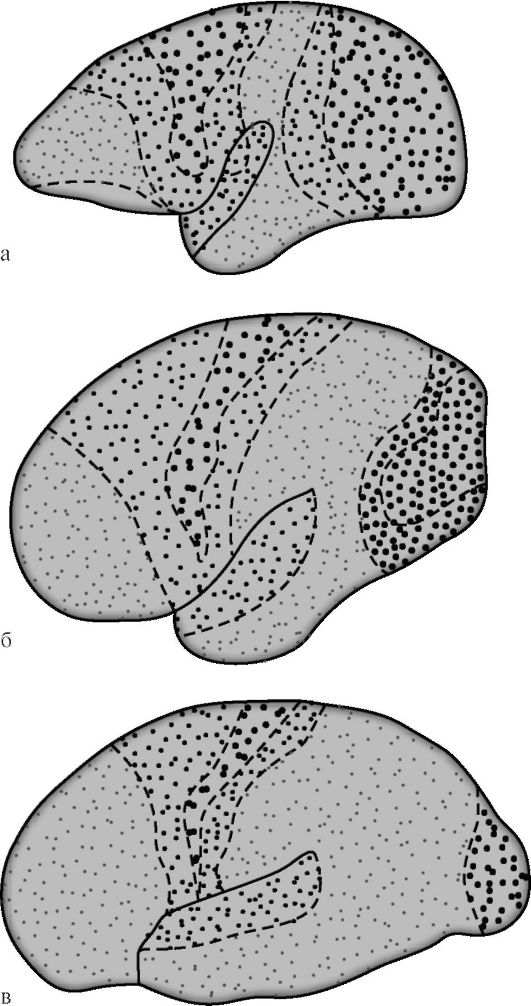
Riža. 14.7. Diferencijacija projekcijskih i asocijativnih područja cerebralnog korteksa tijekom evolucije primata [prema G.I. Polyakov]. a - mozak nižeg majmuna; b - mozak višeg majmuna; c - ljudski mozak. Velike točke označavaju zone projekcije, male točke - asocijativne. Kod nižih majmuna, asocijativne zone zauzimaju 10% površine korteksa, kod viših - 20%, kod ljudi - 50%.
socijalnih polja moždane kore javlja se najkasnije i to samo u povoljnom društvenom okruženju. Za razliku od ostalih kortikalnih polja, tercijarna polja desne i lijeve hemisfere karakterizira izražena funkcionalna asimetrija.
14.8. AKTUELNA DIJAGNOSTIKA LEZIJA MOŽDANE KORE
14.8.1. Manifestacije oštećenja projekcijskih zona cerebralnog korteksa
U korteksu svake cerebralne hemisfere, iza središnjeg girusa, nalazi se 6 projekcijskih zona.
1. U prednjem dijelu parijetalnog režnja, u području stražnjeg središnjeg girusa (citoarhitektonska polja 1, 2, 3) nalazi se projekcijska zona općih tipova osjetljivosti(Slika 14.4). Ovdje smještena područja korteksa primaju osjetljive impulse koji dolaze duž projekcijskih putova općih tipova osjetljivosti iz receptorskog aparata suprotne polovice tijela. Što je veća površina ove projekcijske zone korteksa, to niže smješteni dijelovi suprotne polovice tijela imaju projekcijske veze. Dijelovi tijela s ekstenzivnom recepcijom (jezik, palmarna površina šake) odgovaraju neadekvatno velikim dijelovima područja projekcijskih zona, dok ostali dijelovi tijela (proksimalni udovi, torzo) imaju malu površinu kortikalne kore. reprezentacija.
Iritacija patološkim procesom kortikalne zone općih tipova osjetljivosti dovodi do napada parestezije u dijelovima tijela koji odgovaraju nadraženim područjima cerebralnog korteksa (osjetljivi Jacksonov napadaj), koji se može pretvoriti u sekundarni generalizirani paroksizam. Poraz kortikalnog kraja analizatora općih tipova osjetljivosti može uzrokovati razvoj hipalgezije ili anestezije u odgovarajućoj zoni suprotne polovice tijela, dok mjesto hipestezije ili anestezije može biti vertikalno cirkulacijsko ili radikularno- segmentni tip. U prvom slučaju, poremećaj osjetljivosti manifestira se na strani suprotnoj od patološkog žarišta u području usana, palca ili u distalnom dijelu ekstremiteta s kružnim rubom, ponekad poput čarape ili rukavice. U drugom slučaju, zona poremećaja osjetljivosti ima oblik trake i nalazi se duž unutarnjeg ili vanjskog ruba ruke ili noge; to se objašnjava činjenicom da je unutarnja strana udova predstavljena u prednjoj, a vanjska strana - u stražnjim dijelovima projekcijske zone analizatora općih tipova osjetljivosti.
2. Zona vizualne projekcije nalazi se u korteksu medijalne plohe okcipitalnog režnja u predjelu trnnog žlijeba (polje 17). U ovom polju dolazi do raslojavanja IV (unutarnjeg granularnog) sloja korteksa sa snopom mijelinskih vlakana u dva podsloja. Odvojeni dijelovi polja 17 primaju impulse iz određenih dijelova istoimenih polovica mrežnice oba oka; dok impulsi koji dolaze iz donjih dijelova homonimnih polovica mrežnice dopiru
kora donje usne utora ostruge i impulsi koji dolaze iz gornji dijelovi retine, šalju se u korteks njezine gornje usne.
Poraz patološkog procesa zone vizualne projekcije dovodi do pojave na suprotnoj strani kvadranta ili potpune homonimne hemianopije na strani suprotnoj od patološkog fokusa. Bilateralna lezija kortikalnih polja 17 ili projekcijskih polja koja idu do njih vizualni putevi može dovesti do potpunog sljepila. Iritacija korteksa vidne projekcijske zone može izazvati pojavu vidnih halucinacija u obliku fotopsija u odgovarajućim dijelovima suprotnih polovica vidnih polja.
3. Područje projekcije sluha nalazi se u korteksu Heschlovih vijuga na donja usna bočna (Silvijeva) brazda (polja 41 i 42), koji su zapravo dio gornjeg temporalnog vijuga. Iritacija ove zone korteksa može uzrokovati pojavu slušnih halucinacija (napadaji osjećaja buke, zvonjenja, zviždanja, zujanja itd.). Uništenje slušne projekcijske zone s jedne strane može uzrokovati određeni gubitak sluha na oba uha, u većoj mjeri na suprotnoj strani u odnosu na patološko žarište.
4 i 5. Zone mirisne i okusne projekcije su na medijalnoj površini zasvođenog vijuga (limbička regija) mozga. Prvi od njih nalazi se u parahipokampalnom girusu (polje 28). Zona projekcije okusa obično je lokalizirana u korteksu operkularnog područja (polje 43). Iritacija projekcijskih zona mirisa i okusa može uzrokovati njihovu perverziju ili dovesti do razvoja odgovarajućih mirisnih i okusnih halucinacija. Jednostrani gubitak funkcije projekcijskih zona mirisa i okusa može uzrokovati lagano smanjenje mirisa i okusa, odnosno, s obje strane. Bilateralno uništenje kortikalnih krajeva istih analizatora očituje se odsutnošću mirisa i okusa s obje strane.
6. Zona vestibularne projekcije. Njegova lokalizacija nije navedena. Istodobno, poznato je da vestibularni aparat ima brojne anatomske i funkcionalne veze. Moguće je da lokalizacija zastupljenosti vestibularnog sustava u korteksu još nije razjašnjena jer je polifokalna. N.S. Blagoveshchenskaya (1981) vjeruje da su u cerebralnom korteksu zone vestibularne projekcije predstavljene s nekoliko anatomskih i funkcionalnih kompleksa koji međusobno djeluju, a koji se nalaze u polju 8, na spoju frontalnog, temporalnog i tjemeni režanj a u zoni središnjih vijuga pretpostavlja se da svaki od tih odjeljaka korteksa obavlja svoje vlastite funkcije. Polje 8 proizvoljno je središte pogleda, njegova iritacija uzrokuje skretanje pogleda u smjeru suprotnom od patološkog žarišta, promjene u ritmu i prirodi eksperimentalnog nistagmusa, osobito ubrzo nakon epileptičkog napadaja. U korteksu temporalnog režnja postoje strukture, čija iritacija uzrokuje vrtoglavicu, što se posebno očituje u epilepsiji temporalnog režnja; poraz područja zastupljenosti vestibularnih struktura u korteksu središnjih vijuga utječe na stanje tonusa poprečno-prugastih mišića. Klinička opažanja sugeriraju da nuklearno-kortikalni vestibularni putovi čine djelomičnu križanje.
Treba naglasiti da znakovi iritacije navedenih projekcijskih zona mogu biti manifestacija aure odgovarajuće prirode epileptičnog napadaja.
I.P. Pavlov je smatrao mogućim uzeti u obzir koru precentralnog girusa, koji utječe na motoričke funkcije i mišićni tonus pretežno suprotne polovice tijela, s kojom je povezan prvenstveno kortikalno-nuklearnim i kortikalno-spinalnim (piramidalnim) putovima, kao projekcijska zona tzv motorički analizator. Ova zona zauzima prije svega polje 4, na koje se u obrnutom obliku projicira suprotna polovica tijela. Ovo polje sadrži glavninu ogromnih piramidalnih stanica (Betzovih stanica), čiji aksoni čine 2-2,5% svih vlakana piramidnog puta, kao i srednje i male piramidalne stanice, koje zajedno s aksonima istih stanice koje se nalaze u susjedstvu polja 4, šireg polja 6, uključene su u provedbu monosinaptičkih i polisinaptičkih kortikalno-mišićnih veza. Monosinaptičke veze omogućuju uglavnom brze i precizne ciljane radnje, ovisno o kontrakcijama pojedinca isprugana mišići.
Oštećenje donjih dijelova motoričke zone obično dovodi do razvoja na suprotnoj strani brahiofacijalni (lice ramena) sindrom ili lingvofaciobrahijalni sindrom, koji se često opažaju u bolesnika s oštećenom cerebralnom cirkulacijom u bazenu srednje moždane arterije, dok kombinirana pareza mišića lica, jezika i ruke, osobito ramena u središnjem tipu.
Iritacija korteksa motoričke zone (polja 4 i 6) dovodi do pojave konvulzija u mišićima ili mišićnim skupinama projiciranim na ovu zonu. Češće su to lokalne konvulzije tipa Jacksonove epilepsije, koje se mogu transformirati u sekundarni generalizirani epileptički napadaj.
14.8.2. Manifestacije oštećenja asocijativnih polja cerebralnog korteksa
Između projekcijskih zona kore su asocijacijska polja. Oni primaju impulse uglavnom iz stanica projekcijskih zona korteksa. U asocijativnim poljima postoji analiza i sinteza informacija koje su prošle primarna obrada u projekcijskim poljima. Asocijativne zone korteksa gornjeg parijetalnog lobula osiguravaju sintezu elementarnih osjeta, s tim u vezi, tako složene vrste osjetljivosti kao što su osjećaj lokalizacije, osjećaj težine, dvodimenzionalno-prostorni osjećaj, kao i složeni tu nastaju kinestetički osjeti.
U području interparijetalnog sulkusa nalazi se asocijativna zona koja omogućuje sintezu osjeta koji proizlaze iz dijelova vlastitog tijela. Oštećenje ove regije korteksa dovodi do autopagnozija, oni. na neprepoznavanje ili ignoriranje dijelova vlastitog tijela ili na pseudomelija osjećaj da imate višak ruke ili noge i anozognozija - nedostatak svijesti o tjelesnom nedostatku koji je nastao u vezi s bolešću (na primjer, paraliza ili pareza udova). Obično se sve vrste autopagnozije i anozognozije javljaju kada se patološki proces nalazi desno.
Poraz donjeg parijetalnog lobula može se očitovati poremećajem u sintezi elementarnih osjeta ili nemogućnošću usporedbe sintetiziranih složenih osjeta s "nekoć u percepciji sličnih
na isti način, na temelju rezultata kojih dolazi do prepoznavanja ”(V.M. Bekhterev). To se očituje kršenjem dvodimenzionalnog prostornog osjeta (grafoestezija) i trodimenzionalnog prostornog osjeta (stereognozija) - astereognoza.
U slučaju oštećenja premotornih zona frontalnog režnja (polja 6, 8, 44) obično dolazi do frontalne ataksije, u kojoj je poremećena sinteza aferentnih impulsa (kinestetička aferentacija), signalizirajući položaj dijelova tijela u prostoru koji promjene tijekom izvršenih pokreta.
U slučaju kršenja funkcije kore prednjeg dijela frontalnog režnja, koji ima veze sa suprotnom hemisferom malog mozga (fronto-mostno-cerebelarne veze), statokinetički poremećaji nastaju na suprotnoj strani patološkog žarišta. (frontalna ataksija). Osobito su izraženi poremećaji kasno razvijenih oblika statokinetike - uspravno stajanje i uspravno hodanje. Kao rezultat toga, pacijent ima nesigurnost, nesigurnost hoda. U hodu tijelo mu se naginje unazad. (Hennerov znak) stavlja noge u ravnu liniju (hod lisice) ponekad kod hodanja postoji "pletenje" nogu. U nekih bolesnika s oštećenjem prednjih dijelova frontalnih režnjeva razvija se neobičan fenomen: u nedostatku paralize i pareze i očuvane sposobnosti potpunog pokretanja nogu, bolesnici ne mogu stajati (astazija) i hodati (abazija).
Poraz asocijativnih zona korteksa često je karakteriziran razvojem kliničkih manifestacija kršenja viših mentalnih funkcija (vidi Poglavlje 15).
Opći pregled građe hemisfera velikog mozga
Hemisfere velikog mozga su najmasovniji dio mozga. Prekrivaju mali mozak i moždano deblo. Hemisfere velikog mozga čine približno 78% ukupne mase mozga. U procesu ontogenetskog razvoja organizma iz medule neuralne cijevi razvijaju se moždane hemisfere, pa se ovaj dio mozga još i naziva terminalni mozak.
Hemisfere velikog mozga podijeljene su duž središnje linije dubokom vertikalnom pukotinom na desnu i lijevu hemisferu.
U dubini srednjeg dijela obje hemisfere su međusobno povezane velikom priraslicom - corpus callosum. U svakoj hemisferi razlikuju se režnjevi; frontalni, parijetalni, temporalni, okcipitalni i insula.
Režnjevi moždanih hemisfera međusobno su odvojeni dubokim brazdama. Najvažnije su tri duboke brazde: središnja (Rolandova) koja odvaja frontalni režanj od parijetalnog, lateralna (Sylvian) koja odvaja temporalni režanj od parijetalnog, parijetalno-okcipitalna koja odvaja parijetalni režanj od okcipitalnog na unutarnjoj površini hemisfera.
Svaka hemisfera ima gornju bočnu (konveksnu), donju i unutarnju površinu.
Svaki režanj hemisfere ima cerebralne vijuge, međusobno odvojene brazdama. Odozgo je hemisfera prekrivena korom - tankim slojem sive tvari, koja se sastoji od živčanih stanica.
Moždana kora najmlađa je evolucijska tvorevina središnjeg živčanog sustava. Kod čovjeka dostiže najviši razvoj. Cerebralni korteks ima veliku važnost u regulaciji vitalne aktivnosti tijela, u provedbi složenih oblika ponašanja i formiranju neuropsihičkih funkcija.
Pod korteksom je bijela tvar hemisfera, sastoji se od procesa živčanih stanica - vodiča. Zbog stvaranja cerebralnih vijuga, ukupna površina moždane kore značajno se povećava. ukupna površina hemisferni korteks je 1200 cm 2, s 2/3 njegove površine smještene u dubini brazda, a 1/3 na vidljivoj površini hemisfera. Svaki režanj mozga ima različito funkcionalno značenje.
a - gornja bočna površina: 1 - donji frontalni girus; 2 - srednji frontalni gyrus; 3 - gornji frontalni girus; 4 - prednji središnji gyrus; 5 - središnja (Rolandova) brazda; 6 - stražnji središnji gyrus; 7 - gornji parijetalni lobule; 8 - donji parijetalni režanj; 9 - supramarginalna (supramarginalna) brazda; 10 - kutna (kutna) brazda; 11 - parieto-okcipitalni sulkus; 12 - donji temporalni gyrus;
13 - srednji temporalni gyrus; 14 - gornji temporalni gyrus; 15 - bočna (Sylvian) brazda;
b - unutarnja površina: 1 paracentralni lobule; 2 - središnja brazda; 3 - cingularni gyrus: 4 - corpus callosum; 5 - parieto-okcipitalni sulkus; 6 - klin; 7 - ostružna brazda; 8 - lingularni gyrus; 9 - girus hipokampusa (parahipokampalni girus).
Struktura frontalnog režnja
Frontalni režanj zauzima prednje dijelove hemisfera. Od parijetalnog režnja odvojen je središnjim sulkusom, a od temporalnog režnja lateralnim sulkusom. Četiri su vijuge u frontalnom režnju: jedna vertikalna - precentralna i tri horizontalne - gornji, srednji i donji frontalni girus. Zavoji su međusobno odvojeni brazdama.
Na donjoj površini frontalnih režnjeva razlikuju se izravni i orbitalni girus. Izravna vijuga nalazi se između unutarnjeg ruba hemisfere, olfaktornog žlijeba i vanjskog ruba hemisfere.
U dubini njušne brazde nalaze se olfaktorni bulbus i olfaktorni trakt.
Ljudski frontalni režanj čini 25–28% korteksa; prosječna masa frontalnog režnja je 450 g.
Funkcija frontalnih režnjeva povezana s organizacijom voljnih pokreta, motoričkim mehanizmima govora, regulacijom složenih oblika ponašanja, misaonim procesima. Nekoliko funkcionalno važnih centara koncentrirano je u vijugama frontalnog režnja. Prednji središnji girus je "prikaz" primarne motoričke zone sa strogo definiranom projekcijom dijelova tijela. Lice se nalazi u donjoj trećini girusa, ruka je u srednjoj trećini, noga je u gornja trećina. Trup je predstavljen u stražnjim dijelovima gornjeg frontalnog girusa. Dakle, osoba se projicira u prednjem središnjem girusu naopako i glavom prema dolje.
Prednji središnji girus, zajedno sa susjednim stražnjim i frontalnim vijugama, ima vrlo funkcionalno važnu ulogu. To je središte dobrovoljnih pokreta. U dubini korteksa središnjeg girusa, od takozvanih piramidalnih stanica - središnjeg motoričkog neurona - počinje glavni motorički put - piramidalni, kortikospinalni, staza. Periferni procesi motoričkih neurona izlaze iz korteksa, okupljaju se u jedan snažan snop, prolaze kroz središnju bijelu tvar hemisfera i ulaze u moždano deblo kroz unutarnju kapsulu; na kraju moždanog debla se djelomično križaju (prelazeći s jedne strane na drugu) i zatim se spuštaju u leđnu moždinu. Ove grane završavaju u siva tvar leđna moždina. Tu dolaze u kontakt s perifernim motoričkim neuronom i prenose mu impulse iz središnjeg motoričkog neurona. Impulsi voljnog kretanja prenose se duž piramidalnog puta.
U stražnjim dijelovima gornjeg frontalnog gyrusa također postoji ekstrapiramidalni centar korteksa, anatomski i funkcionalno usko povezana s tvorevinama takozvanog ekstrapiramidalnog sustava. Ekstrapiramidalni sustav - pogonski sustav za pomoć pri dobrovoljnom kretanju. Ovo je sustav "osiguranja" proizvoljnih pokreta. Budući da je filogenetski stariji, ljudski ekstrapiramidalni sustav osigurava automatsku regulaciju "naučenih" motoričkih činova, održavajući opću tonus mišića, spremnost perifer lokomotivnog aparata na pravljenje pokreta, preraspodjela mišićnog tonusa tijekom pokreta. Osim toga, uključen je u održavanje normalnog držanja.
motorna područja korteksa nalaze se uglavnom u precentralnom girusu (polja 4 i 6) i paracentralnom režnju na medijalnoj površini hemisfere. Postoje primarna i sekundarna područja - polja 4 i 6. Ova polja su motorička, ali se po svojim karakteristikama, prema istraživanjima Instituta za mozak, razlikuju. U primarnom motornom korteksu(polje 4) nalaze se neuroni koji inerviraju motoričke neurone mišića lica, trupa i udova.
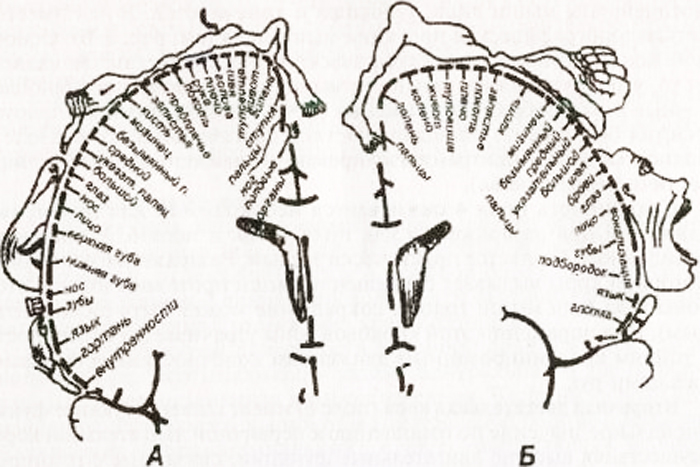
Shema autotopičke projekcije opće osjetljivosti i motoričkih funkcija u moždanoj kori (prema W. Penfieldu):
A - kortikalna projekcija opće osjetljivosti; B - kortikalna projekcija motornog sustava. Relativne veličine organa odražavaju područje moždane kore iz kojeg se mogu izazvati odgovarajući osjećaji i pokreti.
Ima jasnu topografsku projekciju mišića tijela. Glavni obrazac topografskog prikaza je da regulacija aktivnosti mišića koji pružaju najtočnije i najrazličitije pokrete (govor, pisanje, izrazi lica) zahtijeva sudjelovanje velikih područja motoričkog korteksa. Polje 4 u potpunosti je zauzeto centrima izoliranih kretanja, polje 6 samo djelomično (potpolje 6a).
Očuvanje polja 4 pokazalo se nužnim za postizanje pokreta tijekom stimulacije i polja 4 i polja 6. U novorođenčeta je polje 4 praktički zrelo. Stimulacija primarnog motoričkog korteksa uzrokuje kontrakciju mišića suprotna strana tijelo (za mišiće glave kontrakcija može biti obostrana). S porazom ove kortikalne zone izgubljena je sposobnost finih koordiniranih pokreta udova, a posebno prstiju.
sekundarni motorni korteks(polje 6) ima dominantno funkcionalno značenje u odnosu na primarni motorički korteks, obavljajući više motoričke funkcije povezane s planiranjem i koordinacijom voljnih pokreta. Evo, polako raste negativno potencijal spremnosti, javlja se otprilike 1 s prije početka kretanja. Korteks polja 6 prima većinu impulsa iz bazalnih ganglija i malog mozga, te je uključen u kodiranje informacija o složenim pokretima.
Iritacija korteksa polja 6 uzrokuje složene koordinirane pokrete, kao što su okretanje glave, očiju i trupa u suprotnom smjeru, prijateljske kontrakcije fleksora ili ekstenzora na suprotnoj strani. Premotorni korteks sadrži motoričke centre povezane s društvene funkcije ljudski: centar pisanog govora u stražnjem dijelu srednjeg frontalnog girusa (polje 6), centar Brocinog motoričkog govora u stražnjem dijelu donjeg frontalnog girusa (polje 44), koji osigurava govor, kao i glazbenu motoriku centar (polje 45), koji osigurava tonalitet govora, sposobnost pjevanja . Donji dio polja b (podpolje bora), koji se nalazi u području gume, reagira na električnu struju ritmičnim pokretima žvakanja. Neuroni motoričkog korteksa primaju aferentne ulaze kroz talamus od receptora mišića, zglobova i kože, iz bazalnih ganglija i malog mozga. Glavni eferentni izlaz motoričkog korteksa do matičnih i spinalnih motoričkih centara su piramidalne stanice sloja V.
U stražnjem dijelu srednjeg frontalnog girusa nalazi se frontalni okulomotorni centar, koji kontrolira prijateljsku, istodobnu rotaciju glave i očiju (središte rotacije glave i očiju u suprotnom smjeru). Iritacija ovog centra uzrokuje okretanje glave i očiju u suprotnom smjeru. Funkcija ovog centra je od velike važnosti u provedbi tzv. orijentacijskih refleksa (ili refleksa "što je?"), koji su vrlo važni za očuvanje života životinja.
Frontalni korteks moždanih hemisfera također prima Aktivno sudjelovanje u formiranju mišljenja, organizaciji svrhovitih aktivnosti, dugoročnom planiranju.
Građa parijetalnog režnja
Parijetalni režanj zauzima gornje bočne površine hemisfere. Od prednjeg parijetalnog režnja, sprijeda i sa strane, ograničen je središnjim sulkusom, od temporalnog odozdo - lateralnim sulkusom, od okcipitalnog - zamišljenom linijom koja ide od gornjeg ruba parijetalno-okcipitalnog sulkusa do donji rub hemisfere.
Na gornjoj bočnoj površini parijetalnog režnja nalaze se tri zavoja: jedan okomiti - stražnji središnji i dva vodoravna - gornji parijetalni i donji parijetalni. Dio donjeg parijetalnog girusa, koji obavija stražnji dio lateralnog sulkusa, naziva se supramarginalno (supramarginalno), dio koji okružuje gornji temporalni girus naziva se nodalno (angularno) područje.
Parijetalni režanj, kao i frontalni režanj, čini značajan dio moždanih hemisfera. U filogenetskom smislu, u njemu se razlikuje stari dio - stražnji središnji girus, novi - gornji parijetalni girus i noviji - donji parijetalni girus.
Funkcija parijetalnog režnja povezana je s percepcijom i analizom osjetljivih podražaja, prostornom orijentacijom. Nekoliko funkcionalnih centara koncentrirano je u vijugama parijetalnog režnja.
U stražnjem središnjem girusu, centri osjetljivosti se projiciraju s tjelesnom projekcijom sličnom onom u prednjem središnjem girusu. Lice se projicira u donjoj trećini vijuge, ruka i torzo projiciraju se u srednjoj trećini, a noga se projicira u gornjoj trećini. U gornjoj parijetalnoj vijugi nalaze se centri odgovorni za složene vrste duboka osjetljivost: mišićno-zglobni, dvodimenzionalno-prostorni osjet, osjet težine i volumena pokreta, osjet za prepoznavanje predmeta dodirom.
Iza gornjih dijelova stražnjeg središnjeg girusa lokaliziran je centar koji omogućuje prepoznavanje vlastitog tijela, njegovih dijelova, njihovih proporcija i međusobnog položaja (polje 7).
Polja 1, 2, 3 postcentralnog područja čine glavnu kortikalnu jezgru kožnog analizatora. Zajedno s poljem 1, polje 3 je primarna, a polje 2 sekundarna projekcijska zona kože. analizator. Postcentralna regija povezana je eferentnim vlaknima sa subkortikalnim i matičnim formacijama, s precentralnom i drugim područjima moždane kore. Dakle, kortikalni dio osjetljivog analizatora lokaliziran je u parijetalnom režnju.
Primarna osjetilna područja - to su područja osjetilne kore, čija iritacija ili uništenje uzrokuje jasne i trajne promjene u osjetljivosti tijela (jezgra analizatora, prema I.P. Pavlovu). Sastoje se uglavnom od monomodalnih neurona i tvore osjete iste kvalitete. U osnovnoj osjetilna područja obično postoji jasan prostorni (topografski) prikaz dijelova tijela, njihovih receptorskih polja.
Oko primarnih osjetnih područja manje su lokalizirana sekundarna osjetilna područjačiji neuroni odgovaraju na djelovanje više podražaja, tj. oni su polimodalni.
Najvažnije osjetilno područje je parijetalni korteks postcentralni girus i odgovarajući dio paracentralnog lobula na medijalnoj površini hemisfera, koji se označava kao somatosenzorno područje I. Ovdje postoji projekcija osjetljivosti kože suprotne strane tijela od taktilnih, bolnih, temperaturnih receptora, interoceptivne osjetljivosti i osjetljivosti mišićno-koštanog sustava - od mišićnih, zglobnih, tetivnih receptora.
Osim somatosenzornog područja I, postoje i manje veličine somatosenzorni regija II, nalazi se na granici sjecišta središnjeg sulkusa s gornjim rubom temporalnog režnja, u dubini bočnog sulkusa. Ovdje je manje izražen stupanj lokalizacije dijelova tijela.
Smješten u donjem parijetalnom režnju praxis centri. Praxis se podrazumijeva kao svrhoviti pokreti koji su postali automatizirani u procesu ponavljanja i vježbi, a koji se razvijaju u procesu učenja i stalnog vježbanja tijekom individualnog života. Hodanje, jelo, oblačenje, pisanje mehanički element, razne vrste radna aktivnost(primjerice, kretanje vozača u vožnji, košnja itd.) su praksa. Praxis je najviša manifestacija motoričke funkcije čovjeka. Provodi se kao rezultat kombinirane aktivnosti različitih područja moždane kore.
U donjim dijelovima prednje i stražnje središnje vijuge nalaze se centar za analizu interoceptivnog impulsa unutarnji organi i posude. Središte ima bliske veze s subkortikalnim vegetativnim formacijama.
Struktura temporalnog režnja
Temporalni režanj zauzima inferolateralnu površinu hemisfera. Od frontalnog i parijetalnog režnja, temporalni režanj je ograničen lateralnim žlijebom. Na gornjoj bočnoj površini temporalnog režnja nalaze se tri zavoja: gornji, srednji i donji.
Gornji temporalni girus nalazi se između silvijeve i gornje temporalne brazde, srednji girus leži između gornjeg i donjeg temporalnog sulkusa, a donji girus leži između donjeg temporalnog sulkusa i transverzalne medularne fisure. Na donjoj površini temporalnog režnja razlikuju se donji temporalni girus, lateralni okcipitotemporalni girus i girus hipokampusa (noge morskog konja).
Funkcija temporalnog režnja povezana je s percepcijom slušnih, okusnih, olfaktornih osjeta, analizom i sintezom zvukova govora i mehanizmima pamćenja. Glavno funkcionalno središte gornje lateralne površine temporalnog režnja nalazi se u gornjem temporalnom vijugu. Ovdje se nalazi slušno, odnosno gnostičko središte govora (Wernickeovo središte).
Dobro proučena zona primarne projekcije je slušni korteks(polja 41, 42, 52), koji se nalazi u dubini lateralnog sulkusa (korteks transverzalnih Heschlovih temporalnih vijuga). Projekcijski korteks temporalnog režnja također uključuje centar za vestibularni analizator● u gornjem i srednjem temporalnom vijugu (polja 20 i 21).
Područje olfaktorne projekcije nalazi se u girusu hipokampusa, posebno u njegovom prednjem dijelu (tzv. kuka). Uz olfaktorne projekcijske zone nalaze se gustativne.
Temporalni režnjevi igraju važnu ulogu u organizaciji složenih mentalnih procesa, posebice pamćenja.
Struktura okcipitalnog režnja
Okcipitalni režanj zauzima stražnje dijelove hemisfera. Na konveksnoj površini hemisfere, okcipitalni režanj nema oštre granice koje ga odvajaju od parijetalnog i temporalnog režnja, s izuzetkom gornjeg dijela parijetalno-okcipitalnog sulkusa, koji se nalazi na unutarnjoj površini hemisfere. odvaja parijetalni režanj od okcipitalnog režnja. Brazde i vijuge gornje bočne površine okcipitalnog režnja su nestabilne i imaju varijabilnu strukturu. Na unutarnjoj površini okcipitalnog režnja nalazi se utor koji odvaja klin (trokutasta norma lobule okcipitalnog režnja) od lingvalne vijuge i okcipitotemporalne vijuge.
Funkcija okcipitalnog režnja povezana je s percepcijom i obradom vizualnih informacija, organizacijom složenih procesa vizualne percepcije - Istodobno, gornja polovica mrežnice projicira se u području klina, koji percipira svjetlost iz donjih vidnih polja; u području lingularnog girusa je donja polovica mrežnice koja percipira svjetlost iz gornjih vidnih polja.
Smješten u okcipitalnom korteksu primarno vidno područje(korteks dijela sfenoidnog girusa i lingularnog lobula, polje 17). Ovdje je aktualan prikaz retinalnih receptora. Svaka točka mrežnice odgovara svom području vidnog korteksa, dok zona makule ima relativno veliku zonu zastupljenosti. U vezi s nepotpunim križanjem vidnih putova, iste polovice retine projiciraju se u vidno područje svake hemisfere. Prisutnost u svakoj hemisferi projekcije mrežnice oba oka osnova je binokularnog vida. Kora se nalazi blizu polja 17 sekundarno vidno područje(polja 18 i 19). Neuroni ovih zona su polimodalni i reagiraju ne samo na svjetlo, već i na taktilne i slušne podražaje. Sinteza se događa u ovom vizualnom području razne vrste osjetljivosti, nastaju složenije vizualne slike i provodi se njihovo prepoznavanje.
Otočić, limbički korteks
otok, ili tzv. zatvorena lobula, smještena duboko u bočnom utoru. Otočić je od susjednih susjednih dijelova odvojen kružnim utorom. Površina otočića podijeljena je svojom uzdužnom središnjom brazdom na prednji i stražnji dio. U otočiću je projiciran analizator okusa.
limbički korteks. Na unutarnjoj površini hemisfera iznad corpus callosum nalazi se cingulate gyrus. Ova vijuga, s istmusom iza corpus callosuma, prelazi u vijugu u blizini morskog konjica - parahipokampalni girus. Cingulatni girus zajedno s parahipokampalnim girusom čini zasvođeni girus.
Unutarnje i donje površine hemisfera spojene su u takozvani limbički (rubni) korteks, zajedno s jezgrom amigdale iz skupine subkortikalnih jezgri, olfaktornim traktom i bulbusom, područjima frontalnog, temporalnog i parijetalnog režnja cerebralnim korteksom, kao i s hipotuberoznom regijom i retikularnom formacijom trupa. Limbički korteks sjedinjuje se u jednu funkcionalni sustav- limbičko-retikularni kompleks. Glavna funkcija ovih dijelova mozga nije toliko osigurati komunikaciju s vanjskim svijetom, koliko regulirati tonus korteksa, nagone i afektivni život. Oni reguliraju složene, višestrane funkcije unutarnjih organa i reakcije ponašanja. Limbičko-retikularni kompleks je najvažniji integrativni sustav tijela. limbički sustav također je važan u formiranju motivacije. Motivacija (ili unutarnja motivacija) uključuje najsloženije instinktivne i emocionalne reakcije (na hranu, obrambene, seksualne). Limbički sustav također je uključen u regulaciju spavanja i budnosti.
Limbički korteks također obavlja važnu funkciju mirisa. Njuh – percepcija boravka u zraku kemijske tvari. Ljudski olfaktorni mozak osigurava osjet mirisa, kao i organizaciju složenih oblika emocionalnih i bihevioralnih reakcija. Olfaktorni mozak dio je limbičkog sustava.
Olfaktorni mozak sastoji se od dva dijela - perifernog i središnjeg. Periferni dio predstavljen je olfaktornim živcem, olfaktornim žaruljama, primarnim olfaktornim centrima. Središnji dio uključuje vijugu morskog konja - hipokampus, nazubljenu i zasvođenu vijugu.
Njušni receptorski aparat nalazi se u nosnoj sluznici. Kroz sustav živčanih vodiča informacije s receptora prenose se u kortikalni dio olfaktornog analizatora.
Kortikalno područje olfaktornog analizatora nalazi se u cingulatnom girusu, girusu morskog konjica i u kuki morskog konjica, koji zajedno tvore zatvoreno prstenasto područje. Periferni dio olfaktornog analizatora povezan je s kortikalnim regijama obiju hemisfera.
Fiziološki mehanizam percepcije mirisa olfaktornim analizatorom nije sasvim jasan. Postoje dvije glavne hipoteze koje objašnjavaju prirodu ovog procesa s različitih pozicija. Prema jednoj od hipoteza, interakcija između molekula mirisa i kemoreceptora odvija se poput ključa i brave, tj. vrsta molekule odgovara posebnom receptoru. Druga hipoteza temelji se na pretpostavci da molekule mirisne tvari imaju određeni val oscilacije, na koji su "podešeni" olfaktorni receptori. Molekule koje imaju slične vibracije trebale bi imati zajednički val i, prema tome, davati slične mirise.
Pojam "olfaktorni mozak" u odnosu na ljudsku fiziologiju je pomalo proizvoljan i ne otkriva u potpunosti njegovu višestruku i univerzalnu funkciju. "Smještaj" središnje karike olfaktornog mozga u moždane hemisfere nije slučajan i rezultat je goleme "informacijske" uloge koju je osjetilo mirisa imalo u procesu evolucije u prilagodbi vanjskom okruženju i regulaciji kompleksa reakcije ponašanja. Pribavljanje hrane, odabir jedinke suprotnog spola, briga za potomstvo, cjelovitost teritorija, organiziranje grupnih zajednica unutar vrste - sve te dnevne funkcije kod mnogih životinja obavljaju se uz izravno sudjelovanje fino izgrađenog sustava njušne recepcije i , na temelju toga, sposobnost niza životinja da šalju suptilno diferencirane specifične mirisne tvari - signale-informatore.
U životu ljudi osjetilo mirisa izgubilo je biološku informacijsku vrijednost koju je imalo kod životinja. Ljudski olfaktorni sustav dizajniran je kako za obavljanje uske, "vlastite" funkcije, tako i za svojevrsno "punjenje" emocija. Snaga utjecaja mirisa na emocionalnu sferu, da su oni najvažniji "prehrambeni supstrat emocija", dobro je poznata od davnina u povijesti čovječanstva.
Osjetilo mirisa osobe može varirati. U pravilu su ove varijacije beznačajne, ali u nekim slučajevima oštrina mirisa može biti vrlo visoka (kušači u industriji parfema).
Budući da olfaktorni analizator igra važnu ulogu u regulaciji emocija, njegov centralni odjel odnosi se na limbički sustav, slikovito nazvan "zajednički nazivnik" za mnoge emocionalne i viscerosomatske reakcije tijela.
Centar za analizu okusa nalazi se u neposrednoj blizini centra olfaktornog analizatora, tj. u kuki i Amonovom rogu, ali, dodatno, u najnižem dijelu stražnjeg središnjeg girusa (polje 43), kao iu insuli. Poput olfaktornog analizatora, centar osigurava funkciju projekcije, pohranu i prepoznavanje uzoraka okusa.
Na granici temporalnog, okcipitalnog i parijetalnog režnja nalazi se centar za analizu pisanog govora(polje 39), koji je usko povezan sa Wernickeovim centrom temporalnog režnja, sa centrom vidnog analizatora okcipitalnog režnja, kao i sa centrima parijetalnog režnja. Centar za čitanje omogućuje prepoznavanje i pohranu slika pisanog govora.
Glavni dio informacija oko okolina i unutarnja sredina tijela, koja je ušla u osjetilni korteks, prenosi se na daljnju obradu u asocijativni korteks, nakon čega se (ako je potrebno) pokreće reakcija ponašanja uz obvezno sudjelovanje motoričkog korteksa.
Corpus callosum- lučna tanka ploča, filogenetski mlada, povezuje srednje površine obiju hemisfera. Izduženi srednji dio corpus callosuma straga prelazi u zadebljanje, a naprijed se lučno izvija i zavija prema dolje. Corpus callosum povezuje filogenetski najmlađe dijelove hemisfera i ima važnu ulogu u razmjeni informacija među njima.
Arhitektonika kore velikog mozga
Naziva se doktrina o strukturnim značajkama strukture korteksa arhitektonika. Stanice cerebralnog korteksa manje su specijalizirane od neurona drugih dijelova mozga; ipak, određene skupine njih su anatomski i fiziološki usko povezane s određenim specijaliziranim dijelovima mozga.
Mikroskopska građa kore velikog mozga nije ista u različitim dijelovima. Ove morfološke razlike u korteksu omogućile su razlikovanje pojedinih kortikalnih citoarhitektonskih polja. Postoji nekoliko opcija za klasifikaciju kortikalnih polja. Većina istraživača identificira 50 citoarhitektonskih polja čija je mikroskopska struktura prilično složena.
Korteks se sastoji od 6 slojeva stanica i njihovih vlakana. Glavni tip strukture šesteroslojne kore, međutim, nije svugdje ujednačen. Postoje područja korteksa gdje je jedan od slojeva izražen značajno, a drugi je slabo izražen. U drugim dijelovima kore neki su slojevi podijeljeni u podslojeve i tako dalje.
Utvrđeno je da područja korteksa povezana s određenom funkcijom imaju sličnu strukturu. Područja kore koja su po svom funkcionalnom značaju bliska kod životinja i ljudi imaju određenu sličnost u strukturi. Ona područja mozga koja obavljaju isključivo ljudske funkcije (govor) prisutna su samo u ljudskom korteksu, dok životinja, čak i majmuna, nema.
Morfološka i funkcionalna heterogenost cerebralnog korteksa omogućila je izdvajanje centara za vid, sluh, miris itd., koji imaju svoju specifičnu lokalizaciju. Međutim, netočno je govoriti o kortikalnom centru kao o strogo ograničenoj skupini neurona. Specijalizacija područja korteksa formira se u procesu života. U rano djetinjstvo funkcionalne zone korteksa se međusobno preklapaju pa su im granice nejasne i nejasne. Samo u procesu učenja, akumulacije vlastito iskustvo praktične aktivnosti dolazi do postupnog koncentriranja funkcionalnih zona u središta međusobno odvojena.
Bijela tvar hemisfera velikog mozga sastoji se od živčanih vodiča. U skladu s anatomskim i funkcionalnim značajkama vlakna bijele tvari dijele se na asocijativna, komisuralna i projekcijska.
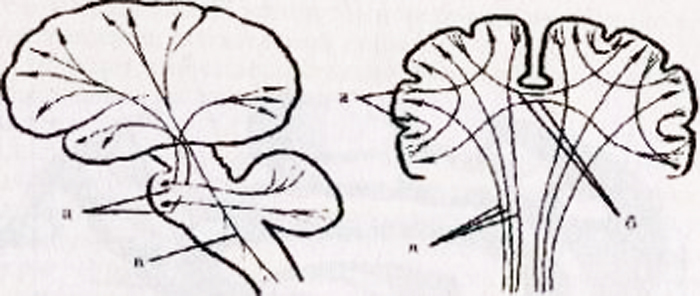
a – asocijacijska vlakna; b - komisuralna vlakna; vlakna u projekciji.
Asocijativni vlakna se sjedinjuju razne sekcije korteks unutar jedne hemisfere. Ova vlakna su kratka i duga. Kratka vlakna su obično lučna i povezuju susjedne vijuge. Duga vlakna povezuju udaljene dijelove korteksa. Uobičajeno je da se komisuralnim vlaknima nazivaju ona vlakna koja povezuju topografski identične dijelove desne i lijeve hemisfere. Komisuralna vlakna tvore tri komisure: prednju bijelu komisuru, komisuru forniksa i corpus callosum. Prednja bijela komisura povezuje olfaktorne regije desne i lijeve hemisfere. Forniks komisura povezuje hipokampalne vijuge desne i lijeve hemisfere. Glavna masa komisuralnih vlakana prolazi kroz corpus callosum, povezujući simetrične dijelove obje hemisfere mozga.
projekcija nazivaju ona vlakna koja povezuju moždane hemisfere s donjim dijelovima mozga – deblom i leđnom moždinom. U sklopu projekcijskih vlakana nalaze se provodni putovi koji nose aferentne (osjetljive) i eferentne (motoričke) informacije.
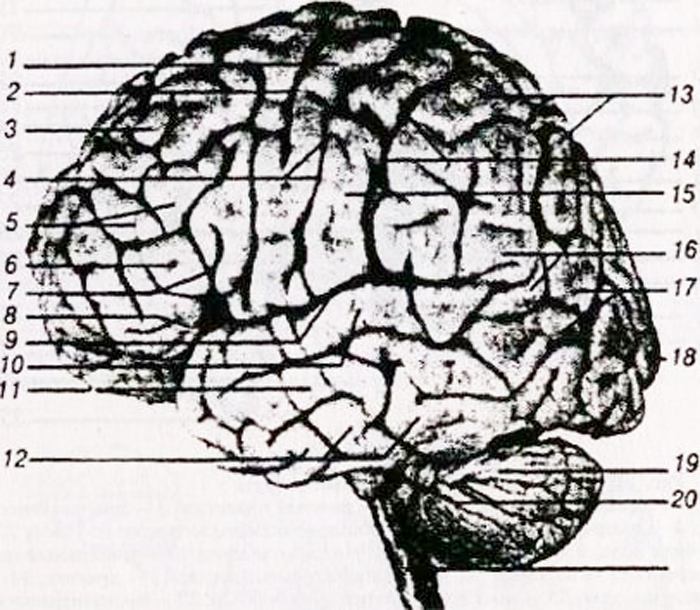
1 - precentralni gyrus; 2 - precentralna brazda; 3 - gornji frontalni girus; 4 - središnja brazda; 5 - srednji frontalni gyrus;
b - donji frontalni girus; 7 - uzlazna grana bočne brazde;
8 - vodoravna grana bočne brazde; 9 - stražnja grana bočna brazda; 10 - gornji temporalni gyrus; 11 - srednji temporalni gyrus;
12 - donji temporalni gyrus; 13 - parijetalni režanj; 14 - post-centralna brazda; 15 - postcentralni gyrus; 16 - supramarginalni gyrus;
17 - kutni gyrus; 18 - okcipitalni režanj; 19 - mali mozak; 20 - horizontalna pukotina malog mozga; 21 - produžena moždina

