5-10-2012, 19:14
Funkcie kôry
čelný lalok. Hlavnou funkciou predných lalokov je kontrola vôľových pohybov, koordinácia motorických mechanizmov expresívnej reči, ako aj poskytovanie „kreatívneho“ alebo abstraktného myslenia.
Predný lalok možno rozdeliť do piatich oblastí:
- motor,
- premotor,
- predný motorický vizuál,
- Brocova motorická oblasť reči
- a frontálnej asociačnej oblasti
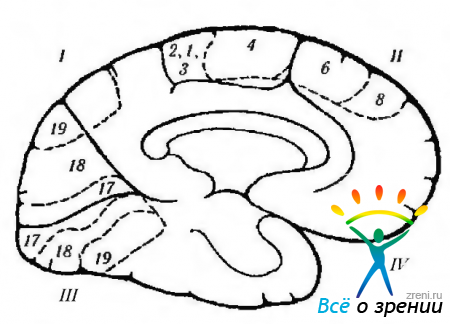
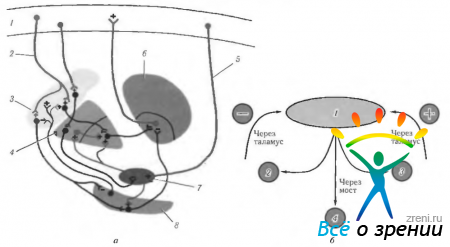
Ryža. 4.1.12. Schematické znázornenie funkcií kôry veľký mozog, lokalizované na mediálnom povrchu, a ich porušenie počas rôzne choroby. sprevádzané excitáciou alebo deštrukciou kôry (podľa Crosby et al., 1962): 1 - parietálny lalok. Centrálny gyrus (4). Vľavo okcipitálna oblasť: -alexia bez agrafie. Zadná kôra: -ťažkosti s prekladaním pohľadu; - Balintov syndróm (bilaterálny), 11 - čelný lalok. Prídavné motorické pole (6): + zdvihnutie opačnej ruky, otočenie očí a hlavy k ruke; + kontrakcia zrenice; + oneskorenie reči. Predné motorické očné pole (8). Pásový gyrus: + rozšírenie zrenice; + slzenie. orbitálny povrch. III - okcipitálny lalok. primárna zraková kôra. Kontralaterálne videnie: - pravostranná hemianopsia. IV-temporálny lalok: paralýza pohybov očí; ipsilaterálna hemiplégia; hydrocefalus; decerebrálna rigidita; stopková halucinóza; kontralaterálna hemianopsia; parinaudov syndróm. Pod povrchom: - rozpoznávanie tváre. Poznámka. Cytoarchitektonické polia zodpovedajú Brodmannovej klasifikácii. Plus označuje výsledok elektrickej stimulácie kôry a mínus - výsledok zničenia poľa
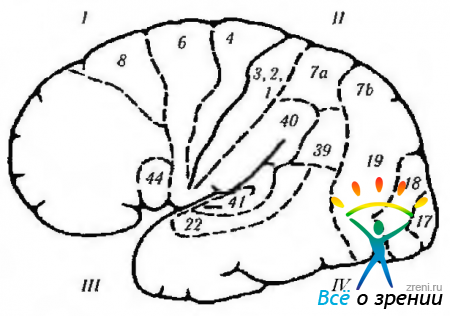
Ryža. 4.1.13. Bočný povrch mozgu (označenia sú podobné tým, ktoré sú uvedené na predchádzajúcom obrázku): 1 - predný lalok. Motorové pole (4). Premotorické pole (6). Motorické očné pole (8): + kontralaterálna sakáda; + zatvorenie opačného oka; + gomolaterálna priateľská deviácia oka. Motorické rečové pole (44): - motorická (motorická) afázia. 11 - parietálny lalok: - (- jednoduchá halucinóza; - ilúzie; - metamorfopsia: - strata pozornosti; - agnózia; - apraxia; - oslabenie optokinetického nystagmu; - hemianopia dolného kvadrantu. Supramarginálny gyrus (40): - amnézia; - žargón afázia. Angular gyrus (39): - agrafia; - akalkulia; - strata schopnosti rozlišovať medzi ľavou a pravou stranou pravá strana; agnózia prstov. somatickej citlivosti. III-temporálny lalok: + vizuálne halucinácie; + deja vu; + zhoršený pocit vône; + záchvaty; + automatizmy; - horná kvadrianopsia; -strata pamäti; - Klüver-Bucyho syndróm. primárna sluchová kôra. Sekundárna sluchová kôra: -sluchová afázia. Planum temporale: - dyslexia. IV-okcipitálny lalok (17, 18, 19): + pocit svetla; -(-vnímanie farieb; -(-vnímanie pohybu; + vnem lineárnych obrazov. Vizuálne asociatívne polia (18, 19): -(-kontralaterálne pohyby očí, ako je plynulé prenasledovanie; + vergenčné pohyby; -Antonov syndróm
Primárna motorická oblasť(motorická oblasť; 4. pole podľa Brodmana).
motorovej oblasti b je pred centrálnym sulcusom, ktorý oddeľuje predný lalok od parietálneho laloku. Skladá sa z veľkého počtu Betzových obrovských pyramídových buniek.
Predstavuje pohyby, nie jednotlivé svaly. Bunky, ktoré riadia pohyby, sú umiestnené tak, že pohyby dolnej polovice tela sú prezentované v hornej časti gyrusu a hornej polovice v dolnej. Neurónové axóny zostupujú cez vnútornú kapsulu a stopky mozgu vo forme kortikobulbárnych a kortikospinálnych dráh.
Odstránenie motorickej kôry vedie k paralýze.
Premotorická oblasť(pole b) leží priamo pred primárom motorická kôra, ktorý zaberá zadnú časť troch horizontálne orientovaných čelných gyri.
Tvorí vlákna frontopontocerebelárnych ciest a tiež dáva početné vlákna do bazálnych ganglií.
Navrhuje sa, aby sa premotorická kôra podieľala na plánovaní motorických akcií. Svedčí o tom skutočnosť, že iba pri premýšľaní o zamýšľanej akcii sú neuróny šiesteho poľa vzrušené. Stimulácia týchto neurónov vedie k zdvihnutiu rúk opačná strana, a hlava a oči sa otáčajú v smere zdvihnutej končatiny. Zreničky sa rozširujú a reč je narušená (pozri obr. 4.1.12, 4.1.13).
Deštrukcia tejto oblasti je sprevádzaná pseudocerebelárna ataxia opačnej strany. Zhoršená je aj schopnosť kopírovať naučené pohyby (apraxia). V tomto prípade sa jednotlivé pohyby nepoškodia, ale pacient ich nedokáže produkovať v požadovanom poradí.
Predná motorická vizuálna oblasť(pole 8 podľa Brodmana).
Táto oblasť (pole 8 a čiastočne 6 a 9) zaberá zadnú časť stredného frontálneho gyru (gyrus frontalis medius).
Predná motorická vizuálna oblasť dôležité pri dobrovoľných pohyboch očí. Niektoré oblasti definujú konvergenciu, divergenciu a akomodáciu.
Je potrebné poznamenať, že pohyb očí, zrenice a očných viečok možno zistiť počas elektrickej stimulácie takmer všetkých oblastí mozgových hemisfér. Elektrická stimulácia prednej motorickej oblasti oka vedie k sakadickým pohybom, na rozdiel od reakcie na stimuláciu polí 18, 19 a 22 (hladké sledovanie a vergenčné pohyby). Stimulácia 8. poľa u opíc je sprevádzaná súčasnou kontralaterálnou deviáciou očí. V tomto prípade sa zrenica rozširuje a hlava a trup sa otáčajú opačným smerom. Poškodenie predného laloku spôsobuje reverzibilnú stratu schopnosti kontralaterálnych sakadických pohybov.
Táto oblasť kôry sa vyznačuje mnohými spojeniami.
Aferentné vlákna vstupujú do zrakovej frontálnej kôry zo zrakového tuberkula, ako aj z hornej temporálnej, peristriatálnej a parietálnej (oblasť 7) kôry. Našli sa tu neuróny, ktoré riadia pohyby očí.
Efferents premietané do bazálnych ganglií, talamu, pretektálnej oblasti (vrátane jadra zrakového traktu), colliculi superior kvadrigeminy a časti retikulárna formácia Most.
Na druhej strane je potrebné poznamenať, že čelné vizuálne oblasti sa premietajú na kontralaterálne čelné oblasti a na kortikálne oblasti z rovnakej strany. Poskytujú zrakové vnímanie (na 7. poli parietálnej kôry a peristriatálny kortex).
Pri premietaní na stredný mozog vlákna prefrontálneho kortexu pretínajú prednú časť vnútorného puzdra a delia sa na dorzálne a ventrálne dráhy v rostrálnom diencefalóne.
Dorzálna transtalamická dráha prechádza dorzomediálnym a intralamylárnym jadrom thalamus opticus a strednou časťou vankúša, pričom zároveň netvorí veľké množstvo synaptické spojenia. Synapsie sa nachádzajú aj v pretektálnych jadrách a v horných tuberkulách kvadrigeminy.
Ventrálna pedunculo-tegmentálna dráha prechádza v najventrálnejšej časti mozgového kmeňa, dodáva vlákna do jadier hypotalamu a hlbšie - do horných tuberkulov kvadrigeminy. Súčasne majú čelné zrakové oblasti topografickú projekciu na horné tuberkuly kvadrigeminy. Cesta končí v retikulárnej formácii mosta.
Tretí prefrontálny väz sa vyskytuje v oblasti diencefalicko-mezencefalického spojenia nachádzajúceho sa v blízkosti červeného jadra. Táto dráha sa premieta na jadro mediálneho pozdĺžneho fascikula a intermediárne jadro Cajala na tej istej strane, ako aj na jadro Darkshevicha a rastrálnu časť okulomotorického jadra na oboch stranách.
Rôzne oblasti prednej vizuálnej kôry kontrolujú sakadické pohyby očí. rozdielna amplitúda, ako aj smer sakád. Poskytujú takzvanú "vizuálnu pozornosť", nasmerujú oči k "cieľu".
Momentálne sa to vyšetruje úloha predného laloku vo fungovaní očných viečok. Dobrovoľné zatvorenie očných viečok si vyžaduje uvoľnenie viečka zdvíhača a kontrakciu m. orbicularis oculi. Existuje kontroverzia týkajúca sa lokalizácie oblastí mozgu, ktoré riadia vôľové pohyby očného viečka. Tieto miesta môžu byť lokalizované v motorických premotorických alebo predných motorických vizuálnych oblastiach. Stimulácia tejto oblasti vedie k uzavretiu kontralaterálneho očného viečka. Jednostranné poškodenie nedominantných predných lalokov môže byť sprevádzané bilaterálnym zlyhaním zatvárania očných viečok.
Brocova oblasť motorickej reči(polia 44 a 45 podľa Brodmana).
Táto oblasť sa nachádza v zadnej-spodnej časti predného laloku. Prispieva k iniciácii reči. Poškodenie oblasti Broca vedie k afázii, ktorá spočíva v ťažkostiach spájania v určitom slede slov na poskytnutie súvisiacej reči (expresívna afázia). Pacienti rozumejú písanému aj hovorenému jazyku, ale zvyčajne nevedia nič vysloviť. Zhoršená je aj schopnosť písať, hoci ruku možno použiť aj na iné činnosti.
frontálna asociačná oblasť. Táto dôležitá oblasť v prednej polovici predného laloku je zodpovedná za spustenie všetkých typov behaviorálnych-intelektuálnych, viscerálnych a emocionálnych funkcií. Zmeny v pamäti, inteligencii a osobnosti nastávajú pri podráždení alebo masívnom zničení čelných lalokov. V týchto prípadoch je to obzvlášť výrazné. keď je ovplyvnená dominantná hemisféra.
Čuchový trakt sa nachádza pod orbitálnym povrchom predného laloka a je jedinou senzorickou dráhou, ktorá sa dostáva do kôry bez tvorby synapsií v talame. Jednostranný nádor orbitálneho povrchu predného laloka (meningióm čuchového sulcus alebo meningióm menšieho krídla) sfenoidálna kosť) môže viesť k stlačeniu čuchového traktu a optický nerv. Výsledkom je anosmia (nedostatok čuchu), ipsilaterálna atrofia zrakového nervu a opuch terča zrakového nervu na opačnej strane (Kennedy Foster syndróm).
Poškodenie štruktúr limbického systému vo frontálnom laloku, najmä gyrus cingulate, môže spôsobiť poruchy v emocionálnej stránke života, ako aj viscerálne znaky, vrátane bezpríčinného slzenia a zmeny v reakcii zreníc na svetelné podnety.
parietálny lalok(pozri obr. 4.1.12, 4.1.13). Vo všeobecnosti je funkcia parietálnych lalokov spojená s citlivosťou, pamäťou súvisiacou s rečou a učením, ako aj s orientáciou tela v priestore prostredníctvom spracovania vizuálnych informácií.
V prednom parietálnom laloku je gyrus umiestnený rovnobežne s precentrálnym gyrusom alebo motorickou kôrou. Ide o postcentrálny gyrus alebo primárny somatosenzorický kortex (polia 3, 1 a 2) (pozri obr. 4.1.13). Zvyšok parietálneho laloku možno rozdeliť na dva laloky - horný a dolný. Polia 5 a 7, umiestnené na povrchu horného parietálneho laloku, sú spojené s koreláciou somatosenzorických informácií, čo umožňuje vedome posúdiť hmotnosť, povrchové vlastnosti, veľkosť a tvar objektu.
Dolný parietálny lalok(supramarginálny a uhlový gyrus) úzko súvisí s rečovými mechanizmami, pretože deštrukcia tejto oblasti kôry v ľavej hemisfére vedie k narušeniu vnímania reči.
Úloha parietálneho laloku v priestorovej orientácii ešte nebola študovaná. U pacientov s jeho porážkou je však zaznamenané porušenie tejto funkcie.
Aferentné vlákna prechádzajú cez parietálny lalok, spájajú oblasti 18 a 19 s čelnými lalokmi a mozgovým kmeňom. Tieto dráhy zahŕňajú horné a dolné pozdĺžne zväzky.
Predpokladá sa, že parietálny kortex (pole 7) poskytuje funkciu "pozornosti" na vizuálne ciele.
Siedme pole prijíma aferentáciu z cingulate gyrus(g. cinguli), z pretektálnej oblasti a horných tuberkul quadrigeminy cez jadrá talamu. Nájdené sú aj aferenty smerujúce z prednej vizuálnej oblasti a prestriátovej kôry do kôry parietálneho laloku.
Eferenty sa premietajú do pretektálnej oblasti, horných tuberkulov štvorkolky, šedej hmoty umiestnenej okolo sylviánskeho akvaduktu a do parietálnych lalokov dvoch hemisfér mozgu.
Keď sa oči pohybujú, aktivujú sa neuróny dolného parietálneho laloku, čo naznačuje ich úlohu pri kontrole dobrovoľných pohybov očí. Aktivita pokračuje počas trvania pozorovania cieľa.
Bunky 7. poľa neurčujú sakadické pohyby, pretože ich aktivita sa počas sakád nezvyšuje.
Stimulácia uhlového gyrusu(g. angularis) vedie u pokusných zvierat k priateľskej deviácii očí. Ide o polia 38 a 39, prechádzajúce okolo zadného okraja stredného temporálneho gyru (g. temporalis medius). Operatívne odstránenie táto oblasť kôry nevedie k zhoršeniu pohybu očí. Obojstranné poškodenie parietálnej kôry môže viesť k rozvoju motorickej apraxie oka.
Porážka parietálneho laloku je tiež sprevádzaná porušením vizuálnych motorických mechanizmov, ktoré poskytujú optokinetický nystagmus, počas ktorého oči produkujú sakády na vyhľadávanie a fixáciu cieľa.
Medzi ďalšie motorické príznaky izolovaných lézií parietálneho laloka patria priateľská odchýlka očí v opačnom smere od miesta poranenia s núteným zatváraním viečok, nízkou pohyblivosťou pacienta a zlým ovládaním (nepochopením) končatín opačnej strany. Ťažkosti s pochopením podstaty činnosti končatín vedú ku konštitučnej a okulomotorickej apraxii. Pacienti s poškodením parietálneho laloku môžu mať ťažkosti s pohybom pohľadu z jedného vizuálneho objektu na druhý v kontralaterálnej polovici zorného poľa.
Ďalším znakom dysfunkcie parietálneho laloku je neschopnosť opraviť vizuálny objekt.
Okrem zmyslových funkcií, ktoré sú lokalizované v zadnej pere Rolandovho centrálneho sulku, je parietálny lalok miestom zložitého procesu rozpoznávania a chápania. Difúzne poškodenie parietálnych lalokov končí agnóziou, zrakovou nepozornosťou, metamorfopsiou, fotopsiou, halucináciami, ilúziami a alexiou (pozri obr. 4.1.12, 4.1.13).
Viac podrobnosti o dôsledkoch dysfunkcie parietálneho laloka sú uvedené v neurologickej literatúre. Tu uvádzame len niektoré zo syndrómov.
vizuálna agnózia, ktorý sa často vyvíja v rozpore s krvným obehom v centrálnom nervovom systéme, sa vyznačuje porušením procesu rozpoznávania. Agnózia môže byť úplná alebo čiastočná. Vizuálna agnózia je diagnostikovaná v prípadoch, keď pacient dokáže identifikovať objekt iba hmatom. Často je narušené rozpoznávanie a chápanie vizuálnych obrazov a farebného významu obrazu.
Obojstranné poškodenie parietálneho laloka (častejšie pri senilnej demencii, mäknutie temporookcipitálnych a parietálnych oblastí kôry, poranenia zadných úsekov oboch hemisfér, encefalitída) sprevádza zraková agnózia a zraková ataxia (Balintov syndróm).
Vizuálna priestorová agnózia charakterizované ťažkosťami pri posudzovaní veľkosti objektu a vzdialenosti k nemu. Zaznamenáva sa aj vznik nesprávneho riešenia „syntetických“ priestorových problémov, ako je čítanie mapy. Simultánna agnózia sa scvrkáva na skutočnosť, že pacienti si zachovávajú schopnosť rozpoznať jednotlivé objekty, ale chýba im schopnosť vnímať skupinu objektov (obrázkov) ako celok.
Asterognostia vyznačujúci sa tým, že pacient dokáže analyzovať predmety podľa ich vzhľadu, ale nie je schopný ich hmatovo charakterizovať. To sa vyvíja v dôsledku narušenia komplexnej interakcie medzi hmatovými a vizuálnymi informáciami v parietálnych lalokoch kôry.
Difúzne lézie parietálnych lalokov môžu viesť k ilúziám a metamorfopsiám. Tieto skreslenia zahŕňajú mikropsiu, makropsiu. Pacienti zaznamenávajú pohyb stacionárnych cieľov, nesprávne odhadujú vzdialenosť k objektom. TO špeciálne typy ilúzie zahŕňajú papinopsiu (porušenie zrakového vnímania vo forme zachovania alebo opätovného objavenia sa vizuálneho obrazu po zmiznutí objektu zo zorného poľa), halucinačnej polyopie atď.
Ilúzie sa vyvíjajú v dôsledku difúznych porúch kôry a halucinácie a fotopsie sa vyvíjajú s rozvojom nádoru alebo toxického poškodenia kôry.
Charakteristiky patológie parietálneho laloku môžu závisieť aj od toho, či je patologické zameranie lokalizované v dominantných (zvyčajne ľavých) alebo nedominantných hemisférach. Dominantný parietálny lalok je zodpovedný za rozvoj porúch reči, ako je sluchová senzorická afázia, vizuálna agnózia a agrafiách.
Poškodenie dominantného uhlového gyrusu(častejšie v dôsledku porúch prekrvenia v povodí uhlovej tepny) vzniká nystagmus, zrakovo-agnostické poruchy, agrafia, akalkulia (neschopnosť sčítať alebo odčítať čísla), digitálna agnózia, neschopnosť rozlišovať medzi pravou a ľavou stranou ( Gerstmannov syndróm; syndróm uhlového gyrusu). Možno vývoj homonymnej hemianopsie.
Ak je poškodený nedominantný uhlový gyrus, je narušená interakcia pacienta s okolím. V dôsledku toho sa vyvíja zraková dezorientácia a topografická agnózia.
Vpredu k gyrusu uhlovému a cez sulcus supratemporalis je supramarginálny gyrus. V dominantnej hemisfére je táto oblasť miestom stretnutia zrakovej a sluchovej zložky jazyka.
Poškodenie zadných parietálnych asociačných oblastí často končí rozvoj apraxie(ťažkosti pri vykonávaní „jemných“ špecializovaných pohybov). Zjavne je to spôsobené nepochopením potrebnej postupnosti pohybov na vykonávanie určitej funkcie (t. j. pacientovi nie je jasné, čo a v akom poradí by sa malo vykonávať). Pacientovi chýba schopnosť nakresliť jednoduchý diagram (konštruktívna apraxia). Menej závažnou formou tejto patológie je hmatová agnózia (neschopnosť rozpoznať objekt pri palpácii). Niekedy sa táto choroba nazýva asterognostia. Zároveň nie sú žiadne známky straty taktilnej alebo proprioceptívnej citlivosti a je narušená integrácia vizuálnych a somatosenzorických informácií. Apraxia a asterognostia sa často vyvinú po poškodení ľavej hemisféry mozgu. Asterognostia je zvyčajne obmedzená na opačnú stranu tela, zatiaľ čo apraxia je zvyčajne obojstranná.
Vizuálne a sluchové informácie sú úzko spojené s funkciou jazyka. Z tohto dôvodu poškodenie oblastí 39 a 40, ako aj častí oblasti sluchovej asociácie, zvyčajne vyústi do afázie, tzv. afázia Wernicke(Wernicke). Ak je poškodenie obmedzené na uhlový gyrus, stráca sa schopnosť porozumieť písanému jazyku (alexia) a schopnosť písať (agraphia). Pacienti zároveň rozumejú reči. Tento stav sa zvyčajne vyvíja po zranení.
Integrácia vizuálnych a somatosenzorických informácií je dôležitá pre vytvorenie "body image", teda pochopenie znakov polohy tela v priestore. Rozsiahle poškodenie polí 7, 39 a 40 jednej hemisféry môže viesť k rozvoju „nepochopenia“ alebo „zanedbávanie“ opačnej polovice tela. Pacienti nie sú schopní obviazať alebo rozviazať svoje vlastné končatiny. Nepoznajú ani vlastné končatiny.
Okcipitálny lalok(polia 17, 18 a 19). Pozdĺž ostrohy (fissura calcarina) je primárna zraková oblasť (pole 17). S ním susedí „hodnotiaca“ vizuálna oblasť (pole 18), o ktorej sa predpokladá, že prevádza signály prijaté poľom 17 na vizuálne obrazy.
Oblasť 17 okcipitálneho kortexu je zóna príjmu a spracovania vizuálnych informácií a má dôležité súvislosti s motorickými vizuálnymi oblasťami kôry. Práve na tejto úrovni zrakovej kôry sa hodnotí význam videného. Podráždenie tejto oblasti spôsobuje halucinácie a snové obrazy.
Pozornosť si zasluhujú aj motorické spojenia okcipitálneho laloku, pretože sa podieľajú na optokinetickom nystagme, akomodačnom reflexe a regulácii binokulárneho videnia.
Polia 18 a 19 sú vzájomne závislé a majú viac spojení so zvyškom mozgu. Predpokladá sa, že oblasť 18 sa viac zaujíma o integráciu vizuálnych informácií. prijímané v 17. poli, pričom pole 19 prevádza tieto informácie do zložitejšej mentálnej a motorickej činnosti.
Nedávne anatomické a fyziologický výskum odhalili, že existujú najmenej, 10 podoblastí polí 18 a 19, ktorých neuróny majú schopnosť reagovať na vizuálne podnety. Každá z týchto podoblastí je spojená so 17. poľom. Navzájom však nesúvisia. Poškodenie týchto podoblastí nevedie k slepote. Niektoré sú porušené zrakové funkcie, ako je analýza farieb, vnímanie integrity objektu atď.
zraková asociačná kôra, ktorý sa nachádza v niektorých oblastiach spánkového laloku, sa tiež podieľa na spracovaní vizuálnych informácií. To platí v najväčšej miere pre pole 21 (v strednej temporálnej kôre). Práve toto pole poskytuje rozpoznanie obrazu a jeho poškodenie končí agnóziou.
Vyššie uvedené vlastnosti zrakovej kôry poskytujú aj početné horizontálne a vertikálne spojenia medzi neurónmi v rôznych vrstvách zrakovej kôry. Je to kvôli ich prítomnosti, že výnimočná plasticita zrakovej kôry je zaznamenaná pri spracovaní vizuálnych informácií rôzneho typu.
Štrukturálna organizácia zrakovej kôry a jej funkcie budú podrobnejšie uvedené v ďalšej časti.
temporálny lalok(polia 41, 22).
Táto časť mozgu spojený s vnímaním sluchových vnemov, podieľa sa na funkcii reči prostredníctvom sluchovej kontroly reči a zohráva úlohu aj pri hodnotení priestorovej a pamäťovej funkcie. Už dlho sa zistilo, že transverzálny temporálny gyrus (pole 41) je primárnym sluchovým centrom. Malé pole (22). okolo tohto gyrusu sa považuje za „mentálny“ alebo „hodnotiaci“, sluchové centrum. Podráždenie väčšiny temporálneho laloku, najmä pozdĺž stredného temporálneho gyrusu, spôsobuje pocity, ktoré pacient popisuje ako sluchové. Pacienti s poškodením „mentálneho poľa“ spánkového laloku niekedy strácajú schopnosť posúdiť tóny zvuku. Pacient zároveň nepozná motívy a hudobné zvuky vníma ako náhodný hluk.
Pole 22 ľavej hemisféry je tiež spojené s funkciou reči, pretože poškodenie tejto oblasti vedie k strate schopnosti porozumieť významu slov. Existujú tiež dôkazy, že temporálny lalok je spojený s „vestibulárnou“ citlivosťou (zmysel pre rovnováhu), pretože stimulácia zadného horného temporálneho gyru u pacienta pri vedomí spôsobuje závraty a pocit rotácie.
Hlboké lézie v temporálnom laloku často zahŕňajú najnižšie vlákna optického žiarenia z laterálneho genikulárneho tela. V čom zrakové postihnutie. Najbežnejšia je hemianopsia horného kvadrantu.
S porážkou temporálneho alebo psychomotorického laloku sa vyvíja epilepsia sprevádzaná zrakovými halucináciami, pocit deja vu a deja pense („už videné“ - fr.), v ktorom majú vizuálny zážitok a myšlienky zvláštny priateľský vzťah, objavujú sa automatizmy, ktoré sprevádzajú amnéziu, nutkavé správanie, hnev a úzkosť.
U pacientov s jednostranným poškodením primár sluchová kôra mierna strata sluchu. Pri porušení sluchového rozpoznávania (sluchová agnózia) sú ovplyvnené asociatívne sluchové polia. Pacienti zvyčajne uvádzajú, že počujú zvuky, ale nedokážu ich identifikovať.
Oblasť zapojená do rozpoznávania tváre sa nachádza na ventrálnom povrchu spánkového laloku. Poškodenie tejto oblasti (častejšie v rozpore s krvným obehom v oblasti bazilárnej tepny mozgu), ktorá sa spravidla rozprestiera na priľahlý okcipitálny lalok (polia 20, 21), obe hemisféry, sprevádzaný rozvojom prosopagnózie(neschopnosť rozpoznať tváre). Vo všeobecnosti tento stav nie je sprevádzaný inými neurologickými príznakmi. Pacient vie čítať a pomenovať predmety. V prípadoch, keď pacient počuje hlas známeho človeka, vie ho okamžite identifikovať.
Zostávajúca časť horného temporálneho gyru je obsadená oblasťou 22 (auditívna asociačná kôra), ktorá prijíma veľké množstvo aferentov z polí 41 a 42, aby poskytla vlákna do parietálnych a okcipitálnych lalokov. Oblasť 22 sa podieľa aj na jazykových funkciách. Poškodenie hornej časti 22. poľa vedie k ťažkostiam s porozumením jazyka.
Spánkové laloky sú obzvlášť dôležité pre pamäťovú funkciu.. Napríklad stimulácia sluchovej asociačnej kôry u pacientov počas neurochirurgických operácií vyvoláva zložité spomienky, sluchové aj zrakové. Pri rozsiahlom poškodení spánkového laloku dochádza k zhoršeniu pamäti.
limbický lalok(obr. 4.1.14).
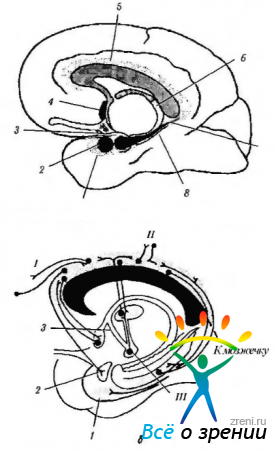
Ryža. 4.1.14. Limbický systém (podľa Brodsla, 1992): a - umiestnenie štruktúr limbického systému (1 - entorálne pole; 2 - amygdala; 3 - hák; 4 - septálne jadro; 5 - gyrus cingulate; b - fornix; 7 - hippocampus; 8 - parahippokampálny gyrus); b - spojenia gyrus cingulate (I - frontálna asociatívna kôra; II - parietálna a temporálna asociatívna kôra; III - mastoidné telo). Cingulate gyrus je spojený s asociatívnymi poľami mozgovej kôry a limbického systému a poskytuje interakciu medzi nimi.
Limbický systém je komplex formácií terminálu, diencefala a stredného mozgu. Vlastne " limbický systém“ sa zhoduje s predtým používaným pojmom – „čuchový mozog“.
Ak nová kôra (neokortex) riadi časopriestorové vzťahy organizmu s prostredím a zodpovedá aj za formálno-logické myslenie a stereognostické schopnosti, potom limbický systém určuje emocionálne funkcie a motiváciu konania ako aj procesy učenia a pamäte. Tiež riadi homeostázu. Jeho dôležitou funkciou je organizácia behaviorálnych reakcií jednotlivca v reakcii na vplyv vonkajšieho prostredia a zmeny vnútorného prostredia organizmu. Tieto reakcie sú zamerané na zachovanie jedinca a sú sprostredkované cez nižšie centrá diencefala.
Limbický lalok má tiež niečo spoločné s funkciou čuchu, pretože podráždenie alebo poškodenie hipokampálnej oblasti je sprevádzané čuchom.
Vytvára sa limbický lalok:
- subkauzálna oblasť (area subcallosa),
- cingulate gyrus (gyrus cinguli),
- isthmus cingulate gyrus
- parahippokampálny gyrus (gyrus parahippocampalis),
- hippocampus (amónový roh, gyrus dentatus a spodok hipokampu alebo subiculum)
- a Brocov diagonálny väz.
Medzi subkortikálne štruktúry limbického systému patria mandle (corpus amygdaloideum), septálne jadrá a predné talamické jadro.
Aferentné a eferentné spojenia štruktúr limbického systému, ako medzi sebou, tak aj s inými časťami mozgu, sú mimoriadne rozmanité. Najvýraznejšie sú jeho vzájomné spojenia s hypotalamom.
Hypotalamus a prsné telieska spojený s hipokampom a septálnou oblasťou cez fornix, s amygdalou cez terminálnu striu a amygdalofugálny zväzok a s čuchovým mozgom cez mediálny zväzok predný mozog. Limbický systém je spojený so stredným mozgom cez hypotalamus a prsné telieska.
Limbický systém komunikuje s neokortexom vo frontálnych a temporálny lalok. Časové regióny sú zodpovedné najmä za prenos informácií zo zrakovej, sluchovej a somatickej kôry do amygdaly a hipokampu. Predné oblasti slúžia ako hlavné oddelenie neokortexu, regulujúc limbické zóny.
Bazálna uzlina predstavujú subkortikálnu akumuláciu neurónov vo forme skupiny jadier (obr. 4.1.15; 4.1.16).
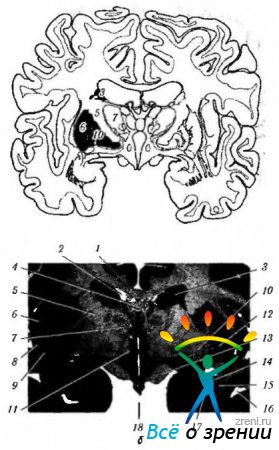
Ryža. 4.1.15. Bazálna uzlina: a - schematické znázornenie umiestnenia štruktúr súvisiacich s bazálnymi gangliami; b - frontálny úsek mozgu prechádzajúci štruktúrami súvisiacimi s bazálnymi gangliami (4 - corpus callosum; 2 - roh bočná komora; 3- telo nucleus caudatus; 4-klenba; 5-vnútorná kapsula; 6 - ostrovček; 7 - vizuálny tuberkul; 8- škrupina; 9 - plot; 10 - bledá guľa; 11 - hypotuberózna oblasť; 12 - bezmenná látka: 13 - zrakový trakt; 14 - zadná spodná časť nucleus caudate; 15 - mandľa: 16 - temporálny roh laterálnej komory; 17 - čelný roh laterálnej komory; 18 - tretia komora)
Ryža. 4.1.16. Najdôležitejšie aferentné, eferentné a vnútorné spojenia bazálnych ganglií a ich spojenie s mozočkom (vysvetlenie v texte): a - (1-motorická kôra; 2 - eferentné vlákna z kôry do striata; 3 - striatum; 4 - bledá guľa; 5 - aferentné vlákna; 6 - zrakový tuberkulum; 7 - subtalamické jadro; 8 - substantia nigra); b - (1 - mozgová kôra (motorická); 2 - bazálne gangliá; 3 - cerebellum; 4 - motorické a interkalárne neuróny)
Ovládajú pohyb tela. Táto funkcia bazálnych ganglií bola stanovená na základe klinických pozorovaní. Keď sú poškodené, dochádza k porušeniu motorická aktivita ako vo forme neschopnosti iniciovať potrebné pohyby, tak aj neschopnosti pohyby potlačiť.
Pri Parkinsonovej chorobe, ku ktorej dochádza pri poškodení bazálnych ganglií, majú pacienti „nevýraznú“ tvár. To je spojené s poruchou motorickej aktivity tvárových svalov a kontroly pohybov očí, najmä poruchy sakadických pohybov. Práve z tohto dôvodu sa bližšie pozrieme na neuroanatómiu tejto oblasti.
Existujú tri hlavné akumulácie subkortikálnych jadier ktoré sa nazývajú striatum ( corpus striatum), plot (claustrum) a amygdala (corpus amigdaloideum).
- Corpus striatum sa skladá z dvoch častí – nucleus caudatus (nucleus caudatus) a lentiformného jadra (nucleus lentiformis).
Caudate jadro leží nad a mediálne od lentikulárneho jadra, oddelené od neho vrstvou bielej hmoty nazývanej vnútorná kapsula (capsula interna). Zhrubnutá predná časť jadra caudate, jeho hlava (caput nuclei caudati) tvorí bočnú stenu predný roh laterálna komora, zatiaľ čo zadná tenká časť nucleus caudatus (corpus et cauda nuclei caudati) sa tiahne späť pozdĺž spodnej časti centrálnej časti laterálnej komory. Na mediálnej strane prilieha nucleus caudatus k zrakovému pahorku, ktorý je od neho oddelený pásikom bielej hmoty (stria terminalis). Prednou a inferiornou hlavicou nucleus caudate dosahuje predná perforovaná substancia (substantia perforata anterior). V tomto bode je hlava spojená s lentikulárnym jadrom (s časťou nazývanou putamen). Okrem tohto širokého spojenia oboch jadier na ventrálnej strane sú tu aj tenké pásiky šedej hmoty popretkávané bielymi chumáčmi vnútorného puzdra. Z nich vznikol názov „prúžkované teleso“ (corpus striatum).
Lentikulárne jadro(nucleus lentiformis) leží laterálne od nucleus caudatus a zrakového pahorku, oddelený od nich vnútorným puzdrom. Na vodorovnom reze hemisféry má stredný povrch lentiformného jadra, smerujúci k vnútornej kapsule, tvar krehkosti s vrcholom smerujúcim do stredu. Predná strana uhla je rovnobežná s nucleus caudate a zadná strana je rovnobežná s talamom. Bočný povrch je mierne konvexný a smeruje k laterálnej strane hemisféry v oblasti ostrovčeka. Predné a ventrálne, ako už bolo naznačené, lentiformné jadro splýva s hlavou nucleus caudatus.
Lentiformné jadro má na frontálnom úseku klinovitý tvar, ktorého vrchol je otočený na mediálnu stranu a báza je laterálne. Lentikulárne jadro je rozdelené na tri segmenty dvoma rovnobežnými bielymi vrstvami (laminae medullares). Bočne umiestnená tmavosivá oblasť sa nazýva škrupina (putamen) a dve stredné, svetlejšie, sa spolu nazývajú bledá guľa (globus pallidus).
Líši sa už svojím makroskopickým vzhľadom, bledá guľa(globus pallidus) má aj histologickú štruktúru, ktorá sa výrazne líši od štruktúry ostatných častí striata.
Vzhľadom na všetky tieto znaky sa bledá guľa rozlišuje na osobitnú morfologickú jednotku nazývanú pallidum, pričom označenie striatum je ponechané len pre putamen a nucleus caudatus. V dôsledku toho výraz „lentikulárne jadro“ stráca svoj pôvodný význam a môže sa používať iba v čisto topografickom zmysle. Súčasne namiesto doterajšieho názvu corpus striatum sa kaudátne a lentikulárne jadrá nazývajú striopallidarový systém.
Striopallidárny systém je hlavnou súčasťou extrapyramídového systému a okrem toho je najvyšším regulačným centrom autonómne funkcie vo vzťahu k termoregulácii a metabolizmu sacharidov, dominujúce nad podobnými vegetatívnymi centrami v hypotalame.
- Plot(claustrum) je tenká doska šedej hmoty, položená v oblasti ostrova, medzi ním a putamenom. Od druhej je oddelená vrstvou bielej hmoty nazývanej vonkajšia kapsula (capsula externa).
- amygdala(corpus amygdaloideum) sa nachádza pod putamenom na prednom konci spánkového laloku. Zdá sa, že amygdala patrí k subkortikálnym čuchovým centrám a limbickému systému. Končí sa vo zväzku vlákien vychádzajúcich z čuchového laloku a končí predná perforovaná substancia (substantia perforata anterior), uvedená v opise zrakového kopca nazývaného stria.
Úloha amygdaly u ľudí nie je dobre pochopená. V podstate bola úloha amygdaly odhalená na základe štúdie neurologických pacientov. Vďaka týmto štúdiám sa zistilo, že mechanická alebo elektrická stimulácia amygdaly u ľudí často vedie k strachu alebo iným emocionálnym reakciám. Teda ľudská amygdala podieľa sa na procesoch emocionálneho vnímania. Ukazuje sa tiež, že pri obojstrannom poškodení mandlí dochádza k strate "emocionálnej" pamäte, vzhľadu "vystrašeného" výrazu tváre a k porušeniu integrácie čuchových a vizuálnych informácií.
Je dobre známe, že človek má schopnosť určiť náladu a emocionálny stav iného jedinca. IN V poslednej dobe zapojenie amygdaly do tohto komplexu fyziologický proces. Pri určovaní emocionálneho stavu človeka náš mozog zachytáva a analyzuje jemné zmeny výrazu tváre a predovšetkým smeru pohľadu. Nervové mechanizmy, ktoré sú základom týchto procesov, nie sú presne jasné. Napriek tomu sa zistilo, že „presné“ nastavenie pohľadu, psychofyziologicky hodnotené ako prítomnosť „pozornosti“, určený top temporálna kôra a amygdala. V štúdii pacientov s autizmom sa ukázalo, že často majú anomáliu amygdaly alebo jej poškodenie. Bezprostredné mechanizmy, ktoré sú základom sociálneho úsudku iných osôb na základe orientácie pohľadu, zostávajú do značnej miery neznáme.
V poslednom čase sa za bazálne gangliá označujú aj substantia nigra a subtalamické jadrá. Okrem toho sa v substantia nigra rozlišujú 2 časti - retikulárna (pars reticulata) a kompaktná (pars compacta).
Morfologické a elektrofyziologické metódy ukázali, že väčšina z nich aferentné signály prichádzajúce do bazálnych ganglií vstupujú do striata (pozri obr. 4.1.16). Tieto signály pochádzajú z mnohých zdrojov, z ktorých hlavné sú:
- všetky oblasti kôry hemisféry;
- intralamelárne jadrá talamu
- substantia nigra (pozdĺž dopaminergnej dráhy).
Z vnútra bledej zemegule pochádza to najdôležitejšie zo všetkých eferentné cesty bazálna uzlina. Táto cesta končí hlavne vo zrakovom tuberkule a streche stredného mozgu.
Bazálne gangliá teda zohrávajú úlohu medzičlánku v reťazci spájajúcej motorické oblasti kôry so všetkými jej ostatnými oblasťami. Ich hlavnou funkciou je „plánovanie“ fyzickej aktivity.
IN posledné roky zistili, že bazálne gangliá spolu s niektorými kortikálnymi oblasťami určujú aj niekt kognitívne funkcie vrátane pozornosti, pamäti. Pojem „pozornosť“ v najširšom zmysle slova je „proces výberu“. Vzhľadom na to, že proces orientácie v priestore a výber cieľa záujmu, na ktorom sa podieľajú sakadické pohyby očí, je spojený s bazálnymi gangliami, je zrejmá významná úloha bazálneho komplexu v procesoch orientácie cez sakády. V tomto prípade má osobitné miesto pri kontrole sakád oka substantia nigra, najmä jej retikulárna časť. Anatomicky aj elektrofyziologicky bola dokázaná prítomnosť jeho spojenia s hornými tuberkulami kvadrigeminy. Hlavnou funkciou neurónov v substantia nigra je inhibovať aktivitu neurónov colliculus superior quadrigeminy, ktoré sa podieľajú na tvorbe sakád.
V motorickej aktivite očí zúčastňuje sa aj caudate nucleus, ktoré má spojenie so substantia nigra a vonkajšími tuberkulami quadrigeminy. Excitácia caudatus nucleus vedie k sakadickým pohybom oka v opačnom smere od excitovaného jadra.
Subtalamické jadro sa tiež podieľa na pohybe očí. Na toto jadro sa premieta bledá guľa, kôra prednej oblasti. Vlákna odchádzajú zo subtalamického jadra do substantia nigra, vonkajších a vnútorných segmentov globus pallidus.
Menej jasná je funkcia globus pallidus v okulomotorickej aktivite. Je to spôsobené tým, že globus pallidus má veľké množstvo spojení medzi rôznymi jadrami bazálnych ganglií a málo spojení s nervových centier nachádza sa mimo bazálnych ganglií. Okulomotorické neuróny boli nájdené iba v dorzálnej časti globus pallidus, t.j. v časti prijímajúcej vstupy z nucleus caudate.
Úloha bazálnych ganglií pri pohybe očí potvrdené mnohými klinickými pozorovaniami. Pri Parkinsonovej chorobe, sprevádzanej degeneráciou bazálnych ganglií a sivej hmoty stredného mozgu a prerušením frontomesencefalických dráh, vzniká progresívna supranukleárna obrna. U takýchto pacientov je rýchlosť sakadických pohybov očí znížená. Sakády sú zvyčajne hypometrické, čo je spojené s porušením funkcie "saccade memory". Očný pohyb sa často nedá dokončiť celý. Vertikálne pohyby očí sú ovplyvnené výraznejšie ako horizontálne. Pacienti tiež vykazujú axiálnu rigiditu a demenciu. Podobné zmeny boli zistené aj pri Huntingtonovej chorobe.
corpus callosum(corpus callosum). Corpus callosum je veľký zväzok vlákien (pozostáva z približne 250 miliónov vlákien), ktorý spája obe hemisféry (obr. 4.1.17).
Ryža. 4.1.17. Lokalizácia jadier talamu: a - jadrová paramagnetická rezonancia; b - predná časť mozgu (1 - telo corpus callosum; 2 - telo nucleus caudatus; 3 - centrálna časť laterálnej komory; 4 - priehľadná priehradka; 5 - choroidný plexus bočná komora; 6 - klenba; 7 - predná skupina jadier vizuálneho pahorku; 8- vonkajšia kapsula; 9 - laterálna skupina jadier talamu; 10- mediálne jadrá talamu: - schránka; 12- laterálna dreňová platnička; 13 - vnútorná kapsula; 14 - retikulárne jadro talamu; 15 - intertalamický hrot; 16 - bočná časť bledej gule; 17 - mediálna medulárna doska; 18 - mamilotalamický zväzok; 19 - HI Pstruhové pole; 20 - gopa incerta; 21 - H2 Pstruhové pole; 22 - plot; 23-mediálna časť bledej gule; 24 - tretia komora; 25 - jadrá hypotalamu; 26-zrakový trakt; 27 - amygdala; 28-vsuvkové telo; 29-základňa nôh mozgu)
Hlavnou funkciou tohto lúča je prenos informácií z jednej hemisféry do druhej.
Hlavné funkcie corpus callosum sú:
- Korelácia obrázkov na pravej a ľavej polovici zorného poľa.
- Integrácia vnemov pochádzajúcich zo spárovaných končatín, potrebná na učenie motorickej koordinácie.
- Integrácia pozornosti a aktivačných procesov v hemisférach.
U pacientov, u ktorých bolo corpus callosum prerezané na terapeutické účely, mozgová dysfunkcia v Každodenný život nebola dodržaná. Boli odhalené iba skrz špeciálne testy. U iných pacientov sa niekedy vyvinú dosť výrazné poruchy správania. Najčastejšie sa to prejavuje „slovnou slepotou“, „slovnou hluchotou“ a stratou koordinácie pohybov pravej a ľavej končatiny.
Pokračovanie v ďalšom článku: Anatómia mozgu? Časť 3
A to v očiach celého hlavného mesta
Kohútik trepotal z pletacej ihlice,
letel na voz
A posadil sa na korunu kráľa,
Preľaknutý, klovaný do koruny
A vzlietli... a zároveň
Dadon spadol z voza -
Raz zalapal po dychu a zomrel.
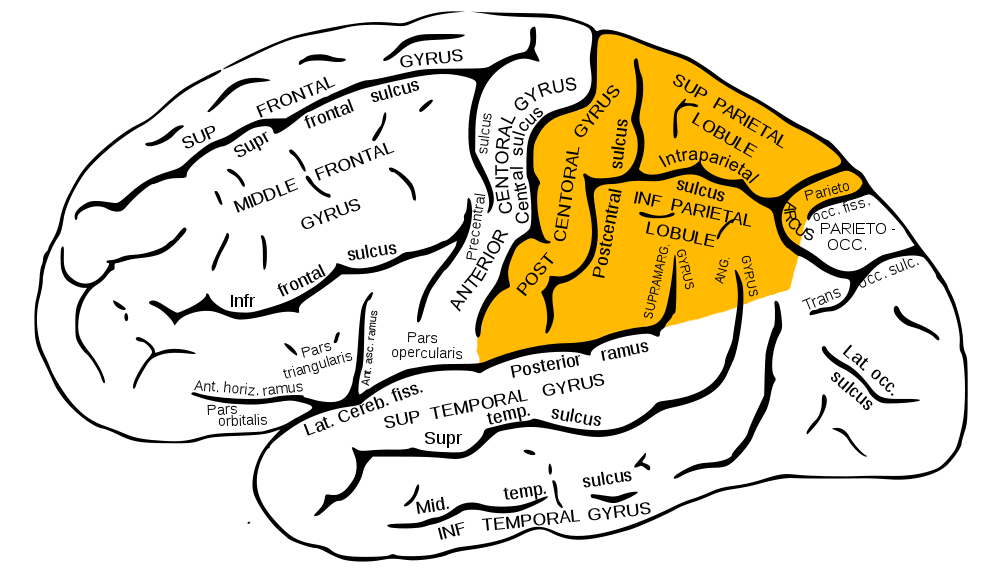
Zdá sa, že ste už pochopili, o čom sa bude diskutovať v ďalšom materiáli cyklu „Ako funguje mozog“. Už sme hovorili o čelných lalokoch, temporálnych a parietálnych, teraz sa obrátime na parietálne laloky. Sú to tiež Lobus parietalis v latinčine známe lekárom.
Parietálne laloky zobrazené žltou farbou
Parietálny lalok sa nachádza tesne nad týlnym hrbolčekom a „pozostáva“ z troch zvinutí: jedna vertikálna- zadná centrálna (najstaršia časť) a dve horizontálne - horná parietálna (nová) a dolná parietálna (novšia).
Rovnako ako štruktúra predných lalokov, časti ľudského tela sa „premietajú“ na centrálny predný gyrus parietálu: dolná tretina je tvár, stredná tretina je ruka a trup, horná je noha. Nezabúdajte, že podiel je „dvojnásobný“, teda jeho polovica je zodpovedná za jednu (opačnú) polovicu tela.
Štruktúra parietálnych lalokov
Okrem toho v hornom parietálnom gyruse sú centrá zodpovedné za komplexné typy hlboká citlivosť: svalovo-kĺbový, dvojrozmerný-priestorový zmysel (definícia čísel, písmen, číslic nakreslených ceruzkou alebo iným tupým predmetom na ľudskej koži), zmysel pre hmotnosť a objem pohybu, zmysel pre rozpoznávanie predmetov hmatom.
V dolnom parietálnom laloku sú centrá praxe, teda pohyby, ktoré sa stali „automatickými“ v procese opakovania a cvičenia, ktoré sa rozvíjajú v procese učenia a neustáleho precvičovania, napríklad chôdza, jedenie, obliekanie a tak ďalej.
Parietálny lalok sa podieľa na spracovaní a vedomom vnímaní somatosenzorických (z receptorov vo svaloch, koži, kĺboch a vnútorné orgány) informácie, ktoré ovplyvňujú dobrovoľné pohyby.
Lézie horného parietálneho laloku sú sprevádzané rozvojom narušenia schopnosti rozpoznávať predmety dotykom so zatvorenými očami. Pacienti opisujú jednotlivé vlastnosti objektu, ale nedokážu syntetizovať jeho obraz.
Pri porážke dolného parietálneho laloku je narušený pocit schémy tela. Človek si nie je schopný uvedomiť, kde je pravá a kde ľavá strana, nepozná vlastné prsty. Ďalším druhom poruchy je neuvedomenie si svojej chyby (pacient tvrdí, že hýbe ochrnutými končatinami). U týchto pacientov sa môže vyvinúť pseudopolymélia.— pocit extra končatiny alebo častí tela. Takíto pacienti môžu nezávisle odstrániť „zasahujúcu“ končatinu alebo prispieť k jej amputácii.
Pri poškodení kôry uhlového gyrusu pacient stráca zmysel pre priestorové vnímanie okolitého sveta, polohy vlastného tela a vzájomných prepojení jeho častí. To je sprevádzané rôznymi psychopatologickými príznakmi: depersonalizácia, derealizácia. Možno ich pozorovať pod podmienkou úplného zachovania vedomia a kritického myslenia.
S léziami parietálneho laloku dominantnej hemisféry sa u človeka vyvinie dyslexia - neschopnosť čítať, rozlišovať medzi pravou a ľavou stranou, ako aj dyskalkúlia - neschopnosť robiť aritmetiku. Je pozoruhodné, že dyskalkúlia je často nezávislou chorobou a nie dôsledkom neurologickej alebo psychické problémy. Okrem týchto problémov je možná aj apraxia - porušenie alebo neschopnosť vykonať nejakú účelnú činnosť (dobre, napríklad dať si pohár a vypiť) pri zachovaní základných zložiek akcie.
Anastasia Sheshuková
Okcipitálny lalok zaberá zadné časti hemisfér. Na konvexnom povrchu hemisféry nemá okcipitálny lalok ostré hranice, ktoré ho oddeľujú od parietálnych a temporálnych lalokov, s výnimkou hornej časti parietálno-okcipitálneho sulcus, ktorý sa nachádza na vnútornom povrchu hemisféry. oddeľuje temenný lalok od okcipitálneho laloku. Brázdy a zákruty horného bočného povrchu okcipitálneho laloku sú nestabilné a majú premenlivú štruktúru. Na vnútornom povrchu okcipitálneho laloku je ostruhová drážka, ktorá oddeľuje klin (trojuholníkový lalok okcipitálneho laloku) od lingválneho gyru a okcipitotemporálneho gyru.
Funkcia okcipitálneho laloku je spojená s vnímaním a spracovaním vizuálnych informácií, organizáciou zložitých procesov vizuálneho vnímania. V tomto prípade sa horná polovica sietnice premieta do oblasti klinu, ktorý vníma svetlo z dolných zorných polí; v oblasti lingulárneho gyru je spodná polovica sietnice, ktorá vníma svetlo z horných zorných polí.
ostrov
Ostrovček alebo takzvaný uzavretý lalok sa nachádza hlboko v bočnej drážke. Ostrovček je oddelený od susedných susedných častí kruhovou drážkou. Povrch ostrovčeka je svojou pozdĺžnou stredovou drážkou rozdelený na prednú a zadnú časť. V ostrovčeku je premietaný analyzátor chuti.
limbická kôra
Na vnútornom povrchu hemisfér nad corpus callosum je gyrus cingulate. Tento gyrus s isthmom za corpus callosum prechádza v blízkosti morského koníka do gyrusu - parahipocampal gyrus. Gyrus cingulate spolu s gyrus paragitpocampal tvorí klenutý gyrus.
Vnútorná a dolná plocha hemisfér sa spája do takzvanej limbickej (okrajovej) kôry, spolu s amygdalou zo skupiny subkortikálnych jadier, čuchovým traktom a bulbom, oblasťami čelných, temporálnych a parietálnych lalokov mozgu. kôra, ako aj s hypotuberóznou oblasťou a retikulárnou formáciou trupu. Limbická kôra je spojená do jedného funkčného systému - limbicko-retikulárneho komplexu. Hlavnou funkciou týchto častí mozgu nie je ani tak poskytovať komunikáciu s vonkajším svetom, ale regulovať tón kôry, pohony a afektívny život. Regulujú zložité, mnohostranné funkcie vnútorných orgánov a behaviorálne reakcie. Limbicko-retikulárny komplex je najdôležitejším integračným systémom tela. Limbický systém je dôležitý aj pri formovaní motivácií. Motivácia (alebo vnútorná motivácia) zahŕňa najzložitejšie inštinktívne a emocionálne reakcie (potravinové, obranné, sexuálne). Limbický systém sa podieľa aj na regulácii spánku a bdenia.
Limbická kôra plní aj dôležitú funkciu čuchu. Vôňa je vnímanie chemikálií vo vzduchu. Čuchový mozogčlovek poskytuje čuch, ako aj organizáciu zložitých foriem emocionálnych a behaviorálnych reakcií. Čuchový mozog je súčasťou limbického systému.
Čuchový mozog sa skladá z dvoch častí – periférnej a centrálnej. Periférny úsek predstavuje čuchový nerv, čuchové bulby, primárne čuchové centrá. Centrálna časť zahŕňa gyrus morského koníka - hipokampus, zubatý a klenutý gyrus.
Aparát čuchového receptora sa nachádza v nosovej sliznici. Prostredníctvom systému nervových vodičov sa informácie z receptorov prenášajú do kortikálnej časti čuchového analyzátora (obr. 8).
Ryža. 8. Čuchový analyzátor (schéma):
1 - čuchový epitel, bipolárne čuchové bunky; 2 - čuchová žiarovka; 3 - čuchový trakt; 4 - primárne čuchové centrá; 5 - vizuálny tuberkul; 6 - kortikálne čuchové centrum; 7 - corpus callosum
Kortikálna oblasť čuchového analyzátora sa nachádza v gyrus cingulate, gyrus morský koník a háčik morského koníka, ktoré spolu tvoria uzavretú prstencovú oblasť. Periférna časť čuchového analyzátora je spojená s kortikálnymi oblasťami oboch hemisfér.
Fyziologický mechanizmus vnímania pachu čuchovým analyzátorom nie je úplne jasný. Existujú dve hlavné hypotézy vysvetľujúce povahu tohto procesu z rôznych pozícií. Podľa jednej z hypotéz k interakcii medzi molekulami odorantu a chemoreceptormi dochádza ako kľúč a zámok, t.j. typ molekuly zodpovedá špeciálnemu receptoru. Ďalšia hypotéza je založená na predpoklade, že molekuly pachovej látky majú určitú oscilačnú vlnu, na ktorú sú „naladené“ čuchové receptory. Molekuly, ktoré majú podobné vibrácie, by mali mať spoločnú vlnu, a teda vydávať podobné pachy.
Pojem „čuchový mozog“ vo vzťahu k ľudskej fyziológii je trochu svojvoľný a neodhaľuje úplne jeho mnohostrannú a univerzálnu funkciu. „Umiestnenie“ centrálneho článku čuchového mozgu do mozgových hemisfér nie je náhodné a je výsledkom obrovskej „informačnej“ úlohy, ktorú čuch zohral v procese evolúcie pri prispôsobovaní sa vonkajšiemu prostrediu a regulácii komplexu. behaviorálne reakcie. Získavanie potravy, výber jedinca opačného pohlavia, starostlivosť o potomstvo, celistvosť územia, organizovanie skupinových spoločenstiev v rámci druhu – všetky tieto každodenné funkcie u mnohých zvierat sa vykonávajú za priamej účasti precízne vybudovaného systému čuchového príjmu a , na základe toho schopnosť množstva zvierat vysielať jemne diferencované špecifické pachových látok- signalizátori-informátori.
Univerzálne formy behaviorálnych reakcií u zvierat, ktoré sa prejavujú v každodennej starostlivosti o biotop, o potomstvo, vytvárajú dojem, že sú obdarené rozumom. Zdanlivá inteligencia je jednoducho výsledkom reakcie na vonkajšie podnety. Samotné tieto podnety a reakcie na ne však dokonale zodpovedajú biologickým potrebám zvierat.
V živote ľudí stratil čuch biologickú informačnú hodnotu, ktorú mal u zvierat. Ľudský čuchový systém je navrhnutý tak, aby vykonával úzku „vlastnú“ funkciu, ako aj akési „nabíjanie“ emócií. Sila vplyvu pachov na emocionálnu sféru, že sú najdôležitejším „potravinovým substrátom emócií“, je v histórii ľudstva dobre známa už od staroveku.
Čuch človeka sa môže líšiť. Tieto variácie sú spravidla zanedbateľné, ale v niektorých prípadoch môže byť ostrosť vône veľmi vysoká (ochutnávači v parfumérskom priemysle).
Keďže čuchový analyzátor zohráva dôležitú úlohu pri regulácii emócií, jeho centrálna časť sa označuje ako limbický systém, obrazne nazývaný „spoločný menovateľ“ mnohých emočných a viscerosomatických reakcií tela.
Veľké hemisféry mozgu
sú najväčšou časťou mozgu. Pokrývajú mozoček a mozgový kmeň. Mozgové hemisféry tvoria približne 78 % celkovej hmoty mozgu.
V procese ontogenetického vývoja organizmu sa z koncového mozgového mechúra nervovej trubice vyvíjajú mozgové hemisféry, preto sa táto časť mozgu nazýva aj telencephalon.
Mozgové hemisféry sa delia na stredná čiara hlboká zvislá štrbina vpravo a ľavá hemisféra. V hĺbke strednej časti sú obe hemisféry prepojené veľkým zrastom - corpus callosum. Každá hemisféra má laloky: čelný, parietálny, časový, tylový.
Laloky mozgových hemisfér sú od seba oddelené hlbokými ryhami. Najdôležitejšie sú tri hlboké ryhy: centrálna (Rolandova), oddeľujúca predný lalok od parietálneho; laterálny (Sylvian), oddeľujúci spánkový lalok od parietálneho a parietálno-okcipitálny, oddeľujúci parietálny lalok od tylového na vnútornom povrchu hemisféry.
Každá hemisféra má horný-laterálny (konvexný) - konvexitný, dolný - bazálny a vnútorný - mediálny povrch. Každý lalok hemisféry má cerebrálne zákruty, ktoré sú navzájom oddelené brázdami. Zhora je hemisféra pokrytá kôrou - tenkou vrstvou šedej hmoty, ktorá pozostáva z nervové bunky.
Cortex- evolučne najmladší útvar centrálnej nervovej sústavy. U ľudí dosahuje svoj najvyšší vývoj. Mozgová kôra má veľký význam pri regulácii vitálnej aktivity tela, pri realizácii zložitých foriem správania a pri tvorbe neuropsychických funkcií.
Pod kôrou je Biela hmota hemisfér, pozostáva z procesov nervových buniek - vodičov. V dôsledku tvorby cerebrálnych konvolúcií sa celkový povrch mozgovej kôry výrazne zvyšuje. Celková plocha hemisférickej kôry je 1200 cm2, pričom 2/3 jej povrchu sa nachádzajú v hĺbke brázd a 1/3 na viditeľnom povrchu hemisfér. Každý lalok mozgu má iný funkčný význam.
Cortex pozostáva zo 4 lalokov, oddelených od seba brázdami. Hlavné drážky oddeľujúce čelné, parietálne a temporálne laloky sú Rolandov a Sylviev.
Laloky mozgovej kôry :
predné (čelný ),
parietálny (parietálny ),
okcipitálny (tylový ),
časové (časový ),
V súvislosti s akciami sa hovorí o lokálnych systémoch mozgu.
LOKÁLNE SYSTÉMY MOZGU:
Okcipitálne oblasti mozgu vykonávať funkcie organizácie vizuálneho vnímania. Primárne zóny okcipitálneho kortexu - elementárne funkcie vízie.
Sekundárne úseky okcipitálneho kortexu - opto-gnostické funkcie.
Časové oblasti mozgu„zodpovedný“ za organizáciu sluchového vnímania. Primárne zóny temporálneho kortexu sú elementárne funkcie sluchu.
Sekundárne zóny temporálneho kortexu – akusticko-gnostické funkcie.
Senzomotorické a premotorické časti mozgu- organizácia pohybov. 1. Postcentrálne časti mozgu - aferentná organizácia pohybov. 2. Premotorické kortexové zóny - eferentná organizácia (programovanie) pohybu.
3. Motorické zóny kôry - motorický analyzátor, realizácia motorického programu. Predné laloky mozgu (prefrontálne oblasti) uskutočňovať reguláciu duševnej činnosti, t.j. regulácia stavov činnosti, dobrovoľné pohyby a činy, kognitívne procesy a emocionálna a osobná sféra, sú vo všeobecnosti zodpovedné za myslenie, intelektuálnu činnosť. Parietálne oblasti mozgu sú reprezentované primárnymi zónami (všetky typy kožno-kinestetickej citlivosti), sekundárnymi zónami (vizuálne-priestorové zobrazenia, predstavy o schéme tela, somatognóza, stereognóza) a terciárne zóny.
čelný lalok zaberá predné časti hemisfér. Od parietálneho laloka je oddelený centrálnym sulcusom a od temporálneho laloku laterálnym sulcusom. Vo prednom laloku sú štyri gyry: jeden vertikálny - precentrálny a tri horizontálne - horný, stredný a dolný frontálny gyrus.
Závity sú od seba oddelené brázdami. Na spodnom povrchu predných lalokov sa rozlišuje priamy a orbitálny gyrus. Priamy gyrus leží medzi vnútorným okrajom pologule, čuchovým žliabkom a vonkajším okrajom pologule.
V hĺbke čuchovej brázdy leží čuchový bulbus a čuchový trakt. Predný lalok človeka tvorí 25 – 28 % kôry, priemerná hmotnosť predného laloku je 450 g.
Funkcia predných lalokov je spojená s organizáciou dobrovoľných pohybov, motorickými mechanizmami reči, reguláciou zložitých foriem správania a myšlienkových procesov. V konvolúciách čelového laloku je sústredených niekoľko funkčne dôležitých centier. Predný centrálny gyrus je "reprezentáciou" primárnej motorickej zóny s prísne definovanou projekciou častí tela. Tvár je „umiestnená“ v dolnej tretine gyrusu, ruka v strednej tretine, noha dovnútra horná tretina. Trup je zastúpený v zadných častiach horného frontálneho gyru. Človek sa teda premieta hore nohami a dole v prednom centrálnom gyre.
Predný centrálny gyrus spolu s priľahlými zadnými úsekmi frontálnych gyri zohráva veľmi funkčne dôležitú úlohu. Je centrom dobrovoľných hnutí. V hĺbke kôry centrálneho gyru z tzv pyramídové bunky -centrálny motorický neurón- začína hlavná motorická dráha - pyramídová, čiže kortikospinálna dráha. Periférne procesy motorických neurónov vychádzajú z kôry, zhromažďujú sa do jedného silného zväzku, prechádzajú centrálnou bielou hmotou hemisfér a vstupujú do mozgového kmeňa cez vnútornú kapsulu; na konci mozgového kmeňa sa čiastočne krížia (prechádzajú z jednej strany na druhú) a potom zostupujú do miecha. Tieto vetvy končia v šedá hmota miecha. Tam prichádzajú do kontaktu s periférnym motorickým neurónom a prenášajú mu impulzy z centrálneho motorického neurónu. Impulzy dobrovoľného pohybu sa prenášajú pozdĺž pyramídovej dráhy.
V zadných úsekoch gyrus frontalis superior sa nachádza aj extrapyramídové centrum kôry, ktoré je anatomicky a funkčne úzko spojené s útvarmi takzvaného extramyramídového systému. Extrapyramídový systém- motorický systém napomáhajúci vykonávaniu ľubovoľného pohybu. Ide o systém „poskytovania“ ľubovoľných pohybov. Ľudský extrapyramídový systém, ktorý je fylogeneticky starší ako pyramídový systém, zabezpečuje automatickú reguláciu „naučených“ motorických úkonov, udržiavanie celkového svalového tonusu, „pripravenosť“ periférneho motorického aparátu na pohyb, redistribúciu svalového tonusu pri pohyboch. Okrem toho sa podieľa na udržiavaní normálneho držania tela.
V zadnej časti stredného frontálneho gyru je frontálne okohybné centrum, ktoré riadi priateľskú, súčasnú rotáciu hlavy a očí (stred rotácie hlavy a očí v opačnom smere). Podráždenie tohto centra spôsobuje otáčanie hlavy a očí opačným smerom. Funkcia tohto centra má veľký význam pri realizácii takzvaných orientačných reflexov, ktoré sú veľmi dôležité pre zachovanie života zvierat.
V zadnej časti je spodný frontálny gyrus motorické centrum reči(Brockov stred).
Prijíma aj čelná kôra mozgových hemisfér Aktívna účasť pri formovaní myslenia, organizovaní cieľavedomej činnosti, dlhodobého plánovania.
temporálny lalok zaberá spodný bočný povrch hemisfér. Spánkový lalok je oddelený od predného a parietálneho laloku bočnou drážkou.
Na hornom bočnom povrchu spánkového laloku sú tri konvolúcie - horný, priemer A nižšie. Horný temporálny gyrus sa nachádza medzi sylvian a superior temporal sulci, stredný gyrus je medzi superior a inferior temporal sulci a gyrus inferior je medzi dolným temporálnym sulcom a transverzálnou cerebrálnou trhlinou. Na spodnom povrchu temporálneho laloku sa rozlišuje dolný temporálny gyrus, laterálny okcipitotemporálny gyrus, hipokampálny gyrus (nohy morského koníka).
Funkcia temporálneho laloku spojené s vnímaním sluchových, chuťových, čuchových vnemov, analýzou a syntézou zvukov reči, pamäťovými mechanizmami. Hlavné funkčné centrum hornej laterálnej plochy spánkového laloku sa nachádza v hornom temporálnom gyrus. Tu je sluchové, alebo gnostické centrum reči (Wernickeho centrum).
V hornom temporálnom gyrus a na vnútornom povrchu temporálneho laloku je oblasť sluchovej projekcie kôry. Oblasť čuchovej projekcie sa nachádza v gyrus hipokampu, najmä v jeho prednom úseku (tzv. hák). Vedľa čuchových projekčných zón sú tiež chuťové.
Časové laloky zohrávajú dôležitú úlohu pri organizácii komplexu mentálne procesy, najmä pamäť.
parietálny lalok zaberá horné bočné plochy hemisféry. Z predného parietálneho laloku, spredu a zboku, je ohraničený centrálnym sulcusom, od temporálneho zdola - laterálnym sulcusom, z tylového - pomyselnou čiarou prechádzajúcou od horného okraja parietálno-okcipitálneho sulcus k spodný okraj pologule.
Na hornom bočnom povrchu parietálneho laloku sú tri zákruty: jedna vertikálna - zadná centrálna a dve horizontálne - horná parietálna a dolná parietálna. Časť gyrus parietalis inferior, ktorá obklopuje zadnú časť laterálneho sulcus, sa nazýva supramarginálna (supramarginálna) a časť obklopujúca horný okraj. temporálny gyrus, nodálna (uhlová) oblasť.
Parietálny lalok, podobne ako predný lalok, tvorí významnú časť mozgových hemisfér. Z fylogenetického hľadiska sa v ňom rozlišuje stará časť - zadný centrálny gyrus, nový - horný parietálny gyrus a novší - dolný parietálny gyrus. Funkcia parietálneho laloku je spojená s vnímaním a analýzou citlivých podnetov, priestorovou orientáciou. V konvolúciách parietálneho laloku je sústredených niekoľko funkčných centier.
V zadnom centrálnom gyre sa centrá citlivosti premietajú s projekciou tela podobnou ako v prednom centrálnom gyre. V dolnej tretine gyrusu sa premieta tvár, v strednej tretine - ruka, trup, v hornej tretine - noha. V gyrus parietalis superior sú centrá, ktoré majú na starosti komplexné typy hlbokej citlivosti: svalovo-kĺbové, dvojrozmerné-priestorové cítenie, zmysel pre váhu a objem pohybu, zmysel pre rozpoznávanie predmetov hmatom.
Kortikálna časť citlivého analyzátora je teda lokalizovaná v parietálnom laloku.
Praxové centrá sa nachádzajú v dolnom parietálnom laloku. Cvičenie sa chápe ako účelové pohyby, ktoré sa zautomatizovali v procese opakovaní a cvičení, ktoré sa rozvíjajú v procese učenia a neustáleho precvičovania počas individuálneho života.
Chôdza, jedenie, obliekanie, mechanické písanie, rôzne druhy pracovná činnosť (napr. pohyby vodiča pri riadení auta, kosenie a pod.) sú praxou.
Prax- najvyšší prejav motorickej funkcie človeka. Vykonáva sa ako výsledok kombinovaných aktivít rôznych území. mozgová kôra.
Okcipitálny lalok zaberá zadné oblasti hemisfér. Na konvexnom povrchu hemisféry nemá okcipitálny lalok ostré hranice, ktoré ho oddeľujú od parietálnych a temporálnych lalokov, s výnimkou hornej časti parietálno-okcipitálneho sulcus, ktorý sa nachádza na vnútornom povrchu hemisféry. oddeľuje temenný lalok od okcipitálneho laloku.
Brázdy a zákruty horného bočného povrchu okcipitálneho laloku sú nestabilné a majú premenlivú štruktúru.
Na vnútornom povrchu okcipitálneho laloku je ostruhová drážka, ktorá oddeľuje klin (trojuholníkový lalok okcipitálneho laloku) od lingválneho gyru a okcipitotemporálneho gyru.
Funkcia okcipitálneho laloku je spojená s vnímaním a spracovaním vizuálne informácie, organizácia zložitých procesov zrakového vnímania. V tomto prípade sa horná polovica sietnice premieta do oblasti klinu, ktorý vníma svetlo z dolných zorných polí; v oblasti lingulárneho gyru je spodná polovica sietnice, ktorá vníma svetlo z horných zorných polí.
ostrov, alebo tzv uzavretý lalok, sa nachádza v hĺbke bočnej brázdy. Ostrovček je oddelený od susedných susedných častí kruhovou drážkou.
Povrch ostrovčeka je svojou pozdĺžnou stredovou drážkou rozdelený na prednú a zadnú časť. Na ostrove sa premieta analyzátor chuti.
Čuchový analyzátor
Nervové bunky, vnímajúce čuchové podráždenia, sa nachádzajú v sliznici horných častí nosnej dutiny. Odtiaľ axóny týchto buniek idú do lebečnej dutiny a vstupujú do čuchových cibuliek. Z nich sú posielané nervové vlákna temporálny lalok (vnútorný povrch) kde sú nervové bunky čuchový analyzátor.
Analyzátor chuti
Tento analyzátor začína v nervových zakončeniach chuťových pohárikov jazyka, ktoré sú v nich zastúpené chuťovými pohárikmi. Nervové vlákna opúšťajúce chuťové poháriky idú do mozgu a končia, podobne ako nervy čuchového analyzátora, v vnútorný povrch temporálny lalok.
corpus callosum- oblúkovitá tenká platnička, fylogeneticky mladá, spája stredové plochy obe hemisféry. Predĺžená stredná časť corpus callosum prechádza vzadu do zhrubnutia a vpredu sa oblúkovito zakrivuje a stáča nadol.
Corpus callosum spája fylogeneticky najmladšie časti hemisfér a zohráva dôležitú úlohu pri výmene informácií medzi nimi.
MOZGOVÝ KMEŇ ALEBO MOZGOVÝ KMEŇ -
tradične rozlišovaný systém oblastí mozgu, ktorý je rozšíreným útvarom, ktorý pokračuje v mieche.
Mozgový kmeň vždy zahŕňa predĺženú miechu, mostík a stredný mozog. Často zahŕňa cerebellum, niekedy diencephalon.
Medulla -
oddelenie mozgu. Existuje aj tradičný názov bulbus (cibuľka, kvôli tvaru tohto oddelenia).
Medulla oblongata vstupuje do mozgového kmeňa.
Vonku sa na ventrálnej (tvárovej) strane nachádzajú pyramídy (obsahujú kortikospinálny trakt - cesta z kôry k motorickým neurónom miechy) a olivy (obsahujú jadrá spodnej olivy spojené s udržiavaním rovnováhy) . Na chrbtovej strane: tenké a klinovité zväzky končiace tuberkulami tenkých a klinovitých jadier (prepínanie informácií o hlbokej citlivosti dolnej a hornej polovice tela), dolnej polovice kosoštvorcovej jamky, čo je dno štvrtej komory a povrazové telá, ktoré ju oddeľujú, alebo dolné končatiny malého mozgu.
Vnútri sú tiež jadrá od VIII do XII (a jedno z jadier VII) hlavových nervov, časť retikulárnej formácie, mediálna slučka a ďalšie vzostupné a zostupné dráhy.
Má tvar zrezaného kužeľa.
Vďaka výskumu vedcov ako R. Magnus a I. F. Klein sa zistilo, že v predĺženej mieche existuje zložitý systém reflexných centier, ktoré zabezpečujú určitú polohu v tele vďaka statickým a staticko-kinetickým reflexom. Tieto reflexy sú v skutočnosti mechanizmami na redistribúciu svalového tonusu takým spôsobom, že je zachované pohodlné držanie tela zvieraťa (posturálno-tonické reflexy) alebo návrat k danému z nepríjemného (rektifikačné reflexy) a rovnováha sa udržiava aj pri zrýchlení (statokinetické reflexy). K realizácii týchto reflexov dochádza za účasti takých kmeňových formácií, ako je retikulárna formácia, červené jadro a vestibulárne jadrá.
Retikulárna formácia - je to útvar, ktorý prebieha z miechy do talamu v rostrálnom (kôrovom) smere. Okrem toho, že sa retikulárna formácia podieľa na spracovaní zmyslových informácií, pôsobí aktivačne na mozgovú kôru, čím riadi činnosť miechy. Prvýkrát je mechanizmus účinku retikulárnej formácie na svalový tonus založil R. Granit: ukázal, že retikulárna formácia je schopná zmeniť aktivitu γ-motorických neurónov, v dôsledku čoho ihaxóny (γ-eferenty) spôsobujú kontrakciu svalových vretien a v dôsledku toho zvýšenie v aferentných impulzoch zo svalových receptorov. Tieto impulzy vstupujúce do miechy spôsobujú excitáciu α-motorických neurónov, čo je príčinou svalového tonusu.
Zistilo sa, že na výkone tejto funkcie retikulárnej formácie sa zúčastňujú dva zhluky neurónov: neuróny retikulárnej formácie mostonosného mosta a neuróny retikulárnej formácie medulla oblongata. Správanie neurónov retikulárnej formácie medulla oblongata podobné správaniu neurónov pri retikulárnej formácii mostíka: spôsobujú aktiváciu a-motorických neurónov ohýbacích svalov, a preto inhibujú aktivitu a-motorických neurónov extenzorových svalov. Neuróny retikulárnej formácie mostíka pôsobia presne opačne, excitujú α-motorické neuróny extenzorových svalov a inhibujú aktivitu α-motorických neurónov flexorových svalov. Retikulárna formácia má spojenie s mozočkom (časť informácií z neho ide do neurónov predĺženej miechy (z jadier korkového a sférického mozočku) a zo stanu do neurónov mosta) a s mozgová kôra, z ktorej prijíma informácie. To naznačuje, že retikulárna formácia je zberačom nešpecifického senzorického toku, ktorý sa pravdepodobne podieľa na regulácii svalovej aktivity.
dôležitý funkčný význam retikulárne alebo sieťový útvar mozgového kmeňa, ktorý sa vyvíja v súvislosti so vznikom systému vagus, vestibulárny a trigeminálnych nervov.
Retikulárna formácia pozostáva z nervových buniek rôznych veľkostí a tvarov, ako aj z hustej siete nervové vlákna, idúce rôznymi smermi a nachádzajúce sa hlavne v blízkosti komorového systému. Retikulárna formácia má primárny význam v kortikálno-subkortikálnych vzťahoch. Nachádza sa na stredných poschodiach. medulla oblongata,hypotalamus, tegmentum sivá hmota, pons.
K retikulárnej formácii pristupujú početné kolaterály zo všetkých aferentných (senzorických) systémov. Prostredníctvom týchto kolaterálov sa akékoľvek podráždenie z periférie, smerujúce do určitých oblastí kôry pozdĺž špecifických dráh nervového systému, dostane aj do retikulárnej formácie. Nešpecifické vzostupné systémy (t.j. dráhy z retikulárnej formácie) zabezpečujú excitáciu mozgovej kôry, aktiváciu jej aktivity.
Spolu so vzostupnými nešpecifickými systémami prechádzajú mozgovým kmeňom aj zostupné nešpecifické systémy, ktoré ovplyvňujú miechové reflexné mechanizmy.
Retikulárna formácia je úzko spojená s limbickým systémom, ako aj s mozgovou kôrou. Vďaka tomu sa vytvára funkčné spojenie medzi vyššími časťami centrálneho nervového systému a trupom. mozog. Tento systém sa nazýva limbicko-retikulárny komplex alebo limbicko-retikulárna os. Tento komplexný štruktúrny a funkčný komplex zabezpečuje integráciu najdôležitejších funkcií, na realizácii ktorých sa podieľajú rôzne časti mozgu.
Je známe, že bdelý stav kôry zabezpečujú špecifické a nešpecifické systémy. Aktivačná reakcia je udržiavaná neustálym prísunom impulzov z receptorov sluchové, vizuálny, čuchové, chuťové a kožné kinestetické analyzátory. Tieto podnety sa prenášajú po špecifických aferentných dráhach do rôznych častí kôry. Od všetkých prihlásených talamus, a následne početné kolaterály k retikulárnej formácii odchádzajú do kôry mozgových hemisfér aferentných dráh, čo zabezpečuje jej vzostupnú aktivačnú činnosť.
Retikulárna formácia zase dostáva impulzy z cerebellum, subkortikálne jadrá, limbický systém ktoré poskytujú emocionálne adaptívne behaviorálne reakcie, motivačné formy správania. Miera zabezpečenia adaptačných nepodmienených reflexných reakcií nešpecifickým systémom u ľudí a zvierat je však rôzna. Ak u zvierat hrajú podkôrové útvary a limbický systém vedúcu úlohu pri napĺňaní životných potrieb organizmu na jeho prežitie v životné prostredie, potom je u človeka v dôsledku dominancie kôry činnosť hlbokých štruktúr mozgu (subkortikálnych útvarov, limbického systému, retikulárneho útvaru) vo väčšej miere ako u zvieraťa podriadená mozgová kôra. Retikulárna formácia hrá dôležitú úlohu pri regulácii svalového tonusu. Svalový tonus je regulovaný dvoma typmi retikulospinálnych dráh. Rýchly vodivý retikulospinálny trakt reguluje rýchle pohyby; pomaly vedúca retikulospinálna dráha - pomalé tonické pohyby.
Retikulárna formácia medulla oblongata sa podieľa na výskyte tuhosti decerebrátu. Keď sa mozgový kmeň pretína nad predĺženou miechou, aktivita neurónov, ktoré majú inhibičný účinok na motorické neuróny miechy, klesá, čo vedie k prudkému zvýšeniu tonusu kostrových svalov.
Funkcie medulla oblongata
Ochranné reflexy (napríklad kašeľ, kýchanie).
Životne dôležité reflexy (napr. dýchanie).
regulácia cievneho tonusu.
Reflexné centrá medulla oblongata:
ochranné (kašeľ, kýchanie atď.)
tónové kontrolné centrá kostrového svalstva na udržanie držania tela.
skrátenie alebo predĺženie času miechového reflexu
trávenie
činnosť srdca
Pons
Pons Varolii (v mene Constanza Varolia), alebo most - časť mozgu, je spolu s mozočkom súčasťou zadného mozgu. patrí do mozgu,
V základoch mosta sú zostupné dráhy: kortikospinálny pyramídový trakt, kortikobulbárny, kortikospinálny trakt.
stredný mozog(lat. Mesencephalon) -časť mozgu, staroveké vizuálne centrum. Zahrnuté v mozgovom kmeni.
funkcie stredného mozgu
1. motor,
2. zmyslové (zrak, sluch),
3. Regulácia žuvania a prehĺtania,

