Lipidi 200401
CHIMICA E METABOLISMO DEI LIPIDI.
I LIPIDI sono sostanze organiche caratteristiche degli organismi viventi, insolubili in acqua, ma solubili nei solventi organici e tra loro.
I gruppi lipidici differiscono nel grado di idrofobicità. Fosfolipidi e glicolipidi sono lipidi polari.
Il colesterolo occupa una posizione intermedia tra i lipidi polari e quelli assolutamente idrofobi.
I trigliceridi e gli esteri del colesterolo sono assolutamente idrofobi.
La maggior parte dei lipidi (eccetto gli steroli e alcuni lipidi minori) ne contengono una quantità maggiore acido grasso(VZHK).
Le membrane contengono solo FOSFOLIPIDI (PL), GLICOLIPIDI (GL) e COLESTEROLO (CS).
ACIDI GRASSI CARATTERISTICI DEL CORPO UMANO
Un acido si dice grasso se il numero di atomi di carbonio nella sua molecola è superiore a quattro. Predominano gli acidi grassi a catena lunga (il numero di atomi di carbonio è pari o superiore a 16).Il numero di atomi di carbonio e di doppi legami è indicato da un doppio indice. Ad esempio: C18:1 (9-10). In questo caso, 18 è il numero di atomi di carbonio e 1 è il numero di doppi legami. La posizione dei doppi legami (per numero di atomi di carbonio) è indicata tra parentesi.
Gli studenti dovrebbero conoscere il numero di atomi di carbonio e di doppi legami nelle seguenti molecole di acidi grassi:
C16:0 - palmitico,
C18:0 - stearico,
C18:1 – oleico (9:10),
C18:2 – linoleico (9-10,12-13),
C18:3 - linolenico (9-10, 12-13, 15-16),
C20:4 - arachidonico (5-6, 8-9, 12-13, 15-16).
Gli acidi grassi che hanno un doppio legame in posizione 3 hanno un effetto antiaterosclerotico molto pronunciato (acido linolenico). Soprattutto molti acidi grassi contenenti 3 si trovano nell'olio di pesce e in altri frutti di mare. Tali acidi grassi non sono sintetizzati nel corpo umano.
Gli acidi grassi che compongono il corpo umano hanno caratteristiche comuni edifici:
Numero pari di atomi di carbonio.
Catena di carbonio lineare (non ramificata).
Gli acidi grassi polinsaturi hanno SOLO doppi legami ISOLATI (ci sono almeno due legami singoli tra doppi legami adiacenti).
I doppi legami hanno solo una configurazione cis.
In base al numero di doppi legami, gli acidi grassi possono essere classificati come SATURI (nessun doppio legame), MONOINSATURI (un doppio legame) e POLINSATURI (due o più doppi legami).
Gli acidi grassi liberi si trovano in quantità molto piccole. Si trovano principalmente in altri lipidi. Inoltre sono legati ad altri componenti lipidici tramite un legame estere (esterificato).
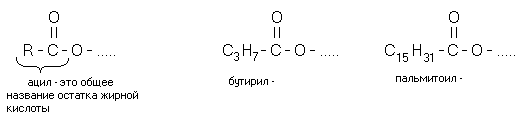
FORME DI ESISTENZA DEI LIPIDI NEL CORPO UMANO
Inclusioni grasse, più abbondanti negli adipociti. Questa è la forma di esistenza dei trigliceridi.
Biomembrane. Non contengono trigliceridi, ma contengono fosfolipidi, glicolipidi e colesterolo.
Lipoproteine. Può includere lipidi di tutte le classi.
Lipoproteine
Le lipoproteine sono particelle sferiche in cui si può distinguere un nucleo idrofobico, costituito da trigliceridi (TRG) ed esteri del colesterolo (ECS) e un guscio anfifilico, che contiene fosfolipidi, glicolipidi e proteine.
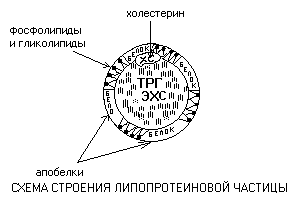
Le proteine del guscio sono chiamate apoproteine. Il colesterolo (CS) occupa solitamente una posizione intermedia tra il guscio e il nucleo. I componenti di una particella sono collegati da legami deboli e sono in uno stato di diffusione costante, capaci di muoversi l'uno rispetto all'altro.
Il ruolo principale delle lipoproteine è il trasporto dei lipidi, quindi possono essere trovate nei fluidi biologici.
Studiando i lipidi del plasma sanguigno, si è scoperto che possono essere divisi in gruppi, poiché differiscono l'uno dall'altro nel rapporto dei componenti. Lipoproteine diverse hanno rapporti diversi tra lipidi e proteine nella composizione delle particelle, quindi anche la densità è diversa.
Le lipoproteine vengono separate in base alla densità mediante ultracentrifugazione; non si depositano, ma galleggiano (galleggiano). La misura di galleggiamento è costante di galleggiamento, indicato con S f (flottazione di Svedberg). In base a questo indicatore, si distinguono i seguenti gruppi di lipoproteine:

Le lipoproteine possono anche essere separate mediante elettroforesi. Nell'elettroforesi alcalina classica, diverse lipoproteine si comportano diversamente. Quando le lipoproteine vengono poste in un campo elettrico, i chilomicroni rimangono all'inizio. VLDL e DILI si trovano nella frazione pre-β-globulina, LDL - nella frazione β-globulina e HDL - nelle β-globuline:
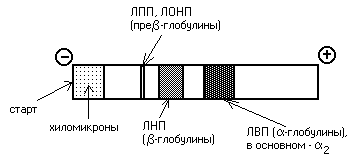
La determinazione dello spettro lipoproteico del plasma sanguigno viene utilizzata in medicina per diagnosticare l'aterosclerosi.
Tutte queste lipoproteine differiscono nella loro funzione.
1. Chilomicroni (HM)- si formano nelle cellule intestinali, la loro funzione è: trasferimento del grasso esogeno dall'intestino ai tessuti (principalmente tessuto adiposo), nonché trasporto del colesterolo esogeno dall'intestino al fegato.
2. Lipoproteine a densità molto bassa (VLDL)- si formano nel fegato, il loro ruolo: trasporto del grasso endogeno, sintetizzato nel fegato dai carboidrati, nel tessuto adiposo.
3. Lipoproteine a bassa densità (LDL)- si formano nel circolo sanguigno dalle VLDL attraverso lo stadio di formazione delle Lipoproteine a Densità Intermedia (IDL). Il loro ruolo: il trasporto del colesterolo endogeno nei tessuti.
4. Lipoproteine ad alta densità (HDL)- si formano nel fegato, il ruolo principale è il trasporto del colesterolo dai tessuti al fegato, cioè la rimozione del colesterolo dai tessuti, e quindi il colesterolo viene escreto con la bile.
Quando si determina il contenuto di lipoproteine di diversa densità nel sangue, queste vengono solitamente separate mediante elettroforesi. In questo caso i CM rimangono all'inizio, le VLDL finiscono nella frazione pre--globulina, le LDL e LPP si trovano nella frazione -globulina e le HDL - 2 -globuline. Se il contenuto di β-globuline (LDL) nel sangue aumenta, ciò significa che il colesterolo si deposita nei tessuti (si sviluppa l'aterosclerosi).
RUOLO DELLE PROTEINE APO NELLA COMPOSIZIONE DELLE LIPOPRETINE
Particolarmente importanti sono le componenti proteiche delle lipoproteine. Si chiamano " apolipoproteine" e sono designati con lettere latine (apoA, apoB, apoC e così via fino ad apoG).
FUNZIONI DELLE PROTEINE APO.
1. Le apoproteine agiscono come emulsionanti perché sono sostanze anfifiliche.
2. Alcune apolipoproteine sono regolatori dell'attività degli enzimi del metabolismo lipidico.
3. Può avere una propria attività enzimatica.
Possono agire come ligandi per i recettori cellulari delle lipoproteine.
Molte apoproteine trasportano i lipidi da una lipoproteina all’altra.
METABOLISMO DEI GRASSI (TRIAGLICEROLO)
Il nome chimico dei grassi è acilgliceroli, cioè grassi. Questi sono esteri del glicerolo e degli acidi grassi superiori. "Acil-" significa "residuo di acido grasso" (da non confondere con "acetil-" - residuo di acido acetico). A seconda del numero di radicali acilici, i grassi si dividono in mono-, di- e trigliceridi. Se la molecola contiene 2 radicali di acidi grassi, il grasso si chiama DIACILGLICEROLO. Se la molecola contiene 1 radicale di acido grasso, il grasso si chiama MONOACILGLICEROLO.
Nel corpo umano e animale predominano i TRIACILGLICEROLI (contengono tre radicali di acidi grassi).
Le proprietà del grasso sono determinate dalla composizione degli acidi grassi.
Funzioni dei grassi:
1. ENERGIA.
A differenza dei carboidrati, i grassi costituiscono la riserva energetica del corpo. Il vantaggio dei grassi come riserva energetica è che i grassi sono sostanze più ridotte rispetto ai carboidrati (le molecole di carboidrati hanno ossigeno in ciascun atomo di carbonio - gruppi “–CHOH-”; i grassi hanno lunghi radicali idrocarburici, in cui predominano i gruppi “”). CH 2 -“ - non hanno ossigeno). Una maggiore quantità di idrogeno può essere rimossa dal grasso, che poi passa attraverso la catena di ossidazione mitocondriale per produrre ATP.
Contenuto calorico di carboidrati e proteine: ~ 4 kcal/grammo.
Contenuto calorico dei grassi: ~ 9 kcal/grammo.
Il vantaggio dei grassi come riserva energetica, a differenza dei carboidrati, è la loro idrofobicità: non sono associati all'acqua. Ciò garantisce la compattezza delle riserve di grasso: vengono immagazzinate in forma anidra, occupando un piccolo volume.
La fornitura media di triacilgliceroli puri per una persona è di circa 13 kg. Queste riserve potrebbero essere sufficienti per 40 giorni di digiuno in condizioni di attività fisica moderata. Per fare un confronto: le riserve totali di glicogeno nel corpo sono di circa 400 g; durante il digiuno, questa quantità non è sufficiente nemmeno per un giorno.
2. PROTETTIVO.
Il tessuto adiposo:
a) protegge gli organi dai danni meccanici.
b) partecipa alla termoregolazione.
La formazione di riserve di grasso nel corpo dell'uomo e di alcuni animali è considerata un adattamento all'alimentazione irregolare e alla vita in un ambiente freddo. Gli animali che vanno in letargo per lungo tempo (orsi, marmotte) e che sono adattati a vivere al freddo (trichechi, foche) hanno una riserva di grasso particolarmente ampia. Il feto non ha praticamente grasso e appare solo prima della nascita.
FONTI DI GRASSO NEL CORPO:
1. Grasso commestibile ( eso genetico)
2. Endo gene del grasso, sintetizzato nel fegato dai carboidrati.
I lipidi sono insolubili in acqua, quindi vengono trasportati nel sangue in combinazione con le proteine. Gli acidi grassi nel sangue sono associati all'albumina, altri lipidi vengono trasportati come parte delle lipoproteine (LP).
Uno dei fattori che contribuiscono allo sviluppo del processo aterosclerotico è una violazione del metabolismo lipidico. Negli ultimi 25 anni, questo fatto ha portato a ricerche approfondite sulla struttura dei farmaci, sulla loro sintesi, sulle trasformazioni metaboliche e sull’internalizzazione delle particelle dei farmaci da parte delle cellule. Numerosi lavori presentano dati sulle proprietà e sulla composizione delle particelle di farmaci di diverse classi, nonché sulle caratteristiche della biosintesi e del catabolismo dei componenti lipidici e proteici.
Nella fig. 1.5 presenta le formule strutturali dei principali lipidi nel plasma sanguigno.
Gli acidi grassi liberi o non esterificati sono il principale fornitore di energia nel corpo. Il sangue umano contiene anche acidi grassi saturi, monoinsaturi e polinsaturi (Tabella 1.7).
Tabella 1.7. Acidi grassi presenti nel plasma Acidi grassi Nome Lunghezza
catena Fonte Myristic C 14:0 Olio di cocco Palmitic C saturo 16:0 Grassi animali Stearic C 18:0 Grassi animali Palmitoleic C monoinsaturi 16:1 co 7 Grassi Oleic C 18:1 co9 Burro Polinsaturi Linolenici C 18:2 Soob Vegetale
olio (poliene) Arachidonic C 20:4 co8 Vegetale
Olio eicosapentaenoico C 20:5 SOZ Grasso di pesce Nota. La formula abbreviata indica il numero di atomi di carbonio e il numero di doppi legami. Il doppio legame più vicino all'estremità metilica è indicato con il simbolo co.
I TG sono esteri del glicerolo e degli acidi grassi; loro, come gli acidi grassi, servono come un'importante fonte di energia.
I fosfolipidi sono esteri del glicerolo o della sfingosina con acidi grassi e acido fosforico. I fosfolipidi includono anche composti contenenti azoto (colina, etanolammina, serina) o altri gruppi idrofili collegati da un legame estere al fosfato. I fosfolipidi sono componenti di tutti membrane cellulari e LP.
CH3(CH2)„SOSG Acidi grassi (n> 14)
Trigliceridi®
O CH2 - O - C - (CH,)„ -CH3II I
CH, -(CH2)„ - s - O-sn circa
II
sn, - o - s - k.
Fosfolipidi O
CH, - O - C - (CH,)„ -SN,
IO
CH, ~(CH2)„ - s - o-cn o
sn2 - O - R - o -1
IO
O
Riso. 1.5. Formule strutturali lipidi.
Il colesterolo è uno steroide sulla base del quale si formano ormoni steroidei e acidi biliari. Circa 2/3 del colesterolo nel sangue è sotto forma di esteri del colesterolo degli acidi grassi e circa 1/3 è sotto forma di colesterolo libero. Il colesterolo libero fa parte di tutte le membrane cellulari; Il farmaco contiene sia colesterolo libero che esterificato.
IN pratica clinica Per identificare i disturbi del metabolismo lipidico, viene determinato il livello di colesterolo e TG nel siero del sangue.
LIPOPROTEINE
Le particelle del farmaco sono complessi macromolecolari, parte interna che contiene lipidi neutri (TG ed esteri del colesterolo), e lo strato superficiale è costituito da fosfolipidi, colesterolo non esterificato e specifiche proteine di trasporto dei lipidi chiamate apolipoproteine (apoLP).
I farmaci sono classificati in base alla loro mobilità in un campo elettrico (durante EP) o alla densità idrata in condizioni di gravità aumentata durante l'ultracentrifugazione preparativa (flottazione o sedimentazione). Quando si esegue l'EF, gli LP vengono divisi in frazioni, una delle quali rimane all'inizio (chilomicroni), altre migrano nelle zone globuliniche: R-LP, pre-r-LP, a-LP. In base al valore della densità idrata, i farmaci vengono solitamente suddivisi in 5 classi: chilomicroni (CM), farmaci a densità molto bassa (VLDL), farmaci a densità intermedia (IDL), farmaci a bassa densità (LDL), LP alta densità(HDL). In termini di mobilità elettroforetica, VLDL corrisponde a pre-r-LP, LDL - R-LP, HDL - a-LP e CM rimangono all'inizio (Fig. 1.6).
Gli ApoLP promuovono la formazione di micelle LP nel reticolo endoplasmatico degli epatociti, servono come ligandi per recettori specifici sulla superficie della membrana plasmatica delle cellule e cofattori (attivatori e inibitori) dei processi di lipolisi e metabolismo LP nel letto vascolare (Tabella 1.8).
A seconda del ruolo di apoLP nell'organizzazione delle particelle primarie del farmaco e nelle loro successive trasformazioni, le apoproteine delle condizioni
ma possono essere divisi in due classi. Uno di questi dovrebbe includere l'apoLP, che forma la struttura micellare dei complessi LP e funge da nucleo delle particelle LP. Una caratteristica di tali apoproteine è che le proteine non lasciano la particella lipidica alla cui formazione partecipano. Questo gruppo di apoLP comprende apoB (apoB-100 e apoB-48) e apoA (A-1 e A-H), responsabili del trasporto lipidico afferente ed efferente. ApoB è la principale proteina strutturale delle particelle lipidiche ricche di trigliceridi; non lascia la micellare
Tabella 1.8. Proprietà e funzioni delle apolipoproteine DpoLP Incluso Luogo
sintesi Mol.
peso,
kD Funzioni C* in
plasma, mg/l A-1 HDL,
hmm Sottile
intestino,
fegato 28.3 Attivazione di LCAT, trasporto dei lipidi, ligando per il recettore HDL 100-1600 A-N HDL Sottile
intestino,
fegato 17 Proteina strutturale, ligando per il recettore HDL 300-500 A-1U hmm,
lpvp,
lpon sottile
intestino 46 Attivazione di LCAT, ligando per il recettore HDL 150 B-100 LDL,
lppp,
lpon Liver 549 Trasferimento di lipidi dal fegato, legame ai recettori B, E 500-900 B-48 hmm Sottile
intestino 265 Trasferimento di lipidi dall'intestino tenue lpon,
Attivatore di fegato HDL 6.5 LCAT lponp,
HDL Fegato 8.8 Attivatore LPL 30-80 S-Sh hmm,
lponp.
HDL Fegato 8,9 Inibitore dell'attività LPL 8-150 B HDL3 20 Attivatore LCAT ~100 E lponp,
Hmm,
Fegato HDL 36,5 Legame ai recettori B ed E 30-50 (a) Fegato LP(a) 270-1000 Il complesso nel processo di successive trasformazioni metaboliche delle VLDL in LDLP e successivamente in LDL, il cui accumulo nel sistema vascolare il muro funge da nesso patogenetico processo aterosclerotico. Le apoproteine A-1 e A-H sono le principali proteine dell'HDL. La base per supporre che apoB e apoA formino classi di farmaci diverse per composizione e funzione era il fatto che apoB e apoA non sono presenti insieme nelle particelle di farmaco a lunga circolazione. Disturbi genetici nella sintesi di queste apoproteine sono la causa di disturbi nel trasporto dei lipidi afferenti ed efferenti.
Un'altra classe include gli apoLP, il loro ruolo principale è la regolazione del metabolismo dei lipidi nel letto vascolare e il processo di internalizzazione da parte delle cellule. Queste apoproteine sono contenute nel farmaco in quantità significativamente inferiori e, nel processo di interconversione delle particelle del farmaco nel flusso sanguigno, si spostano tra farmaci di classi diverse sotto forma di complessi proteina-lipidi. Le apoLP metabolicamente attive rimangono nel letto vascolare più a lungo delle particelle del farmaco in cui erano incluse al momento della sintesi. I principali rappresentanti del gruppo delle apoLP metabolicamente attive sono apoE (con le isoforme E2, E3, E4) e apoC (C-1, C-H, S-III).
ApoLP ha Grande importanza per il metabolismo dei farmaci, svolgendo il ruolo di ligandi delle particelle di farmaco che interagiscono con i recettori cellulari per farmaci specifici. ApoB-100 e apoE interagiscono con il recettore LDL, dando inizio all'endocitosi di assorbimento, accompagnata dal catabolismo delle LDL. L'ApoE interagisce con il recettore per i componenti residui del CM, che svolge un ruolo importante nella rimozione di questi componenti da parte del fegato. Si ritiene che l'apoA-1 interagisca con il recettore HDL e faciliti la rimozione del colesterolo dalle cellule periferiche per il trasporto di ritorno al fegato.
Il plasma contiene 4 classi principali di farmaci: colesterolo, VLDL, LDL e HDL e diversi altri farmaci presentati in concentrazioni relativamente basse: LDLP e LP(a). Nella tabella 1.9 presenta alcune proprietà del LP.
I ChM sono sintetizzati nell'intestino e servono a trasportare i lipidi esogeni. La proteina principale dei CM nascenti (primari) è apoB-48. Subito dopo la secrezione di CM da parte degli enterociti, due ulteriori apoproteine contenute nelle HDL, apoC-H e apoE, vengono incluse nella loro composizione nel letto vascolare. L'ApoC-H attiva la lipoproteina lipasi eparina-dipendente, un enzima fissato
P R a Luogo di formazione Intestino tenue Catabolismo epatico
VLDL Catabolismo delle VLDL attraverso LDLP Fegato, intestino tenue, catabolismo di CM e VLDL Funzione principale Trasporto di TG esogeni Trasporto di TG endogeni Precursore di Trasporto LDL
Inversione del colesterolo
trasporto
colesterolo Composizione: TG 90% 65% 20% 5% 5% colesterolo 5% 15% 25% 50% 20% fosfolipidi 4% 10% 35% 25% 25% proteine 1% 10% 20% 20% 50% apoproteine A, B-48, C, E B-100, C, E B-100, E B-100 A, C, E Tabella 1.9. Caratteristiche delle lipoproteine
1.5. Lipidi e lipoproteine -f- 143
sull'endotelio dei capillari. La lipoproteina lipasi idrolizza i TG dei chilomicroni per formare acidi grassi liberi. Durante l'idrolisi dell'LP, le particelle perdono circa il 96% della loro massa, principalmente a causa dei TG, nonché dell'apoLP A e C. Di conseguenza, la particella dell'LP viene convertita in un residuo CM più piccolo, arricchito in colesterolo, in quali apoB e apoE fungono da apoLP principale. I componenti residui risultanti del CM hanno una densità idrata di VLDL e LDPP e normalmente vengono eliminati dal siero sanguigno dagli epatociti attraverso recettori specifici (recettori apoE). Pertanto, negli individui sani, si presentano forme intermedie di LP breve periodo vita e in condizioni fisiologiche si trovano in basse concentrazioni nel siero sanguigno. Tuttavia, con alcune forme di HLP, si verifica un accumulo di DILI nel siero sanguigno. Come risultato di queste trasformazioni metaboliche, l'HLP transitorio, che si verifica entro poche ore dall'ingestione, viene eliminato, dopodiché avviene l'utilizzo dei lipidi esogeni nel fegato. La composizione in acidi grassi dei lipidi che entrano nel fegato corrisponde ai lipidi esogeni. Le particelle di LP secrete dal fegato nel flusso sanguigno hanno una composizione di acidi grassi caratteristica del corpo umano.
Le VLDL si formano nel fegato, vengono secrete dagli epatociti nel flusso sanguigno e trasportano i TG endogeni. Le VLDL nascenti contengono solo apoB-100. Nel flusso sanguigno, le VLDL includono anche apoLP da HDL - apoC-P e apoE. Studi sul metabolismo hanno dimostrato che la radioattività inizialmente presente nella frazione VLDL finisce rapidamente nelle particelle lipidiche di densità LDL idratate. Nelle persone sane, quasi tutte le LDL si formano nel letto vascolare dalle VLDL. Durante il metabolismo dei farmaci ricchi di TG contenenti apoB, a causa dell'idrolisi dei farmaci TG con densità inferiore e grandi formati vengono convertiti in DILP più densi e più piccoli e poi in LDL. Alcuni componenti residui di VLDL e LDPP se ne vanno letto vascolare attraverso l'interazione con specifici recettori apoB/E sulla superficie delle cellule endoteliali. Nel processo di trasformazione metabolica di LDPP in LDL, oltre alla lipoproteina lipasi eparina-dipendente, prende parte un altro enzima lipolitico: la lipasi trigliceridica epatica. Questo enzima è in grado di agire non solo come idrolasi TG, ma anche come fosfolipasi.
LDL negli esseri umani contiene maggior parte colesterolo circolante e trasportarlo ai tessuti periferici per i processi di formazione della membrana e steroidogenesi. L'LDL interagisce con i recettori sulla membrana plasmatica delle cellule epatiche, delle ghiandole surrenali e dei tessuti periferici, inclusi le cellule staminali mesenchimali e i fibroblasti. Dopo l'interazione con il recettore, le LDL subiscono endocitosi e i componenti delle LDL vengono catabolizzati nei lisosomi. Come risultato di questi processi si formano classi separate di lipidi; le apoproteine vengono scomposte in amminoacidi.
La Lp(a) è una particella ricca di colesterolo e proteine simile all'LDL, contenente 2 molecole di apo(a) oltre a 1 molecola di apoB. Livelli aumentati di Lp(a) sono associati allo sviluppo precoce della malattia coronarica: il 50% dei pazienti con malattia coronarica aveva un livello di Lp(a) superiore a 40 mg/l rispetto al 17% nel gruppo di controllo. Un aumento della concentrazione della lipoproteina(a) nel sangue è considerato un fattore di rischio indipendente per l'aterosclerosi.
L'HDL si forma in diversi modi, inclusa la sintesi e la secrezione di HDL nascente da parte del fegato e dell'intestino, durante la lipolisi delle lipoproteine ricche di TG o come risultato dell'interazione delle apoproteine neoformate con le membrane cellulari. Le HDL nascenti sono dischi a doppio strato costituiti da apoLP e fosfolipidi. Il colesterolo libero si diffonde nell’HDL dalle cellule endoteliali, dai globuli rossi e da altre cellule. Inoltre, durante la lipolisi delle VLDL o del CM, i lipidi superficiali (fosfolipidi e colesterolo) e le proteine (apoA-1, apoA-P, apoproteine del gruppo C) vengono trasferiti alla frazione HDL. Questi componenti possono formare particelle HDL discoidali nascenti o essere incorporati in particelle HDL preesistenti. L'HDL nascente discoidale nel plasma sanguigno interagisce con l'enzima LCAT, provocando la formazione di esteri del colesterolo, che si spostano rapidamente nel nucleo delle particelle; si formano particelle HDL sferiche (mature) - HDL3.
Negli esseri umani, la maggior parte degli esteri del colesterolo formati nelle HDL vengono trasferiti ai lipidi più grandi ricchi di TG. Proteina,
il trasferimento degli esteri del colesterolo, promuove lo scambio di esteri del colesterolo da HDL a TG da lipidi contenenti apoB, che porta ad una diminuzione del contenuto di esteri del colesterolo e ad un aumento del contenuto di TG nell'HDL. L'effetto combinato della lipolisi e del trasferimento dei lipidi determina l'arricchimento delle particelle HDL con ulteriori lipidi e apoproteine, con conseguente aumento delle dimensioni e diminuzione della densità delle particelle HDL, accompagnato dalla conversione delle particelle HDL3 in HDLg-Le particelle HDL2 sono quindi esposti alla trigliceride lipasi epatica, un enzima presente principalmente nelle cellule endoteliali dei sinusoidi epatici. La lipasi dei trigliceridi epatici ha sia attività di trigliceridi idrolasi che di fosfolipasi, che porta ad una diminuzione del contenuto di TG e fosfolipidi nell'HDL e ad una diminuzione della dimensione delle particelle.
Pertanto, i farmaci plasmatici, tradizionalmente suddivisi in classi mediante metodi fisici, chimici e immunochimici, costituiscono metabolicamente un unico sistema.
Il contenuto delle classi di farmaci nel siero del sangue può essere misurato quantitativamente. Tali misurazioni vengono effettuate mediante ultracentrifugazione analitica, che richiede sofisticate apparecchiature analitiche. È anche possibile calcolare la concentrazione del farmaco in base all'analisi di ciascun componente. Questo approccio richiede l'isolamento quantitativo e la purificazione di ciascuna classe di farmaci mediante ultracentrifugazione preparativa. Il livello dei farmaci plasmatici nella biochimica clinica viene solitamente valutato in base al loro contenuto di colesterolo. Il contenuto di TG nelle singole classi di farmaci, di regola, non viene studiato, poiché è soggetto a fluttuazioni più significative rispetto ai livelli di colesterolo. Va sottolineato che la maggior parte dei dati epidemiologici sulla connessione tra i farmaci e lo sviluppo di malattie cardiovascolari si basa sulla misurazione del contenuto di colesterolo nelle singole classi di farmaci.
Rapporto colesterolo totale il plasma sanguigno e il colesterolo delle principali classi di farmaci possono essere espressi con la seguente formula:
colesterolo nel sangue = colesterolo VLDL +
+ Colesterolo LDL (- Colesterolo HDL.
Per misurare le classi di colesterolo dei farmaci, determinare il colesterolo totale e i TG nel siero del sangue e la concentrazione di colesterolo HDL nel surnatante dopo la precipitazione dei farmaci contenenti apoB (VLDL e LDL). Concentrazione colesterolo LDL calcolato utilizzando la formula di Friedwald:
Colesterolo LDL = colesterolo totale -
- Colesterolo HDL - Colesterolo VLDL.
Per una valutazione approssimativa della concentrazione di colesterolo VLDL si utilizza il valore TG/5 se le concentrazioni di colesterolo e TG sono espresse in mg/dL, oppure TG/2,2 se le concentrazioni di colesterolo e TG sono espresse in mmol/l. L'utilizzo di questa formula a concentrazioni di TG superiori a 400 mg/dl porta ad una sovrastima del contenuto di colesterolo VLDL e ad una sottostima del contenuto di colesterolo LDL. Per valutare la distribuzione di LP in pazienti con livelli di TG molto elevati e presenza di una quantità significativa di CM, è necessaria un'ultracentrifugazione preliminare per separare CM e VLDL. Dopo la separazione della frazione VLDL e la precipitazione delle LDL nel surnatante, è possibile determinare la concentrazione del colesterolo HDL.
Un metodo affidabile per valutare il contenuto di singole classi di farmaci nel siero del sangue è il metodo EF su uno dei supporti di supporto: gel di agarosio, gel di poliacrilammide, acetato di cellulosa, carta. Questi metodi forniscono solo la percentuale delle singole classi di farmaci, inclusi HM, |3-, pre-f- e a-LP. L'EF è la base per la fenotipizzazione dell'HLP secondo la classificazione proposta da Fredrickson (Tabella 1.10). La decisione finale sul fenotipo HLP viene presa sulla base dei dati di LP EF, tenendo conto della concentrazione di colesterolo e TG nel sangue, nonché del colesterolo LDL.
Significato clinico della determinazione delle lipoproteine nel sangue
Numerosi studi clinici ed epidemiologici hanno stabilito che l’aumento dei livelli di colesterolo nel sangue è uno dei principali fattori di rischio per l’aterosclerosi. Una volta-
Tabella 1.10. Classificazione delle iperlipoproteinemie adottata dall'OMS Tipo CM LPD LDL Colesterolo TG Carattere
violazioni I t N N N TT T hm Pa - N TT TT N t lpn Pp - t t t t T lpn e lpd III - Floating r-LP t t t ACC e DILI IV - t N N (T) t T lpn V t 1 N N (T) tt Nota di T hmm e lponp. | - promozione; OKH - componenti residui di composti chimici.
Lo sviluppo dell'HLP può essere causato da anomalie genetiche e fattori ambientali (HLP primario), nonché da malattie come diabete, patologia del fegato, dei reni, disturbi ormonali(BPL secondario). La predisposizione ereditaria gioca un ruolo importante nella formazione dell'HLP primario. Nella popolazione bianca americana, il 63% della variabilità dei livelli di colesterolo è dovuta alla variazione genetica. Secondo uno studio sui gemelli mono e dizigoti in Russia, l’82% della variabilità del colesterolo totale è dovuta a fattori genetici.
Attualmente sono state studiate molte anomalie ereditarie del metabolismo lipidico, ma solo per alcune sono noti gli esatti difetti biochimici che consentono di diagnosticare la malattia. Queste forme di HLP comprendono la disbetalipoproteinemia familiare (HLP di tipo III) e l'iperchilomicronemia familiare (HLP di tipo I).
IPERCOLESTEROLEMIA FAMILIARE
Tra i pazienti con ipercolesterolemia primaria, i disturbi del metabolismo lipidico nell'ipercolesterolemia familiare sono i più studiati. Questa è una malattia monogenica ereditaria in cui alto livello siero e colesterolo LDL, nonché sviluppo iniziale aterosclerosi e cardiopatia ischemica.
Eredità dell'anca ipercolesterolemia familiare- autosomico dominante. Negli omozigoti la malattia è più grave che negli eterozigoti. Nel 60% degli omozigoti, l'IHD si sviluppa prima dei 10 anni. Il livello di colesterolo sierico negli eterozigoti varia da 250 a 550 mg/dl; negli omozigoti è stato osservato un aumento fino a 800 mg/dl. Gli xantomi tendinei sono considerati patognomonici per i pazienti con ipercolesterolemia familiare. L'incidenza dell'ipercolesterolemia familiare tra le popolazioni europee e americane è di 1: 1.000.000 per gli omozigoti e 1: 500 per gli eterozigoti, sebbene esistano popolazioni con una maggiore incidenza della malattia - 1: 100 (canadesi francesi, finlandesi, caucasici in Sud Africa).
Il difetto molecolare che determina lo sviluppo dell'ipercolesterolemia familiare è stato descritto da Brown e Goldstein. La causa dell'ipercolesterolemia è un difetto del recettore LDL, che provoca una forte diminuzione dell'assorbimento delle LDL, con conseguente accumulo di particelle LDL e, di conseguenza, un aumento della loro concentrazione nel sangue. Il grado di riduzione dell’attività del recettore LDL nei pazienti con ipercolesterolemia familiare è correlato ai livelli di colesterolo LDL, ma non alle manifestazioni cliniche della malattia. manifestazioni precoci IHD, presenza di xantomi tendinei). Esistono 4 tipi di difetti genetici nel recettore LDL:
completa assenza di proteine recettrici;
interruzione del trasporto delle proteine del recettore sulla superficie cellulare;
un difetto del recettore che impedisce il legame delle LDL;
un difetto nel recettore che ne impedisce l'internalizzazione dopo il legame con le LDL. Attualmente sono state identificate più di 150 mutazioni di questa proteina.
Nonostante l'instaurarsi di un difetto genetico che determina lo sviluppo di ipercolesterolemia familiare, le caratteristiche manifestazioni cliniche malattie e disturbi del metabolismo lipidico, i criteri per la diagnosi di ipercolesterolemia familiare non sono stati completamente determinati. Sfortunatamente, la determinazione dell’attività del recettore LDL per la diagnosi dell’ipercolesterolemia familiare non è ampiamente utilizzata. Si ritiene che l'uso del test del DNA per diagnosticare l'ipercolesterolemia familiare sia inappropriato a causa di grande quantità mutazioni. Aumento dei livelli di colesterolo - sfocato criterio diagnostico ipercolesterolemia familiare, poiché sono stati descritti pazienti con ridotta attività dei recettori apoB e livelli di colesterolo normali.
Una diminuzione del legame delle particelle LDL al recettore può essere dovuta non solo a difetti genetici che causano l'assenza del recettore LDL o al suo deterioramento funzionale, ma anche a cambiamenti ereditari nel ligando, apoB-100. L'analisi del DNA di pazienti con ipercolesterolemia ha rivelato una mutazione nel gene che codifica per la sintesi di apoB-100, a seguito della quale l'arginina in posizione 3500 viene sostituita dalla glutammina. Questa patologia è chiamata “difetto ereditario di apoB-100”. La frequenza di distribuzione di questo difetto nella popolazione è paragonabile a quella dell'ipercolesterolemia familiare (1: 500), secondo altri dati è significativamente inferiore (1: 1300).
IPERLIPIDEMIA COMBINATA FAMILIARE Il termine “iperlipidemia combinata familiare” (FCHL) è stato proposto per la prima volta da Goldstein et al. nel 1973 per caratterizzare un disordine ereditario del metabolismo lipidico, in cui si notano il probando e i suoi parenti di primo grado di parentela tipi diversi GLP (Na, Hn, IV). I disturbi del metabolismo dei lipidi compaiono solitamente dopo i 20 anni, ma possono essere rilevati in età più avanzata. gioventù. L'incidenza della SCHF nella popolazione è, secondo diversi ricercatori, dallo 0,5 al 2%, mentre tra i pazienti con infarto miocardico è del 5%, e tra i pazienti di età inferiore ai 60 anni con malattia coronarica documentata angiograficamente è di 15-20 %. Lo studio dei pedigree ha mostrato che SKGL - malattia ereditaria con ereditarietà di tipo autosomico dominante. Nonostante numerosi studi, il marcatore biochimico della SCHF non è stato identificato; L’esatto difetto molecolare che porta alla formazione di SCGL è ancora sconosciuto.
IPERCOLESTEROLEMIA POLIGENICA Tra i disturbi ereditari del metabolismo lipidico, l'ipercolesterolemia poligenica è la più comune. In questa malattia, la distribuzione dei livelli di colesterolo nei membri di tale
le famiglie vengono spostate verso qualcosa di più valori elevati rispetto alla media della popolazione. Inoltre, la distribuzione è unimodale, indicando l'assenza di eredità monogenica. Si ritiene che la presenza di ipercolesterolemia poligenica sia dovuta all'influenza cumulativa di diversi geni, mentre la manifestazione dell'ipercolesterolemia poligenica è in gran parte provocata da fattori ambientali, in particolare dalla natura della nutrizione. Il termine “ipercolesterolemia poligenica” viene utilizzato per caratterizzare i pazienti con ipercolesterolemia ereditaria con un difetto genetico sconosciuto e in assenza di segni di ereditarietà monogenica.
Data la natura poligenica dell'eredità dell'ipercolesterolemia, il difetto genetico che definisce la malattia e il suo marcatore (o marcatori) biochimici sono sconosciuti. La prevalenza dell'ipercolesterolemia poligenica nella popolazione e tra i pazienti con malattia coronarica dipende dai valori dei livelli di colesterolo accettati come normali: quando la norma diminuisce, la proporzione di pazienti con ipercolesterolemia poligenica aumenta naturalmente.
IPERTRIGLICERIDEMIA FAMILIARE
L'ipertrigliceridemia familiare è una malattia ereditaria del metabolismo lipidico, in cui il probando e i suoi parenti presentano un aumento moderato o significativo dei livelli di TG con concentrazioni normali di colesterolo LDL - HLP di tipo IV o V. La malattia di solito compare dopo i 30 anni. Questi pazienti spesso, ma non sempre, sperimentano una diminuzione dei livelli di colesterolo HDL. Il tipo di ereditarietà è autosomica dominante. L'ipertrigliceridemia familiare si verifica nella popolazione con una frequenza dell'1% e nei pazienti con malattia coronarica di età inferiore ai 60 anni con una frequenza del 5-8%. Il difetto genetico che causa lo sviluppo dell'ipertrigliceridemia familiare e il marcatore biochimico di questa malattia sono attualmente sconosciuti.
Criteri diagnostici per l'iperlipoproteinemia
Per stabilire l’HLP sono necessari criteri biochimici chiari. Attualmente esistono due approcci a questo problema. Per molti anni, i livelli normali di colesterolo sierico e TG, nonché di colesterolo LDL e HDL, sono stati stabiliti sulla base dei risultati di studi epidemiologici. Per ciascuna popolazione, sono stati determinati i propri criteri per la norma degli indicatori del metabolismo lipidico in base al 5-10% dei valori minimo e massimo con una distribuzione gaussiana. In questo caso, l'iperlipidemia è stata considerata come una concentrazione di colesterolo e TG che superava il livello del 90 o 95% su un diagramma gaussiano (a seconda del sesso e dell'età). Sono questi criteri per i valori normali che costituiscono la base per la classificazione della GLP proposta da Fredrickson et al. e dopo alcune integrazioni adottate dall'OMS. Successivamente, tenendo conto dei dati accumulati sulla relazione livello più alto Colesterolo e rischio di malattia coronarica, gruppi di esperti della Nazionale programma educativo sul colesterolo negli USA e nella Società Europea per lo Studio dell'Aterosclerosi è stata proposta una diversa classificazione della GLP. A seconda del rischio di malattia coronarica, si propone di assegnare il livello di colesterolo nel sangue desiderato rispettivamente a 6,2 o > 6,5 mmol/l per uomini e donne di età superiore ai 20 anni. Sulla base di questi criteri si propone di distinguere tra ipercolesterolemia moderata (6,2-7,5 o 6,5-7,8 mmol/l) e ipercolesterolemia grave (> 7,5 o > 7,8 mmol/l).
Di conseguenza, i criteri per il colesterolo LDL sono stati modificati, mentre il livello normale per il colesterolo TG e HDL è rimasto lo stesso. Sebbene per la popolazione americana l'ipercolesterolemia moderata corrisponda ad un livello del 75-95% della distribuzione gaussiana, questi criteri vengono utilizzati per la tipizzazione dell'HLP negli USA e in Russia. La scelta di valori normali più bassi è senza dubbio giustificata dal punto di vista della prevenzione primaria e secondaria della malattia coronarica. Inoltre, l'utilizzo di criteri diagnostici uniformi consente di confrontare la prevalenza dell'HLP in popolazioni con diversi livelli medi di colesterolo. D'altro canto, il 95% delle persone affette da HLP, accertato secondo criteri epidemiologici, hanno un HLP ereditario. Di conseguenza, man mano che diminuisci valori normali aumenterà la percentuale di persone il cui SLP è causato solo da fattori ambientali. A questo proposito, quando si ricercano pazienti con forme ereditarie gravi di ipercolesterolemia
il livello di colesterolo del 90-95% stabilito epidemiologicamente viene ancora utilizzato in base al sesso e all'età
paziente.
Ipolipoproteinemia
Le ipolipoproteinemie si verificano a causa di alcuni difetti negli enzimi che regolano il metabolismo dei lipidi o di mutazioni nel gene apoB.
L'abetalipoproteinemia e l'ipobetalipoproteinemia sono malattie ereditarie causate da mutazioni nel gene apoLP B, i principali apoLP XM e VLDL. Non è stata sviluppata una terapia specifica per questi disturbi.
Abetalipoproteinemia: nel sangue non sono presenti lipoproteine p, CM, lipoproteine con densità inferiore a 1,063 (LDL e VLDL), i globuli rossi presentano molteplici proiezioni a forma di punta (acantociti), si sviluppa una carenza di vitamine E e A (il risultato dell’assenza di LDL che trasportano le vitamine liposolubili) . Il malassorbimento intestinale è comune, così come i disturbi di incoordinazione (dovuti alla demielinizzazione assonale), l'atassia, il nistagmo, la degenerazione pigmentaria retinica e il ritardo mentale. La steatorrea si verifica a causa della mancanza di apoB, che determina la formazione di colesterolo nelle cellule intestinali. Una biopsia dell'intestino tenue rivela inclusioni di grasso nelle cellule epiteliali.
L'ipobetalipoproteinemia è accompagnata da una diminuzione del contenuto di p-lipoproteine ed è spesso asintomatica. Il contenuto di HDL è normale o aumentato, trigliceridi 70-120 mg%, LDL 20-70 mg%. L'assorbimento dei grassi solitamente non è compromesso.
TRIGLICERIDI
I trigliceridi (TG; triacilgliceroli) sono esteri dell'alcol trivalente glicerolo e degli acidi grassi a catena lunga, tra cui nel corpo umano dominano gli acidi stearico, palmitico, palmitooleico, oleico e linoleico. Meno comunemente sono presenti gli acidi linolenico e arachidonico polinsaturi. Le cellule dei tessuti umani non sintetizzano l'acido linoleico, quindi è classificato come acido essenziale. La distribuzione degli acidi grassi nella molecola TG dipende dalla lunghezza della catena e dal grado di insaturi™. La posizione centrale è spesso occupata dall'acido con la catena più lunga e il numero più grande doppi legami, le posizioni estreme sono solitamente occupate da acidi saturi.
Gli acidi grassi entrano nel corpo con il cibo sotto forma di triacilgliceroli, che intestino tenue sotto l'influenza della lipasi subiscono idrolisi. Il glicerolo e gli acidi grassi liberi risultanti vengono assorbiti dalle cellule epiteliali, dove avviene la risintesi dei TG. La formazione dei TG avviene nel fegato; gli acidi grassi entrano nel fegato dal tessuto adiposo o sono sintetizzati negli epatociti. La composizione degli acidi grassi dei TG nel corpo, in misura maggiore rispetto ai fosfolipidi, dipende dalla composizione dei grassi alimentari. Gli acidi grassi a catena lunga, che fanno parte dei TG, sono la fonte ottimale di composti ad alta energia formati durante la β-ossidazione degli acidi grassi nei mitocondri. La maggior parte delle autorità implementa il TG per coprire il fabbisogno energetico. L'eccezione è il cervello; il suo approvvigionamento energetico avviene attraverso il metabolismo del glucosio (glicolisi aerobica).
Nel plasma sanguigno, i TG, come altri lipidi (colesterolo e suoi esteri, fosfolipidi), vengono trasportati come parte dei complessi micellari macromolecolari - LP. I TG sono presenti in tutte le classi di farmaci; nelle VLDL e nei CM costituiscono la maggior parte dei lipidi. Con una certa esperienza aspetto il plasma consente di determinare approssimativamente il contenuto di TG in esso. Se il plasma sanguigno è trasparente, il livello di TG in esso contenuto non supera 180-200 mg%; L'opalescenza pronunciata del plasma sanguigno è caratteristica dell'ipertrigliceridemia dell'ordine di 300-500 mg%; con un livello di TG superiore a 600 mg%, il plasma e il siero sono opachi. La presenza di CM nel sangue può essere rilevata eseguendo un "test di permanenza" - l'aspetto di una "crema" sulla superficie del plasma quando viene conservato per 16 ore ad una temperatura di 4 °C. Il siero del sangue trasparente e la presenza di CM indicano il tipo I di HLP, l'assenza di CM con opalescenza pronunciata o colore lattiginoso del siero indica il tipo IV di HLP; la presenza di CM sulla superficie di un campione di siero con marcata opalescenza o colore lattiginoso suggerisce che il paziente è affetto da HLP di tipo III o V.
Significato clinico della determinazione della concentrazione di TG
Va ricordato che i CM possono essere presenti nel siero sanguigno di una persona sana che ha mangiato cibo meno di 12 ore prima del prelievo del sangue. A causa dell'influenza significativa dell'induzione del substrato sui livelli di TG, il livello di TG nel sangue può variare entro limiti significativi durante il giorno. L’ipertrigliceridemia può essere caratterizzata come fisiologica o patologica. L'ipertrigliceridemia fisiologica si manifesta dopo i pasti e può durare 12-14 ore; il grado della sua manifestazione dipende dalla natura e dalla quantità di cibo assunto. Ipertrigliceridemia in N-III trimestre anche la gravidanza è fisiologica. L'ipertrigliceridemia patologica, in base alle sue caratteristiche patogenetiche, può essere differenziata in primaria e secondaria. L'ipertrigliceridemia primaria è causata da disturbi genetici del metabolismo lipidico o da una maggiore induzione del substrato (eccesso di cibo). I disturbi del metabolismo dei TG che si verificano come complicazione del processo patologico principale sono classificati come ipertrigliceridemia secondaria.
Sono note diverse forme primarie di ipertrigliceridemia causate da difetti ereditari nel metabolismo dei farmaci: ipertrigliceridemia familiare, iperlipidemia familiare combinata, disbetalipoproteinemia familiare (HLP di tipo III), iperchilomicronemia familiare (HLP di tipo I). Anche l'ipotrigliceridemia causata da alterazioni genetiche nella sintesi o nella secrezione di apoB può essere classificata come forma primaria di disturbo del metabolismo dei TG. In queste malattie (abetalipoproteinemia ereditaria o ipobetalipoproteinemia omozigote), il contenuto di trigliceridi può essere ridotto al 19-20 mg%.
Lo sviluppo di ipertrigliceridemia secondaria è stato notato nel diabete mellito, nell'ipotiroidismo, nella sindrome nefrosica, nell'assunzione di contraccettivi orali e nel trattamento con glucocorticoidi. Generalmente, trattamento patogenetico, rivolto al principale processo patologico, è accompagnato da una diminuzione dei livelli di TG.
La correzione dei livelli glicemici con insulina o farmaci ipoglicemizzanti orali aiuta a normalizzare i livelli di TG nel sangue. Quando la sindrome nefrosica viene alleviata, anche il livello di TG nel sangue diminuisce, ma a volte rimane elevato per lungo tempo. Anche la sospensione dell’assunzione di contraccettivi ormonali, glucocorticoidi e di una serie di altri farmaci (β-bloccanti) aiuta a normalizzare i livelli di TG.
L'ipertrigliceridemia si nota con infiltrazione grassa del fegato. La patogenesi di questa malattia si basa da un lato su un aumento dell'apporto di acidi grassi liberi al fegato (aumento della lipolisi) o sulla loro aumentata sintesi (risintesi) e, dall'altro, su una violazione della secrezione di VLDL da parte del fegato . Molti composti tossici, alcol, preparazioni farmacologiche contribuiscono all'interruzione della sintesi di apoLP, in particolare di apoB, in assenza o carenza della quale non avviene la formazione di VLDL. In queste condizioni, i TG provenienti dal sangue o appena sintetizzati finiscono nel fegato, formando la struttura del “fegato di maiale”, provocandone l’infiltrazione di grassi e la cirrosi.
L'ipertrigliceridemia si osserva solitamente nei pazienti con pancreatite acuta. Tuttavia, un aumento del livello di TG non è tipico di un danno acuto al pancreas; al contrario, una grave ipertrigliceridemia (>1000 mg%), sia primaria che secondaria, contribuisce allo sviluppo di malattie del pancreas e spesso è complicata da pancreatite. La pancreatite è considerata una delle manifestazioni cliniche delle forme familiari di ipertrigliceridemia. Lo sviluppo della pancreatite è spesso osservato nei pazienti con diabete, alcolismo e donne che assumono contraccettivi ormonali.
I dati sulla relazione tra ipertrigliceridemia e malattia coronarica sono contraddittori, sebbene studi epidemiologici condotti in molte popolazioni abbiano dimostrato l’indipendenza dei trigliceridi come fattore di rischio per la malattia coronarica. Valutare l’indipendenza dei TG come fattore di rischio per la malattia coronarica è estremamente difficile a causa della presenza di una relazione negativa tra il livello dei TG e il colesterolo HDL, indipendente fattore negativo rischio di cardiopatia ischemica. Tuttavia, se un aumento della concentrazione di TG non è un fattore di rischio indipendente per l’IHD, la stretta relazione tra il metabolismo dei TG e quello delle HDL non fornisce motivo per negare l’importanza dell’ipertrigliceridemia nella genesi dell’IHD. Basi biochimiche Ciò potrebbe essere dovuto all'accumulo di DILI nel sangue, che ha una pronunciata aterogenicità.
Il significato dell'ipertrigliceridemia nella formazione della patologia dei vasi periferici e cerebrali, in contrasto con la cardiopatia ischemica, è più definito. Studi condotti da specialisti giapponesi hanno dimostrato che in questa popolazione, con bassi livelli di colesterolo nel sangue e incidenza di infarto miocardico, l'ipertrigliceridemia rappresenta un fattore di rischio per la patologia delle arterie periferiche. Dati simili sono stati ottenuti in altre popolazioni.
Lo studio del contenuto di TG nel siero del sangue ha molto in comune con la determinazione del colesterolo. Sia il colesterolo che i TG - componenti strutturali Farmaci che devono essere distrutti per rendere i lipidi disponibili per reazioni chimiche o enzimatiche. Entrambi i composti sono idrofobici e hanno una bassa solubilità in soluzione acquosa. Tuttavia, ci sono anche differenze significative nella determinazione del contenuto di TG nel siero sanguigno: per valutare il contenuto di TG, viene determinato il glicerolo rilasciato dopo l'idrolisi dei TG.
Metodi per determinare il contenuto di TG nel sangue
Esistono due modi per determinare il contenuto di TG nel siero del sangue: chimico ed enzimatico; ciascuno di essi comprende numerose tecniche metodologiche. Attualmente la determinazione dei TG è possibile anche utilizzando il metodo della “chimica secca”.
Metodi chimici. Per determinare la TG, i metodi chimici diretti (senza estrazione preliminare con un solvente organico) sono inaccettabili, poiché il siero del sangue contiene molti composti che, durante il processo di idrolisi chimica (saponificazione degli acidi grassi), possono anche essere convertiti in glicerolo, come i fosfolipidi e glucosio. A questo proposito, nell metodi chimici determinazione dei TG, la prima fase è l'estrazione dei TG e la rimozione dei composti interferenti. Per l'estrazione vengono utilizzati solventi organici: metanolo, etanolo, isopropanolo e cloroformio. I solventi denaturano il farmaco e promuovono la dissociazione dei TG. I composti interferenti vengono rimossi mediante estrazione con solventi organici (esano, nonano) o utilizzando adsorbenti (zeolite, acido silicico, florisil).
La seconda fase della determinazione chimica dei TG - idrolisi dei TG con formazione di glicerolo e acidi grassi liberi - viene solitamente effettuata mediante reazione con una soluzione etanolica di idrossido di potassio a temperatura elevata (saponificazione).
Dopo che il glicerolo è stato ossidato in formaldeide, viene determinato da una delle seguenti reazioni:
reazione della formaldeide con una miscela di acidi cromotropici e solforici (misurazione a 570 nm);
reazione con una miscela di fenilidrazina, cianuro ferrico e acido cloridrico (misurazione a 540 nm);
reazione con una miscela di 3-metil-2-benzotiazolina e cloruro ferrico (misurazione a 620 nm);
reazione con acetato di ammonio e acetilacetone.
Il prodotto di reazione può essere misurato colorimetricamente a
412 nm o fluorimetrico. Il metodo fluorimetrico è più comune ed è adatto agli analizzatori semiautomatici.
Metodi enzimatici. Nei metodi enzimatici per la determinazione dei TG, la distruzione dei complessi lipidici e l'idrolisi dei TG vengono effettuate in presenza di detergenti, lipasi e proteasi. In condizioni di idrolisi enzimatica dei TG, il glicerolo non viene rilasciato dai fosfolipidi e dal glucosio, pertanto la specificità della determinazione aumenta.
Per determinare il glicerolo formato durante l'idrolisi dei TG vengono utilizzate numerose tecniche metodologiche, utilizzando in ciascun caso una serie di reazioni enzimatiche accoppiate. I metodi enzimatici consentono la determinazione dei TG mediante diminuzione o aumento della concentrazione di NADH ad una lunghezza d'onda di 340 nm. Sono stati proposti anche metodi colorimetrici: riduzione dei sali nitroblu di tetrazolio utilizzando NADH e misurazione del formazan formato (lunghezza d'onda 505 nm) o ossidazione del 4-amminofenazone in presenza di perossidasi per formare un cromogeno (lunghezza d'onda 510 nm).
L'utilizzo del metodo enzimatico evita l'uso di solventi organici, la purificazione degli estratti per rimuovere i composti interferenti, l'uso di idrossido di potassio concentrato e l'idrolisi a alta temperatura. Rispetto ai metodi chimici, la determinazione enzimatica dei TG ha una specificità e una precisione più elevate, il che è conveniente per l'automazione.
Le principali difficoltà della determinazione enzimatica dei TG sono la possibilità di distruzione incompleta del complesso farmacologico e di idrolisi incompleta con rilascio di glicerolo libero. È noto da studi sperimentali che i TG formati da acidi grassi saturi e insaturi vengono idrolizzati a velocità diverse per azione della lipasi; Anche il rapporto tra acidi grassi a catena corta e lunga gioca un ruolo nella velocità di idrolisi.
Poiché la determinazione del contenuto di TG nel siero sanguigno, sia con metodi chimici che enzimatici, si basa sul rilascio di glicerolo, è necessario tenere conto del contenuto di glicerolo libero nel siero sanguigno. Per la maggior parte dei campioni di siero fresco con livelli di TG inferiori a 700 mg%, le variazioni del glicerolo libero non influiscono in modo significativo sui risultati ottenuti. L'accumulo di glicerolo libero nel siero del sangue si verifica nel diabete mellito, in alcuni tipi di patologie epatiche, in condizioni di emodialisi, in malattie acute e croniche insufficienza renale, assunzione di nitroglicerina, terapia con eparina. Infezioni acute, stati di stress a breve termine, assunzione medicinali, che hanno proprietà lipolitiche, aumentano significativamente il contenuto di glicerolo libero nel sangue. L'idrolisi spontanea dei TG si verifica anche nel siero sanguigno, soprattutto quando i campioni vengono conservati a temperatura ambiente.
Per determinare con precisione il livello di TG, è necessario misurare il livello di glicerolo libero e sottrarre il risultato significato generale. Quando si determina il livello di glicerolo libero mediante metodi colorimetrici e fluorimetrici, può essere diverso nello stesso campione di plasma sanguigno. La fluorometria Fornisce risultati più stabili, ma è meno specifica: in condizioni di insufficienza renale e proteolisi attivata, i peptidi fluorescenti possono gonfiare significativamente i risultati.
Per ottenere dati accurati durante la determinazione del TG, è necessario utilizzare norma primaria- una soluzione di triacilgliceroli in solventi organici. La trioleina viene solitamente utilizzata come standard, sebbene una miscela di trioleina e tripalmitina (2:1 in peso) sia più simile nel grado di insaturazione degli acidi grassi ai TG umani. Di norma, una soluzione di trioleina in isopropanolo può essere utilizzata solo per metodi di ricerca chimica, poiché l'isopropanolo attiva in modo non specifico gli enzimi utilizzati per determinare la TG. A questo proposito, quando si determinano i TG con metodi enzimatici, vengono utilizzati standard secondari (calibratori): siero di sangue umano, il cui contenuto di TG è determinato con metodi di riferimento utilizzando standard primari.
Il prelievo di sangue per determinare i TG deve essere effettuato 12-14 ore dopo il pasto. I valori di TG determinati nel plasma sanguigno sono inferiori del 2-4% rispetto a quelli nel siero. Ciò è dovuto alla diluizione dovuta al fluido perso dai globuli rossi sotto l'influenza dell'anticoagulante. Alcuni anticoagulanti (fluoro, citrato e ossalato) causano una maggiore perdita di liquidi dai globuli rossi, che si riflette in una maggiore diminuzione dei valori TG (fino al 10%). È preferibile utilizzare l'EDTA (1 mg/ml di sangue intero) come anticoagulante per la determinazione dei TG. Per ridurre l'effetto di diluizione, è necessario centrifugare i campioni di sangue e separare il plasma risultante entro 2 ore dal prelievo del sangue.
COLESTEROLO
Il colesterolo, 3-idrossicolest-5-ene, è uno sterolo monoinsaturo con la composizione C27H40(0H)b; Di struttura chimicaè un alcol secondario monovalente (colesterolo). La molecola del colesterolo si basa sulla struttura del ciclopentanoperidrofenantrene, nella terza posizione c'è un gruppo ossidrile, c'è un doppio legame tra il 5o e il 6o atomo di carbonio e una catena alifatica laterale di 8 atomi di carbonio. Il termine “colesterolo” definisce la fonte da cui fu isolato per la prima volta (dal greco cbo1e - bile). Massa molecolare colesterolo 387 D, cm forma puraè polvere bianco. Colesterolo
si dissolve bene nei solventi organici (particolarmente bene negli alcoli con 6-7 atomi di carbonio). La solubilità del colesterolo in acqua è estremamente bassa, 4,7 µmol/l a 25 °C.
La struttura chimica del colesterolo limita il numero di reazioni biochimiche in cui entra. La principale trasformazione biochimica del colesterolo (come alcol) è la formazione di un legame estere con gli acidi, in particolare con gli acidi grassi liberi, con formazione di esteri del colesterolo. L'80% del colesterolo nel corpo è colesterolo libero, quasi tutto fa parte delle membrane biologiche. Allo stesso tempo, 2/3 del colesterolo nel siero del sangue si presentano sotto forma di esteri con acidi grassi.
Il colesterolo fa parte di tutte le cellule viventi. Nelle membrane biologiche, il colesterolo svolge un ruolo strutturale. La sua funzione, che determina la permeabilità della membrana e la creazione di un microambiente per gli enzimi incorporati nella membrana, è indissolubilmente legata alla sua struttura. Caratteristica importante colesterolo - il suo ruolo di precursore per la sintesi degli ormoni steroidei.
Tutte le cellule del corpo sintetizzano il colesterolo dall'acetato, passando successivamente attraverso gli stadi di acetoacetato, metilglutaril-CoA e, infine, acido mevalonico. Conversione del metilglutaril-CoA in acido mevalonico mediante l'azione dell'idrossimetilglutaril-CoA (HMG-CoA) reduttasi - momento chiave sintesi del colesterolo. L'acido mevalonico risultante può essere utilizzato solo nella sintesi del colesterolo, quindi l'attivazione della HMG-CoA reduttasi porta sempre all'ipercolesterolemia.
La maggior parte del colesterolo è sintetizzato nel fegato e proviene dal cibo; Il corpo sintetizza 1 g di colesterolo al giorno. Una parte del colesterolo viene ossidata in acidi biliari e una parte viene eliminata con le feci. Esiste una relazione reciproca tra il colesterolo fornito con il cibo e quello sintetizzato dall'organismo. Il colesterolo, come altri lipidi, viene trasportato nel siero sanguigno come parte dei farmaci.
La maggior parte del colesterolo sierico contiene LDL; il loro apoLP principale è apoB-100. LDL è la principale forma di trasporto del colesterolo e degli acidi grassi polienici da
Biochimica clinica del fegato ai tessuti periferici. Le LDL, quando interagiscono con i recettori apoB sulla membrana plasmatica, subiscono internalizzazione; nei liposomi la distruzione della particella lipidica avviene con rilascio di colesterolo libero. La cellula utilizza il colesterolo per costruire le membrane e una parte di esso è esterificato. Molecole di colesterolo membrane plasmatiche, una volta nel siero del sangue, iniziano la via afferente al fegato come parte delle HDL. Il colesterolo che entra nel fegato come parte dell'HDL subisce idrossilazione microsomiale, trasformandosi in acidi biliari e viene anche escreto nella bile sotto forma di colesterolo libero o suoi esteri.
I livelli di colesterolo sono influenzati dalla natura del cibo e dall'assunzione di alcol, intensa allenamento fisico, farmaci farmacologici, compresi contraccettivi ormonali, steroidi, farmaci ipolipemizzanti. Le variazioni stagionali e diurne non hanno un effetto significativo sui livelli di colesterolo nel siero. Le fluttuazioni individuali dei valori del colesterolo vanno, secondo vari autori, dal 3 al 14%. Le fluttuazioni giornaliere dei livelli di colesterolo negli uomini e nelle donne (0,7-4,3%) sono significativamente inferiori alle fluttuazioni durante
mesi (2,0-12,2%) e 1 anno (3,9-10,9%).
Significato clinico dell'ipercolesterolemia
L’ipercolesterolemia è il fattore di rischio più documentato per l’aterosclerosi coronarica. Ciò è stato confermato da numerosi studi epidemiologici e studi clinici che hanno stabilito una connessione tra ipercolesterolemia e aterosclerosi coronarica, la frequenza delle manifestazioni cliniche della malattia coronarica (angina pectoris e infarto del miocardio). A prevenzione secondaria L'aterosclerosi coronarica ha mostrato una regressione durante la terapia ipolipemizzante quadro clinico IHD e cardiosclerosi con normalizzazione dei livelli di colesterolo sierico. Per una caratterizzazione più dettagliata dei fattori di rischio per l'aterosclerosi coronarica, viene studiato il contenuto di colesterolo nelle singole classi di farmaci; Lo studio più utilizzato è quello del colesterolo LDL e HDL, che sono rispettivamente fattori di rischio e anti-rischio affidabili per l’aterosclerosi coronarica.
Lo studio del colesterolo non fornisce informazioni diagnostiche riguardanti una malattia specifica, ma caratterizza la patologia dei lipidi e del metabolismo lipidico. La determinazione dei livelli di colesterolo è di importanza sociale, poiché l’aumento dei livelli di colesterolo nella popolazione richiede l’attuazione di misure organizzative prevenzione primaria aterosclerosi coronarica. I numeri più alti di ipercolesterolemia si osservano nei disturbi genetici del metabolismo lipidico: ipercolesterolemia familiare omo ed eterozigote, iperlipidemia familiare combinata, ipercolesterolemia poligenica.
Con un numero condizioni patologiche si sviluppa un'ipercolesterolemia secondaria. Tali processi includono ipotiroidismo, Sindrome nevrotica, diabete mellito, ecc. L'ipocolesterolemia si verifica con l'ipertiroidismo, in situazioni cliniche gravi che accompagnano varie forme nosologiche malattie, con fase acuta processo infiammatorio, così come il cancro.
Metodi per la determinazione del colesterolo sierico
I metodi per determinare il colesterolo sierico sono numerosi; Si possono distinguere metodi chimici ed enzimatici. A loro volta, tra i metodi chimici esistono metodi diretti e indiretti (estrazione).
METODI CHIMICI PER LA DETERMINAZIONE DELLA CONCENTRAZIONE DI COLESTEROLO
Metodi diretti. La base dei metodi diretti per determinare il colesterolo è la reazione di Liebermann-Burkhard, in cui il colesterolo reagisce con una miscela di acido solforico e acetico, nonché con anidride acetica. La reazione avviene in un mezzo anidro fortemente acido. Durante la reazione, la molecola di colesterolo viene successivamente ossidata e si forma il composto colorato colestagessaene - acido solforico con massimi di assorbimento a 410 e 610 nm. Il colore in via di sviluppo è instabile, quindi il tempo fotometrico deve essere mantenuto con precisione. In letteratura si possono trovare diversi rapporti degli ingredienti nel reagente di Liebermann-Burkhard; Maggiore è il contenuto di anidride acetica, più velocemente avviene la reazione. Reak
la reazione del colesterolo con la miscela Liebermann-Burkhard non è specifica, nella reazione definizione diretta colesterolo, il colore della miscela di reazione con la soluzione standard è smeraldo; I campioni di siero possono dare un colore verde, blu o marrone. Ciò è dovuto al fatto che molti componenti del siero sanguigno reagiscono. Inoltre, nella reazione di Liebermann-Burkhard, il colesterolo libero ed i suoi esteri formano complessi colorati con coefficienti di assorbimento molare diversi: nel caso degli esteri del colesterolo, la densità ottica è maggiore. Ciò introduce intrinsecamente un errore nello studio, poiché il rapporto colesterolo libero/esteri di colesterolo in ciascuno dei campioni non è noto.
La reazione Killiany-Zak è un altro metodo comune per determinare il colesterolo (la reazione del colesterolo con sali di ferro, acido acetico e solforico). Questo reazione chimica avviene in presenza di ioni ferro ferrico con la formazione sequenziale di 2, 3 e 4 doppi legami coniugati nella molecola del colesterolo, mentre il massimo di assorbimento dei composti risultanti varia da 420 a 563 nm. Nella reazione Killiany-Zak, il colesterolo libero e i suoi esteri danno complessi colorati simili.
Metodi indiretti. I metodi indiretti includono tecniche in cui i lipidi dal siero del sangue vengono prima estratti con solventi organici e, dopo l'evaporazione, viene eseguita la reazione di Liebermann-Burkhard. Questo metodo è più riproducibile e accurato, poiché è possibile rimuovere le sostanze interferenti rimaste nella fase acquosa. Per l'estrazione vengono utilizzati sistemi etanolo-dietiletere, etanolo-acetone, metanolo-cloroformio; i più comunemente usati sono l'esano e l'alcol isopropilico. L'inclusione di una fase di estrazione aumenta la specificità del metodo; i risultati della determinazione del colesterolo sono inferiori del 7% rispetto al metodo diretto.
I metodi di riferimento per determinare il colesterolo passano attraverso 3 fasi; questi includono l'estrazione dei lipidi in un solvente non polare, la saponificazione degli esteri del colesterolo e la riestrazione del colesterolo dalla miscela saponificata. Il metodo di riferimento maggiormente preferito è il metodo Abell-Kendall, proposto nel 1952: l'estratto lipidico viene saponificato con una soluzione alcolica di idrossido di potassio ed estratto con etere di petrolio. La reazione Liebermann-Burkhard viene effettuata dopo l'evaporazione dell'etere di petrolio.
Le difficoltà nella preparazione di soluzioni standard di colesterolo sono associate alla sua idrofobicità. A lungo Come soluzione standard è stata utilizzata una soluzione di colesterolo in acido acetico glaciale. Inoltre, come standard è stata proposta una soluzione di colesterolo in isopropanolo; tuttavia, tale standard si è rivelato scomodo per i metodi enzimatici per la determinazione del colesterolo, poiché l'isopropanolo attivava i sistemi enzimatici, aumentando la densità ottica. Per determinare il colesterolo quando si lavora con analizzatori biochimici, è generalmente accettato l'uso di standard secondari (calibratori): siero del sangue, il cui livello di colesterolo è determinato mediante metodi di riferimento.
Nonostante la relativa semplicità di implementazione e il basso costo della procedura, i metodi chimici sono tossici e il loro utilizzo sui moderni analizzatori è associato alla corrosione del sistema.
METODI ENZIMATIVI PER LA DETERMINAZIONE DELLA CONCENTRAZIONE
COLESTEROLO
Vantaggi della determinazione enzimatica: realizzazione della reazione in fase acquosa; facilità di automazione; elevata sensibilità e specificità.
La determinazione enzimatica del colesterolo comprende anche diverse fasi: idrolisi enzimatica degli esteri del colesterolo sotto l'azione della colesterolo esterasi con formazione di colesterolo libero e acidi grassi liberi; ossidazione del colesterolo con l'ossigeno disciolto nel mezzo di reazione sotto l'azione della colesterolo ossidasi con formazione di colest-4-en-3-olo e H2O2. Nei metodi più utilizzati, il perossido di idrogeno risultante ossida alcuni composti per formare cromogeni che possono essere misurati fotometricamente. Tali cromogeni possono includere 4-amminofenazone (assorbimento massimo 500-525 nm), nonché la formazione dell'eterociclo Henach nella reazione con metanolo (assorbimento massimo 405 nm). È possibile registrare il perossido di idrogeno formato nella reazione con deidrogenasi NAD-dipendenti e misurare la densità ottica ad una lunghezza d'onda di 340 nm.
Come nei metodi chimici, alcuni componenti del colesterolo sierico possono influenzare i risultati della determinazione. La bilirubina inibisce la reazione della perossidasi con 4-aminoantipirina e fenolo. Anche i riducenti dei gruppi sulfidrilici (glutatione, cisteina, ditiotreitolo) influenzano i risultati della reazione. Metodi basati su diverse reazioni enzimatiche possono dare risultati diversi.
Le difficoltà metodologiche nella determinazione enzimatica del colesterolo sono associate all'eterogeneità della distribuzione del colesterolo tra i farmaci, nonché alla sua esterificazione nel sangue. La completezza dell'idrolisi degli esteri del colesterolo dipende dalla fonte dell'enzima. La colesterolo esterasi microbica idrolizza più attivamente gli esteri del colesterolo formati da acidi grassi saturi, mentre la colesterolo esterasi pancreatica idrolizza gli acidi grassi polinsaturi.
Il prelievo del sangue per la determinazione di tutti i lipidi viene solitamente effettuato dopo 12-16 ore di digiuno. La concentrazione di colesterolo nel sangue non cambia prima e dopo un pasto, tuttavia, una marcata opalescenza del siero del sangue dovuta alla presenza di CM e VLDL può interferire con la determinazione. È consigliabile che il paziente non modifichi la sua dieta abituale per almeno 2 settimane prima del prelievo del sangue. Siero o plasma possono essere utilizzati per determinare i livelli di colesterolo. L'EDTA secco (1 mg/ml di sangue) viene solitamente utilizzato come anticoagulante per la produzione di plasma, che ha anche proprietà antiossidanti. La concentrazione di colesterolo nel siero del sangue è inferiore del 2-4% rispetto a quella del plasma sanguigno.
Lipidi– sostanze simili ai grassi che fanno parte di tutte le cellule viventi. Questo è un gruppo ampio composti organici, compresi gli acidi grassi, nonché i loro derivati - sia il gruppo radicale che quello carbossilico. I lipidi svolgono un ruolo molto importante nei processi vitali.
I lipidi svolgono un’ampia varietà di funzioni. Fanno parte delle membrane cellulari e servono come precursori degli ormoni steroidei, degli acidi biliari, delle prostaglandine e dei fosfoinositidi.
Il sangue contiene singoli componenti lipidici: acidi grassi saturi (SFA), acidi grassi monoinsaturi (MUFA) e acidi grassi polinsaturi (PUFA), trigliceridi, colesterolo, esteri del colesterolo e fosfolipidi.
I lipidi sono i componenti principali delle membrane biologiche, influenzano la permeabilità cellulare e l'attività di molti enzimi e sono coinvolti nella trasmissione di impulso nervoso, nella contrazione muscolare, nella creazione di contatti intercellulari, nei processi immunochimici.
I lipidi sono caratterizzati dalla loro solubilità nei solventi organici e, di regola, insolubili in acqua; si trovano in tutte le cellule viventi. I lipidi costituiscono una riserva energetica e creano coperture protettive idrorepellenti e termoisolanti negli animali e nelle piante, oltre a proteggere vari organi dallo stress meccanico. (basato su materiali di TSB, Wikipedia, SMT).
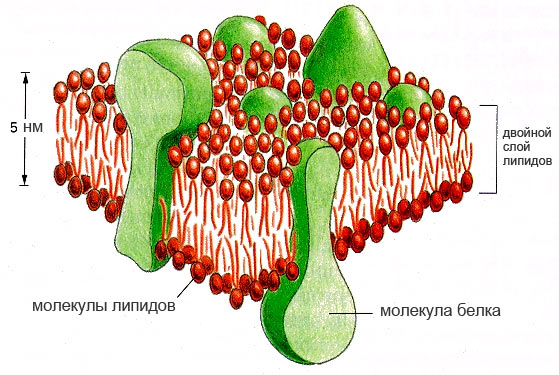
Membrana cellulare
Lipidi trasportato attraverso il flusso sanguigno a vari organi e tessuti come parte di complessi composti macromolecolari - lipoproteine , che sono composti di varie proteine (apoproteine) e lipidi.
Lipoproteine sono divisi in:
Le lipoproteine differiscono per dimensioni, composizione e ruolo nell'aterogenesi.
Nella pratica clinica valore più alto LDL e HDL svolgono un ruolo nello sviluppo della patologia vascolare.
LDL composto principalmente da esteri del colesterolo. La loro apoproteina funzionale è apoB-100.
Affinché le LDL diventino aterogene, devono subire una modificazione, che molto spesso è causata dal processo della loro perossidazione. LDL ossidato cambia le sue proprietà. Innanzitutto, la loro interazione con i recettori epatici viene interrotta, quindi diventano irritanti attivi per i monociti. I monociti del sangue attivati penetrano nello spazio subendoteliale del vaso, trasformandosi in macrofagi, che fagocitano le LDL modificate e si trasformano in cellule schiumose, ad es. cellule traboccanti di esteri del colesterolo. È proprio il colesterolo LDL, dato il suo importante ruolo nella formazione della placca aterosclerotica, il principale bersaglio della terapia ipolipemizzante.
HDL effettuare il trasporto inverso del colesterolo da parete vascolare e macrofagi al fegato, da dove vengono escreti dal corpo come parte degli acidi biliari. Esistono due sottoclassi di HDL: HDL-2 e HDL-3. È l'HDL-3 che cattura il colesterolo dalle cellule periferiche e dai macrofagi, trasformandosi in HDL-2, costituito da esteri del colesterolo e fosfolipidi. L'HDL si lega ai recettori del fegato e alle cellule della parete vascolare attraverso le apoproteine (apoA-1 e apoA-2).
Lipoproteine a bassa densità- Si tratta di composti di proteine con colesterolo che trasportano quest'ultimo dal fegato ai tessuti. L’LDL trasporta il colesterolo di cui ha bisogno ai tessuti, ma all’aumentare della loro concentrazione aumenta il rischio di sviluppare l’aterosclerosi.
Sotto aterogenicoè necessario comprendere le lipoproteine, la cui elevata concentrazione nel plasma sanguigno è associata in modo affidabile allo sviluppo dell'aterosclerosi.
Macrofagi- grandi cellule formate da monociti, capaci di fagocitosi - il processo di cattura attiva e assorbimento di particelle viventi e non viventi da parte di organismi unicellulari o cellule speciali (fagociti) di organismi animali multicellulari.
Letteratura
Diagnosi e correzione dei disturbi del metabolismo lipidico per la prevenzione e il trattamento dell'aterosclerosi. Raccomandazioni russe. Sviluppato dal comitato di esperti GFOC. M., 2004.
Diagnosi e correzione dei disturbi del metabolismo lipidico ai fini della prevenzione e del trattamento dell'aterosclerosi // Brevi raccomandazioni russe. Sviluppato da un gruppo di esperti VNOK. M., 2005.
Orlov R.S., Nozdrachev A.D. Fisiologia normale: Manuale. – M.: GEOTAR-Media, 2009.
Pogozheva A.V. Fondamenti di dietoterapia razionale per le malattie cardiovascolari // Dietetica clinica. 2004.T.1. N. 2.
Pogozheva A.V. Rappresentazioni moderne sulla dietoterapia per l'ipertensione arteriosa // Medicina. La qualità della vita. 2005. N. 3.
Dieta, nutrizione e prevenzione malattie croniche. CHI. Ginevra, 1993.
Riabilitazione per malattie del sistema cardiovascolare/Ed. IN. Makarova. M., 2010.

