Veľké hemisféry veľké hemisféry
mozog, párové útvary, spojené corpus callosum do takzvaného telencephalonu. Povrch hemisféry reprezentované početnými veľkými alebo malými hlbokými zákrutami. Existujú laloky: čelné, parietálne, temporálne, ostrovné, okcipitálne. Sivá hmota mozgu sa skladá z nervové bunky- neuróny, tvorí mozgovú kôru a podkôrové gangliá (uzliny). Biela hmota tvorené procesmi neurónov, ktoré tvoria dráhy mozgu.
VEĽKÉ POLOSFÉRYVEĽKÉ HEMISFÉRY mozgu, párové útvary spojené corpus callosum (cm. corpus callosum) v tzv. terminálny mozog. Povrch mozgových hemisfér je reprezentovaný početnými veľkými alebo malými hlbokými zákrutami. Existujú laloky: čelné, parietálne, temporálne, ostrovné, okcipitálne. Sivá hmota mozgu, pozostávajúca z nervových buniek - neurónov, tvorí mozgovú kôru a subkortikálne gangliá (cm. GANGLION)(uzly). Biela hmota je tvorená procesmi neurónov, ktoré tvoria dráhy mozgu.
encyklopedický slovník. 2009 .
Pozrite sa, čo sú „veľké hemisféry“ v iných slovníkoch:
Mozog sú párové útvary, spojené corpus callosum v tzv. terminálny mozog. Povrch mozgových hemisfér je reprezentovaný početnými veľkými alebo malými hlbokými zákrutami. Existujú laloky: čelný, parietálny, temporálny, ostrovný... Veľký encyklopedický slovník
Mozog, párové útvary, spojené corpus callosum v tzv. terminálny mozog. Povrch B. p. je zastúpený početnými. b. alebo m. hlboké zákruty. Existujú laloky: čelné, parietálne, temporálne, ostrovné, okcipitálne. Šedá v ... ... Prírodná veda. encyklopedický slovník
VEĽKÉ HEMISFÉRY MOZGU- vyššie časti mozgu, pozostávajúce z povrchovej vrstvy mozgovej kôry a hlbokých častí subkortexu; pokrýva mozoček a mozgový kmeň. B. p. g. m. delené podľa stredná čiara na pravej a ľavej hemisfére, ktorá v hĺbke ... Psychomotor: Slovník
Severný ľadový oceán na rozdiel od juhu predstavuje úplne stredomorský charakter. Má prirodzené hranice na značnú vzdialenosť a len na troch miestach priamo splýva s vodami Atlantiku a Tichého oceánu ... ...
Severný ľadový oceán má na rozdiel od južného úplne stredomorský charakter. Má prirodzené hranice na značnú vzdialenosť a len na troch miestach priamo splýva s vodami Atlantiku a Tichého oceánu ... ... Encyklopedický slovník F.A. Brockhaus a I.A. Efron
Tiež sa nazýva komparatívna morfológia, je to štúdium vzorcov štruktúry a vývoja orgánov porovnávaním rôzne druhyŽivé tvory. Porovnávacie anatomické údaje sú tradičným základom biologickej klasifikácie. Pod morfológiou... Collierova encyklopédia
Pohľad v reze na mozog dospelého muža. Ľudský mozog (lat. encephalon) je o ... Wikipedia
Veda, ktorá študuje stavbu tela jednotlivé orgány, tkanivá a ich vzťahy v tele. Všetky živé veci sa vyznačujú štyrmi vlastnosťami: rast, metabolizmus, podráždenosť a schopnosť reprodukovať sa. Kombinácia týchto znakov ... ... Collierova encyklopédia
Živočíchy (Mammalia), trieda stavovcov, najviac slávna skupina zvierat, vrátane viac ako 4600 druhov svetovej fauny. Zahŕňa mačky, psy, kravy, slony, myši, veľryby, ľudí atď. V priebehu evolúcie cicavce vykonali najširšie ... ... Collierova encyklopédia
I Medicína Medicínsky systém vedecké poznatky a praktické činnosti zamerané na upevnenie a udržanie zdravia, predĺženie života ľudí, prevenciu a liečbu ľudských chorôb. Na splnenie týchto úloh M. študuje štruktúru a ... ... Lekárska encyklopédia
Ľudský mozog je mimoriadne zložitý systém. Vďaka tomuto orgánu ľudia dosiahli úroveň vývoja, ktorá sa pozoruje teraz. Čo predstavuje?
evolučný vývoj
V modernom školskom kurze biológie sa témy zvažujú od jednoduchých po zložité. Najprv rozprávame sa o bunkách, prvokoch, baktériách, rastlinách, hubách. Neskôr dochádza k prechodu k zvieratám a človeku. Do určitej miery to odráža hypotetický priebeh evolúcie. Ak vezmeme do úvahy napríklad štruktúru červov, je ľahké vidieť, že je oveľa jednoduchšia ako štruktúra ľudí alebo vyšších zvierat. Ale tieto organizmy majú niečo dôležité - ganglion, ktorý vykonáva funkcie mozgu.
predný mozog
Ak niekoho požiadate, aby nakreslil obsah ľudskej lebky, s najväčšou pravdepodobnosťou budú hemisféry schematicky znázornené. Toto je skutočne jedna z najviditeľnejších a najväčších častí. Ale predný mozog obsahuje aj predĺženú miechu. Vo všeobecnosti je ich štruktúra pomerne zložitá. A ak vezmeme do úvahy podrobnejšie členenie, tak môžeme dokonca vymenovať všetky oddelenia predný mozog:
- hippocampus;
- bazálna uzlina;
- veľký mozog.
Samozrejme, existuje ešte podrobnejšie rozdelenie, ale spravidla je to zaujímavé iba pre odborníkov. No pre tých, ktorí si len rozširujú obzory, bude oveľa zábavnejšie zistiť, čo všetky tieto oddelenia robia. Aké sú teda funkcie predného mozgu? A prečo sú rozdiely medzi myslením pravákov a ľavákov?
Funkcie
Predný mozog obsahuje najnovšie vyvinuté časti. A to znamená, že práve vďaka nim má človek vlastnosti, ktoré má. A ak sa diencephalon zaoberá hlavne reguláciou metabolizmu, primitívnymi reflexmi a potrebami, ako aj jednoduchými motorická aktivita, potom sú hemisféry práve tým miestom, kde sa rodia vedomé myšlienky, kde sa učia a ukladajú informácie a vytvára sa niečo nové.
Hemisféry sú tiež podmienene rozdelené do niekoľkých častí-zón: parietálna, čelná, zadná a časová. A tu sú bunky, ktoré sa okrem iného zaoberajú analýzou informácií prichádzajúcich zvonka: zrakové, sluchové, čuchové, chuťové a hmatové centrá.
Najzaujímavejšie je, že z funkčného hľadiska je ľavá a pravá hemisféra odlišná. Samozrejme, existujú prípady, keď bola jedna časť mozgu poškodená, iná prevzala jej úlohy, to znamená, že existuje určitá zameniteľnosť, ale v bežnom prípade môže byť situácia nasledovná: ľavá hemisféra sa zaoberá analýzou intonácie reči inej osoby a tej správnej - interpretáciou významu toho, čo bolo povedané. Preto trochu inak rozmýšľajú ľaváci a praváci, ktorí majú vyvinutejšie odlišné partie.
Medzi funkcie predného mozgu patrí aj pamäť, rôzne reakcie na vonkajšie podnety, plánovanie a budovanie budúcich scenárov a situácií. Nachádza sa tu aj rečové centrum. Tu sa odohráva všetka vyššia nervová činnosť: tvorivosť, úvahy, nápady.
Je tiež celkom zaujímavé, že predný mozog sa aktívne rozvíja nielen v prenatálnom období, ale aj v prvých rokoch života. Každá nová zručnosť a zručnosť, naučené slovo, akékoľvek dôležitá informácia- toto všetko tvorí nové neurónové spojenia. A tento druh karty je jedinečný pre každého človeka.

- Schopnosti myslenia nezávisia od hmotnosti mozgu, ale korelujú s takou hodnotou, ako je počet konvolúcií.
- Rýchlosť signálov medzi neurónmi dosahuje 288 kilometrov za hodinu. S vekom sa toto číslo znižuje.
- Mozog spotrebúva najväčšie množstvo energie ľudské orgány- asi 20 %. Je to obrovské číslo, pretože jeho hmotnosť vo vzťahu k telu je iba 2%. Taktiež pre jeho normálnu činnosť je potrebné dostatočné množstvo tekutín v tele.
- Tvrdenie, že mozog využíva len 10 % svojich zdrojov, je mýtus. V skutočnosti nemôže fungovať toľko centier súčasne, ale tak či onak sú zapojené všetky.
textové polia
textové polia
šípka_nahor
Diencephalon spolu s mozgovým kmeňom je pokrytý zhora a zo strán veľké hemisféry - terminálny mozog. Hemisféry pozostávajú z podkôrových uzlín (bazálnych ganglií) a majú dutiny -. Vonku sú hemisféry zakryté (plášťom).
Bazálne gangliá alebo bazálne gangliá
textové polia
textové polia
šípka_nahor
Bazálna uzlina alebo subkortikálne uzliny (nuclei basales)- útvary sú fylogeneticky staršie ako kôra. Bazálne gangliá dostali svoje meno vďaka tomu, že ležia akoby na dne mozgových hemisfér, v ich bazálnej časti. Patria sem kaudátne a lentikulárne jadrá spojené do striata, plota a amygdaly.
Caudate jadro
textové polia
textové polia
šípka_nahor
Caudate jadro (nucleus caudatus) pretiahnuté v sagitálnej rovine a silne zakrivené (obr. 3.22; 3.32; 3.33). Jeho predná, zosilnená časť - hlavu- je uložený pred talamom, v laterálnej stene predného rohu bočná komora, zozadu sa postupne zužuje a prechádza do chvost. Caudatus nucleus pokrýva zrakový tuberkul spredu, zhora a zo strán.
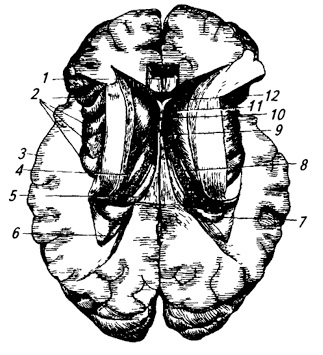
Ryža. 3.22.
1 - nucleus caudatus;
2 - stĺpy oblúka;
3 - epifýza;
4 - horná a
5 - dolný colliculus;
6 - vlákna stredného cerebelárneho stopky;
7 - dráha horného cerebelárneho stopky (vypreparovaná);
8 - jadro stanu;
9 - červ;
10 - sférický,
11 - korok a
13 - zubaté jadro;
12 - kôra cerebelárnych hemisfér;
14 - horný cerebelárny peduncle;
15 - trojuholník vodítka;
16 - vankúš talamu;
17 - vizuálny tuberkul;
18 - zadná komisura;
19 - tretia komora;
20 - predné jadro zrakového pahorku
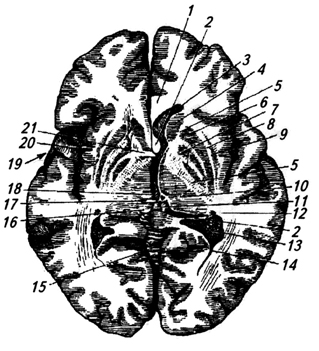
 Ryža. 3.32.
Ryža. 3.32. Ryža. 3.32. Mozog - horizontálny rez laterálnymi komorami:
1 - corpus callosum;
2 - ostrovček;
3 - kôra;
4 - chvost jadra caudate;
5 - oblúk;
6 - zadný roh laterálnej komory;
7 - hipokampus;
8 – choroidný plexus;
9 - interventrikulárny otvor;
10 - priehľadná priečka;
11 - hlava nucleus caudate;
12 – predný roh bočná komora
Jadro v tvare šošovice
textové polia
textové polia
šípka_nahor
Jadro v tvare šošovice (nucleus lentiformis) nachádza sa mimo vizuálneho kopca, na úrovni ostrova. Tvar jadra je blízky trojstennej pyramíde so základňou otočenou von. Jadro je jasne rozdelené vrstvami bielej hmoty na tmavšiu bočnú časť - škrupina a mediálne bledá guľa, pozostávajúce z dvoch segmentov: vnútorného a vonkajšieho (obr. 3.33; 3.34).
 Ryža. 3.33.
Ryža. 3.33. Ryža. 3.33. Horizontálny rez mozgovými hemisférami na úrovni bazálnych ganglií:
1 - corpus callosum;
2 - klenba;
3 - predný roh laterálnej komory;
4 - hlava nucleus caudate;
5 - vnútorná kapsula;
6 - škrupina;
7 - bledá guľa;
8 - vonkajšia kapsula;
9 - plot;
10 - talamus;
11 - epifýza;
12 - chvost jadra caudate;
13 - choroidný plexus laterálnej komory;
14 - zadný roh laterálnej komory;
15 - cerebelárna vermis;
16 - kvadrigemina;
17 - zadná komisura;
18 - dutina tretej komory;
19 - jama bočnej brázdy;
20 - ostrovček;
21 - predná komizúra
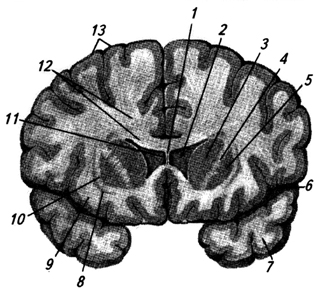 Ryža. 3.34.
Ryža. 3.34. Ryža. 3.34. Predný rez cez mozgové hemisféry na úrovni bazálnych ganglií:
1 - corpus callosum;
2 - laterálna komora;
3 - caudate nucleus (hlava);
4 - vnútorná kapsula;
5 - jadro šošovkovitého tvaru;
6 - bočná brázda;
7 - temporálny lalok;
8 - plot;
9 - ostrovček;
10 - vonkajšia kapsula;
11 - priehľadná priečka;
12 - vyžarovanie corpus callosum;
13 - mozgová kôra
Shell
textové polia
textové polia
šípka_nahor
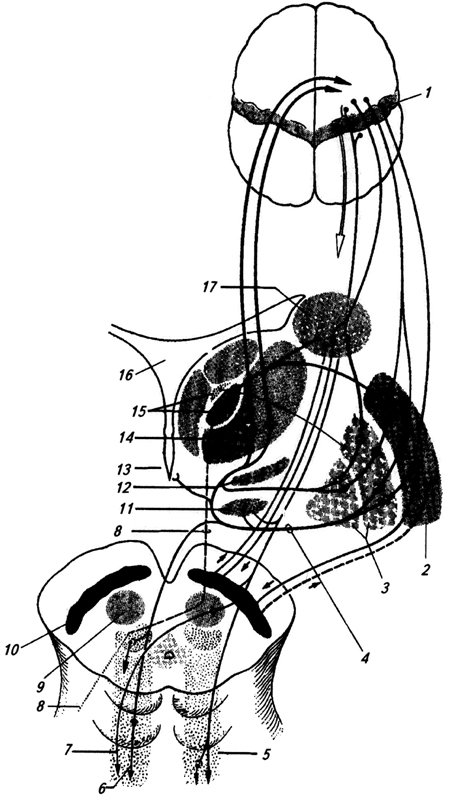 Ryža. 3.35.
Ryža. 3.35. Shell (putamen) geneticky, štrukturálne a funkčne blízko k jadru caudate.
Oba tieto útvary majú zložitejšiu štruktúru ako bledá guľa. Pristupujú k nim vlákna najmä z mozgovej kôry a talamu (obr. 3.35).
Ryža. 3.35. Aferentné a eferentné spojenia bazálnych ganglií:
1 - precentrálny gyrus;
2 - škrupina;
3 - vonkajšie a vnútorné segmenty bledej gule;
4 - lentikulárna slučka;
5 - retikulárna formácia;
6 - retikulospinálny trakt,
7 - rubrospinálny trakt;
8 - cerebelárno-talamický trakt (z dentátneho jadra mozočka);
9 - červené jadro;
10 - čierna látka;
11 - subtalamické jadro;
12 - Zona incerta;
13 - hypotalamus;
14 - ventrolaterálna,
15 - intralaminárne a centromedické jadrá talamu;
16 - III komora;
17 - kaudátne jadro
bledá guľa
textové polia
textové polia
šípka_nahor
Globus pallidus (globus pallidus) je spojený hlavne s vedením impulzov po mnohých zostupných dráhach do nižších mozgových štruktúr - červené jadro, čierna látka atď. Vlákna z neurónov pallidus smerujú do rovnakých jadier talamu, ktoré sú spojené s mozočkom. Z týchto jadier vedú početné cesty do mozgovej kôry.
Bledá guľa dostáva impulzy z nucleus caudate a putamen.
striatum ( corpus striatum) (striatum), ktoré spája kaudátne a lentikulárne jadrá, patrí medzi eferentné extrapyramídový systém. Dendrity striatálnych neurónov sú pokryté početnými tŕňmi. Ukončujú vlákna z neurónov kôry, talamu a čiernej hmoty (obr. 3.35). Striatálne neuróny zase posielajú axóny do intralaminárneho, predného a laterálneho jadra talamu. Z nich idú vlákna do kôry, a tak sa medzi kortikálnymi neurónmi a striatom uzatvára spätná väzba.
V procese fylogenézy boli tieto jadrá postavené nad jadrami stredného mozgu. Striatum, ktoré prijíma impulzy z talamu, sa podieľa na vykonávaní takých zložitých automatických pohybov, ako je chôdza, lezenie, beh. V jadrách striata sú oblúky najzložitejších nepodmienených uzavreté, t.j. vrodené, reflexy. Extrapyramídový systém je fylogeneticky starší ako pyramídový systém. U novorodenca toto ešte nie je dostatočne vyvinuté a impulzy sa dostávajú do svalov z podkôrových ganglií cez extrapyramídový systém. V dôsledku toho sa pohyby dieťaťa v prvých mesiacoch života vyznačujú zovšeobecňovaním, nediferenciáciou. S vývojom mozgovej kôry rastú axóny ich buniek k bazálnym gangliám a ich činnosť začína byť regulovaná kôrou. Subkortikálne gangliá sú spojené nielen s motorickými reakciami, ale aj s vegetatívne funkcie- tieto sú najvyššie subkortikálne centrá autonómna nervová sústava.
amygdala
textové polia
textové polia
šípka_nahor
amygdala (corpus amygdaloideum) (amygdala) - akumulácia buniek v bielej hmote spánkového laloku. S pomocou predná komisura spája sa s telom rovnakého mena na druhej strane. Amygdala prijíma impulzy z rôznych aferentných systémov vrátane čuchového systému a súvisí s emocionálnymi reakciami (obr. 3.36).
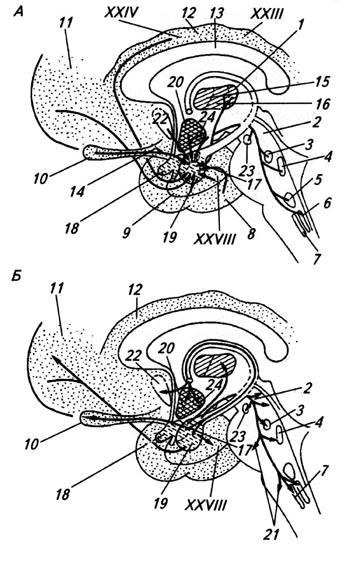 Ryža. 3.36.
Ryža. 3.36. Ryža. 3.36. Štruktúry mozgu spojené s amygdalou: aferentné (A) a eferentné (B) spojenia amygdaly:
1 - jadrá talamu;
2 - periakveduktálna šedá hmota;
3 - parabrachiálne jadro;
4 - modrá škvrna;
5 - šijacie jadrá;
6 - jadro jednej cesty;
7 - dosálny nucleus X nerv;
8 - časová kôra;
9 - čuchová kôra;
10 - čuchová žiarovka;
11 - predná kôra;
12 - cingulárny gyrus;
13 - corpus callosum;
14 - čuchové jadro;
15 - predno-ventrálna a
16 - dorzomediálne jadrá talamu;
17 - centrálna,
18 - kortikálna a
19 - bazolaterálne jadro amygdaly;
20 - hypotalamus;
21 - retikulárna formácia;
22 - priečka;
23 - čierna látka;
24 - ventromediálne jadro hypotalamu; XXIII, XXIV, XXVIII - kortikálne polia
Mozgové hemisféry sú najväčšie oblasti mozgu. Ľudská hemisféra veľký mozog dostali maximálny vývoj v porovnaní so zvyškom častí, čo do značnej miery odlišuje mozog človeka a zvieraťa. vľavo a pravá hemisféra mozgu sú od seba oddelené pozdĺžnou trhlinou prechádzajúcou pozdĺž stredovej čiary. Ak sa pozriete na povrch mozgu zhora a zboku, môžete vidieť štrbinovitú priehlbinu, ktorá začína 1 cm dozadu od stredného bodu medzi predným a zadným pólom mozgu a ide hlbšie. Toto je centrálna (Rolandova) brázda. Pod ním prechádza pozdĺž bočného povrchu mozgu druhá veľká štrbinovo-laterálna (Sylviánska) drážka. Funkcie hemisféra predný mozog - téma článku.
1 106654
Fotogaléria: Funkcie mozgovej hemisféry predného mozgu
Laloky mozgu
Veľké hemisféry sú rozdelené na laloky, ktorých názvy sú dané kosťami, ktoré ich kryjú: Čelné laloky sa nachádzajú pred Rolandovou a nad Sylviovou drážkou.
Parietálny lalok leží za centrálnou a nad zadnou časťou laterálneho sulcus; siaha späť do parietookcipitálneho sulcus - medzery, ktorá sa oddeľuje parietálny lalok z okcipitálu, ktorý tvorí zadnú časť mozgu.
Spánkový lalok je oblasť pod Sylvian sulcus a hraničí za okcipitálnym lalokom.
Keďže mozog rýchlo rastie už pred narodením, mozgová kôra začína zväčšovať svoj povrch a vytvárať záhyby, čo vedie k vytvoreniu charakteristického vzhľad mozog pripomínajúci Orech. Tieto záhyby sú známe ako konvolúcie a drážky, ktoré ich oddeľujú, sa nazývajú brázdy. Určité ryhy u všetkých ľudí sa nachádzajú na rovnakom mieste, preto sa používajú ako orientačné body na rozdelenie mozgu na štyri laloky.
Vývoj zákrutov a brázd
V 3.-4. mesiaci vývoja plodu sa začínajú objavovať brázdy a zákruty. Až do tohto bodu zostáva povrch mozgu hladký, ako mozgy vtákov alebo obojživelníkov. Vytvorenie zloženej štruktúry poskytuje zväčšenie povrchovej plochy mozgovej kôry v obmedzenom objeme lebky. Rôzne oblasti mozgovej kôry vykonávajú určité, vysoko špecializované funkcie. Mozgovú kôru možno rozdeliť do nasledujúcich oblastí:
Motorické zóny – iniciujú a riadia pohyby tela. Primárna motorická oblasť riadi dobrovoľné pohyby na opačnej strane tela. Priamo pred motorickou oblasťou kôry je takzvaná premotorická kôra a tretia oblasť - prídavná motorická oblasť - leží na vnútornom povrchu predného laloku.
Senzorické oblasti mozgovej kôry vnímajú a sumarizujú informácie zo zmyslových receptorov v celom tele. Primárna somatosenzorická oblasť dostáva informácie z opačnej strany tela vo forme impulzov zo zmyslových receptorov pre dotyk, bolesť, teplotu a polohu kĺbov a svalov (proprioceptívne receptory).
Povrch ľudského tela má svoje „zastúpenia“ v zmyslových a motorických oblastiach mozgovej kôry, ktoré sú určitým spôsobom organizované. Kanadský neurochirurg Wilder Penfield, ktorý sa venoval praxi v 50. rokoch, vytvoril akúsi mapu senzorické zóny mozgová kôra, ktoré vnímajú informácie z rôznych častí tela. V rámci výskumu robil pokusy, pri ktorých ponúkal človeku pod lokálna anestézia opísať svoje pocity v momente, keď stimuloval určitých oblastiach povrchu mozgu. Penfield zistil, že stimulácia postcentrálneho gyru vyvolala hmatové vnemy v špecifických oblastiach na opačnej strane tela. Iné štúdie ukázali, že objem motorickej kôry, ktorá je zodpovedná za rôzne oblasti ľudského tela, závisí viac od úrovne zložitosti a presnosti vykonávaných pohybov ako od sily a objemu. svalová hmota. Mozgová kôra sa skladá z dvoch hlavných vrstiev: sivá hmota - tenká vrstva nervových a gliových buniek s hrúbkou asi 2-A mm a biela hmota, ktorá je tvorená nervovými vláknami (axónmi) a gliovými bunkami.
Povrch mozgových hemisfér je pokrytý vrstvou šedá hmota, ktorej hrúbka sa v rôznych častiach mozgu pohybuje od 2 do 4 mm. Šedú hmotu tvoria telá nervových buniek (neurónov) a gliových buniek, ktoré plnia podpornú funkciu. Vo väčšine mozgovej kôry možno pod mikroskopom nájsť šesť samostatných vrstiev buniek.
Neuróny mozgovej kôry
- Pyramídové bunky dostali svoje meno vďaka tvaru tela neurónu, ktorý pripomína pyramídu; ich axóny (nervové vlákna) vychádzajú z mozgovej kôry a prenášajú informácie do iných častí mozgu.
- Nepyramídové bunky (všetky ostatné) sú určené na vnímanie a spracovanie informácií z iných zdrojov.
Hrúbka šiestich vrstiev buniek, ktoré tvoria mozgovú kôru, sa značne líši v závislosti od oblasti mozgu. Nemecký neurológ Korbinian Brodmann (1868-191) skúmal tieto rozdiely farbením nervových buniek a ich skúmaním pod mikroskopom. Výsledkom Brodmannovho vedeckého výskumu bolo rozdelenie mozgovej kôry na 50 samostatných častí na základe určitých anatomických kritérií. Následné štúdie ukázali, že takto identifikované „Brodmannove polia“ zohrávajú špecifickú úlohu. fyziologickú úlohu a majú jedinečné spôsoby interakcie.
Mozog (pokračovanie)
Najväčšiu veľkosť a zložitosť u cicavcov dosahuje predný alebo terminálny mozog (telencephalon), pozostávajúci z dvoch mozgových hemisfér (hemispheri cerebri). Vraj hemisféry vznikli najmä (a možno výlučne) v súvislosti s čuchovým príjmom. Vône v živote vyšších primátov, vrátane človeka, toľko neznamenajú. Avšak na viac skoré štádia evolúcia, až po predkov stavovcov, bol čuch hlavným kanálom, ktorým zvieratá dostávali informácie o svete okolo seba. Preto je celkom prirodzené, že čuchové centrá mozgu slúžili ako základ, na ktorom sa následne vyvíjali zložité štruktúry. nervové mechanizmy. Už v raných štádiách evolúcie tetrapodov sa hemisféry menia na veľké a dôležité centrá korelácie zmyslových signálov. V čase, keď sa objavili cicavce, silne rozšírený povrch hemisfér sa stal dominantným asociačným centrom, miestom lokalizácie vyššej duševnej aktivity. U rôznych predstaviteľov triedy sa pomer hmotnosti hemisfér predného mozgu k hmotnosti celého mozgu líši: u ježka ( Erinaceus europaeus) je to 48% pre bielkoviny ( Sciurus vulgaris) - 53 %, vlk ( canis lupus) – 70 %, na spoločnom boku ( Delphinus delphis) - 75%, u väčšiny primátov - 75-80%, u ľudí - asi 85%. U vtákov veľké hemisféry približne zodpovedajú hmotnosti zvyšku mozgu alebo sú pod ním, niekedy aj niekoľkokrát. Napokon, o mimoriadnom význame mozgových hemisfér svedčí fakt, že ich zničenie vedie k úplnému funkčnému zlyhaniu cicavca.
Zospodu priliehajú k prednej časti hemisfér čuchové bulby (bulbi olfactorii). Tieto útvary sú najviac vyvinuté u zvierat s dobrým čuchom a v čisto vodných formách sú značne obmedzené. Podľa rozdielu v ich vývoji sa rozlišuje niekoľko typov štruktúry mozgu. U vačkovcov, hmyzožravcov, bezzubých, mäsožravcov, hlodavcov a niektorých ďalších sú čuchové cibuľky veľké a dobre vyčnievajú pri pohľade na mozog zhora. Tento typ mozgu s dokonalým vyvinutím čuchových lalokov sa nazýva makrosmatický. U plutvonožcov, sirén a mnohých primátov sú cibule slabo vyvinuté; Tieto zvieratá majú mikrosmatický mozog. Napokon u veľrýb tzv. anosmatický mozog so zníženými čuchovými bulbmi. Kedysi sa predpokladalo, že schopnosť rozlišovať medzi chemickými signálmi u veľrýb a delfínov sa úplne stratila, no ukázalo sa, že to nie je celkom pravda.
Povrchové vrstvy hemisfér predného mozgu cicavcov tvoria pálium, čiže mozgový fornix (pallium). Horná vrstva, pozostávajúca z tiel neurónov a nemäsitých nervové vlákna, sa nazýva kôra (cortex cerebri) a je sivou hmotou oblúka. Telá neurónov sú v kôre umiestnené vo vrstvách a tvoria akési mriežkové štruktúry. Táto organizácia mozgu vám umožňuje priestorovo zobraziť vonkajší svet na základe informácií prichádzajúcich zo zmyslov. Štruktúry obrazovky sú charakteristické pre najdôležitejšie mozgové centrá cicavcov, zatiaľ čo u iných stavovcov sú menej časté, hlavne vo zrakových centrách. Pod kôrou je vrstva myelinizovaných procesov neurónov - biela hmota fornixu. Nervové vlákna bielej hmoty tvoria vodivé zväzky, spájkujúce hemisféry - cerebrálne komisúry. Aj keď je hlavná časť šedej hmoty sústredená v kôre, je prítomná aj v podkôrovej vrstve, kde ju predstavujú relatívne malé zhluky – jadrá. Patria sem striatálne telieska (corpora striata), ktoré ležia pod dutinou bočnej komory a dostali svoje meno vďaka nervovým vláknam, ktoré ich križujú. Striatum pod kontrolou kôry vykonáva funkciu regulácie stereotypných, automatizovaných reakcií - nepodmienených reflexov.
Schémy odrážajúce progresívny vývoj hemisfér predného mozgu. Bočný pohľad na hemisféru s čuchovým bulbom. Rôzne oddelenia, ktoré sa navzájom cytologicky líšia, sa farbia inak.
A - hemisféra je len čuchový lalok.
B - rozlišuje sa dorzálny úsek - archipallium (= hippocampus) a ventrálny úsek - bazálne jadro (striatum).
B - bazálne jadrá sa presunuli do vnútornej oblasti hemisféry.
G - objaví sa malá oblasť, čo je neopálium.
E - paleopallium je zatlačené späť na mediálny povrch hemisféry, ale neopálium je stále skromné a výrazne vyvinuté čuchové oblasti zostávajú pod čuchovým žliabkom.
E - primitívna čuchová oblasť je zachovaná iba vo ventrálnej oblasti a neopálium dosahuje mimoriadne silný vývoj. (Podľa Romera a Parsonsa, 1992.) Fornix mozgu sa začal formovať aj u rýb. Vďaka progresívnemu vývoju vône zmyslový systém majú paleopallium alebo starodávnu klenbu (paleopallium), úplne pokrývajúcu malé hemisféry. V štádiu laločnatých rýb sa v chrbtovej časti hemisfér, bližšie k osi tela, objavuje archipallium, čiže stará klenba (archipallium). U obojživelníkov a primitívnych plazov sa ďalej rozvíja, v dôsledku čoho je paleopálium odsunuté nabok a zachováva sa iba pozdĺž bočného povrchu hemisfér. Paleopallium má zároveň naďalej prevažne čuchový charakter a v najvyšších štádiách vývoja tvorí čuchové laloky hemisférickej kôry. Archipallium je do určitej, aj keď malej miery korelačným centrom, prijímajúcim vzostupné vlákna z diencephalonu, ako aj vlákna z bulbu olfactorius a laloku čuchu; zdá sa, že súvisí aj s emocionálnym správaním. Nervová dráha z tejto oblasti do hypotalamu je hlavným prvkom zväzku vlákien, ktorý sa u cicavcov nazýva fornix.
U obojživelníkov sa po prvýkrát objavuje rudiment neopália, čiže nová klenba (neopallium). U plazov už neopálium tvorí malú oblasť medzi antickou a starou klenbou. Od samého začiatku svojho vývoja bola táto oblasť asociatívnym centrom, ktoré podobne ako bazálne jadrá prijíma vlákna, ktoré naň prepínajú zmyslové signály z mozgového kmeňa a naopak, prenášajú príkazy priamo do motorických stĺpcov.
Pri monotrémoch je neopálium stále vklinené medzi paleopalium zvonku a archipalliom zvnútra. U vačkovcov prerastá cez strechu a bočné steny hemisfér. Súčasne je archipallium vytlačené na mediálny povrch a paleopallium je obmedzené na ventrolaterálnu časť hemisféry, ktorá sa nachádza pod nosovou ryhou (fissura rhinalis) - ryha, ktorá predstavuje hranicu medzi čuchovým a non- čuchové oblasti kôry. V placentárnej v dôsledku ďalšej komplikácie a expanzie neopália dosahujú hemisféry takú veľkosť, že prevyšujú objem zvyšku mozgu dohromady. Hemisféry rastú dozadu a do strán a postupne pokrývajú diencephalon, stredný mozog a časť cerebellum. Spárované komory a staré štruktúry zodpovedné hlavne za čuch (čuchové cibuľky, staré vrstvy fornixu a s nimi spojené nervové zväzky a jadrá) sú v placentárnom mozgu odsunuté a deformované. Paleopallium je teda zachované na ventrálnom povrchu hemisféry vo forme malej čuchovej oblasti nazývanej hruškovitý lalok (lobus piriformis) a archipallium je zvinuté v hĺbke záhybu spánkového laloka do zvitok nazývaný hipokampus. Nadradenosť mozgových hemisfér nad ostatnými časťami mozgu je badateľná u všetkých cicavcov, no obzvlášť výrazná je u pokročilých foriem, ako je človek. Funkčne dominujú aj hemisféry. Na rozdiel od cicavcov sa komplikácia predného mozgu u vtákov prejavuje hlavne rastom bazálnych jadier (nuclei basales), a nie zostávajúcim tenkým oblúkom. 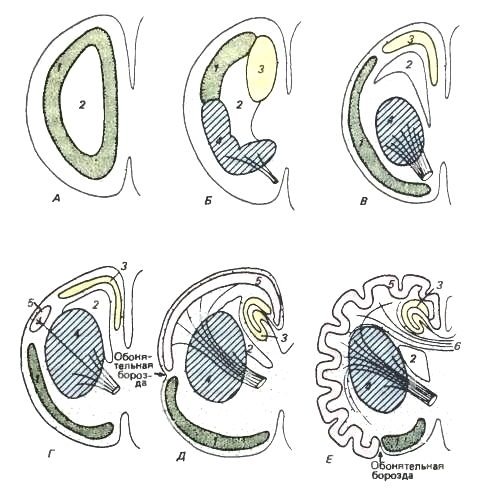
Schematické rezy ľavou hemisférou predného mozgu. Kódy farieb sú rovnaké ako na predchádzajúcom obrázku.
1 - paleopalium; 2 - laterálna komora; 3 - archipallium; 4 - bazálne jadrá; 5 - neopálium; 6 - corpus callosum.
A je primitívne štádium. Hemisféra je v skutočnosti čuchový lalok. Vo vnútri mozgu sa nachádza slabo diferencovaná šedá hmota.
B - štádium pozorované u moderných obojživelníkov. Sivá hmota sa ešte nachádza ďaleko od vonkajšieho povrchu, ale už sa delí na paleopallium (= čuchový lalok), archipallium (= hippocampus) a bazálne jadrá (= striatum). Ten naberá na význame asociatívne centrum, ktoré má aferentné a eferentné spojenia s talamom (zobrazené ako čiary symbolizujúce narezané zväzky vlákien).
B - progresívnejšie štádium, v ktorom sa bazálne jadrá ponoria do hemisféry, zatiaľ čo časti kôry sa trochu posunuli smerom von.
G - štádium, v ktorom sa nachádzajú pokročilé plazy. Objaví sa neopálium.
D - štádium primitívneho cicavca. Neopálium sa zvýšilo. Má rozsiahle spojenie s mozgovým kmeňom. Archipallium na mediálnom povrchu hemisféry sa zabaľuje ako hipokampus. Paleopalium je stále vysoko vyvinuté.
E - štádium vysoko organizovaného cicavca. Neopálium rastie extrémne veľké a zhromažďuje sa do záhybov. Paleopalium zaberá obmedzenú ventrálnu oblasť, ktorá predstavuje hruškovitý lalok. Vyvíja sa corpus callosum - silný most spájajúci neopálové oblasti dvoch hemisfér. (Podľa Romera a Parsonsa, 1992.) Neopáliová kôra sa nazýva nová kôra alebo neokortex (neokortex). U cicavcov slúži ako centrum vyššieho (podmieneného reflexu) nervová činnosť koordinácia práce iných častí mozgu. Odtiaľ sa vysielajú impulzy do rôznych orgánov a tkanív tela a vykonáva sa tu aj regulácia. fyziologické procesy podľa podmienok prostredia. Práve nová kôra hromadí stopy jednotlivých vzruchov a ich kombinácií, v dôsledku čoho sa obohacuje pracovná pamäť, ktorá poskytuje možnosť výberu optimálnych riešení v nových situáciách. Tieto rozhodnutia sú často novými kombináciami predtým známych prvkov správania, ale existuje aj vývoj a konsolidácia nových možností konania. Ako sa vyvíja, nová kôra preberá nielen funkcie korelačného a asociatívneho centra pre novovznikajúce typy vyššej nervovej aktivity, ale začína vykonávať aj mnohé funkcie, ktoré predtým patrili k centrám mozgového kmeňa a bazálnych jadier. Staroveké centrá, ktoré ovládajú inštinktívne akty, sa zároveň nelikvidujú, ale iba podliehajú vyššej kontrole.
V súvislosti s vývojom neokortexu stráca strecha stredného mozgu svoj bývalý význam, zostáva len reflexným a prenosovým centrom. Sluchové a iné somatické zmyslové impulzy sa prenášajú dopredu do talamu, väčšina zrakových vlákien je tu prerušená a všetky tieto signály z talamu sa prenášajú do hemisfér po mocných nervových zväzkoch. Podobné talamické spojenia s bazálnymi jadrami vznikli aj u málo organizovaných skupín stavovcov a boli najviac vyvinuté u vtákov. Na rozdiel od vtákov u cicavcov prevažná časť vlákien prechádza cez striatum a cezeň a rozchádza sa na povrch novej kôry. Tak k nemu prúdi kompletný súbor senzorických dát, na základe ktorých sa v kôre robia zodpovedajúce motorické „rozhodnutia“.
Ako už bolo spomenuté, niektoré signály sa prenášajú z kôry do mozočka cez mostík a poskytujú potrebné regulačné účinky. Neokortex má tiež spojenie so striatom a dokonca aj s hypotalamom - a teda s autonómnym nervový systém. Väčšina motorických príkazov je však smerovaná pozdĺž pyramídovej dráhy (tractus corticospinalis) - špeciálne nervový zväzok ktorý priamo, bez prepínania, prechádza z mozgovej kôry cez stredný mozog do somatických motorických oblastí mozgového kmeňa a miecha. Zároveň sa vlákna laterálnej časti tejto dráhy krížia a inervujú opačná strana telo (t.j. ľavé vlákno inervuje pravá strana telo a naopak) a ventrálne vlákna zostávajú spojené s ich stranou tela. Pyramídová dráha je prítomná len u cicavcov, čo jasne dokazuje dominantné postavenie novej kôry u nich. Najväčší rozvoj dosahuje táto štruktúra u opíc a najmä u ľudí, pričom hrá dôležitú úlohu pri bipedálnej lokomócii. U vačkovcov siahajú pyramídové axóny len po hrudný a pri jednoprejazde pyramídová dráha úplne chýba. 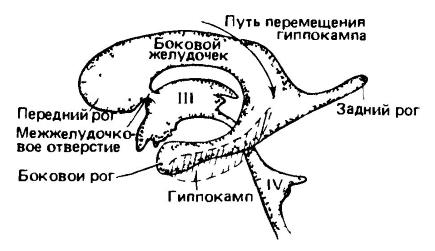
Komory ľudského mozgu; bočný pohľad z ľavej strany. Komory sú zobrazené ako odliatok, zatiaľ čo mozgové tkanivá nie sú zobrazené. S rastom hemisféry predného mozgu sa bočná komora šíri späť s tvorbou zadného rohu v okcipitálnom laloku a v jeho bočnej časti - nadol a dopredu s tvorbou bočného rohu v okcipitálnom laloku. temporálny lalok. Podobné výrastky smerujúce tam a späť viedli k zmenám v umiestnení rôzne časti mozog. Hipokampus, ktorý sa vyvinul v dorzálnej polohe na strednom povrchu hemisféry, sa u vysoko vyvinutých cicavcov posunul späť a dole do ventrálnej polohy. (Podľa Romera a Parsonsa, 1992.) Keďže neokortex je tenká vrstva vrstveného bunkového materiálu, pod ktorým leží biela vláknitá hmota mozgu, jednoduché zväčšenie objemu hemisfér nemôže spôsobiť proporcionálne rozšírenie kôry. Zároveň sa v pokročilých formách môže plocha kôry výrazne zväčšiť v dôsledku jej skladania. Takto vytvorené záhyby sa nazývajú konvolúcie (gyri) a hlboké medzery medzi nimi sa nazývajú brázdy (sulci). Obidve obsahujú spoločné morfologické zložky. V najjednoduchšom prípade je jedna hlboká Sylviova ryha oddeľujúca predný lalok (lobus frontalis) od spánkového laloku (lobus temporalis). Potom sa nad a pred Sylvian sulcus objaví priečny Rolandov sulcus, ktorý oddeľuje čelný lalok od parietálneho (lobus parietalis) zhora. U primátov oddeľuje malý zadný diel priečna brázda okcipitálny lalok(lobus occipitalis). Okrem hlavných brázd sa vytvára mnoho ďalších; ich počet je vysoký najmä u primátov a zubatých veľrýb. Predtým sa verilo, že brázdy v niektorých prípadoch označujú morfologické hranice zodpovedajúce určitým oblastiam kôry. Ďalšie štúdie však ukázali absenciu pevného vzťahu medzi distribúciou skladania a konštrukčná jednotka kôra (okrem nosovej ryhy a do určitej miery aj centrálnej ryhy u primátov, o ktorej bude reč neskôr). Je pozoruhodné, že skladanie kôry sa vyvinulo v niekoľkých evolučných kmeňoch cicavcov celkom nezávisle. U relatívne primitívnych cicavcov, ako sú monotrémy, vačkovce a niektoré placenty (hmyzožravce, netopiere, hlodavce, zajacovité), je kôra vyvinutá skromnejšie a má hladký povrch. 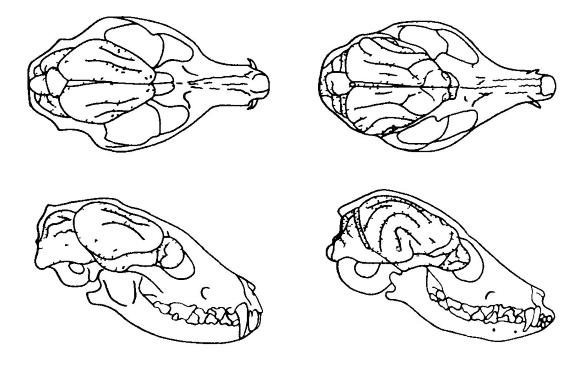
Umiestnenie mozgu v lebke fosílie a živého psa. Dochádza k výraznému nárastu veľkosti a komplikácií mozgu, najmä hemisfér predného mozgu. Hesperocion ( Hesperocyon gregarius) (vľavo) je oligocénna forma, ktorá žila približne pred 30 miliónmi rokov. Fenech ( Vulpes zerda) (napravo) - moderná forma podobné veľkosti. (Podľa Romera a Parsonsa, 1992.) Sivá hmota neokortexu sa vyznačuje zložitou histologickou štruktúrou. U placentárnych cicavcov sa rozlišuje 6 vrstiev buniek ležiacich nad sebou a vlákna zasahujúce medzi ne; to výrazne odlišuje neokortex od zvyšných častí kôry paleopalia a archipallia, kde možno rozlíšiť iba 2 až 4 bunkové vrstvy. Podľa doterajších odhadov sa u cicavcov s obzvlášť veľký mozog, počet buniek novej kôry môže dosiahnuť miliardy.
Biela hmota, ktorá sa nachádza pod šedou, okrem vejára spojení idúcich z kôry do základných častí mozgu a späť, zahŕňa obrovské množstvo prepletených priečnych vlákien spájajúcich rôzne oblasti samotnej kôry. Takto vytvorená komizúra je natiahnutá dozadu (podľa priebehu rastu hemisfér) a rozdelená na dve platničky zrastené pozdĺž zadného okraja. Spodná, tenšia a predným okrajom vychýlená smerom nadol, je oblúk (fornix), komisúra archipallium cortex (t.j. hipokampus). Horná, hrubšia, horizontálne umiestnená komisúra patrí do novej kôry a nazýva sa corpus callosum (corpus callosum). Táto formácia umožňuje spojiť pamäť oboch hemisfér a výrazne zvyšuje schopnosť mozgu učiť sa. Corpus callosum je vďaka výraznému rozvoju novej kôry prítomný len v placentároch, sú oň zbavené monotrémy a vačkovce. Okrem toho majú všetky cicavce prednú komisuru (commissura anterior), ktorá spája čuchové oblasti kôry. 
Vrstvené usporiadanie nervových buniek v kôre telencephalon cicavec (podľa Naumova a Kartasheva, 1979.) Komplexný systém „vodičov“ spájajúcich všetky časti kôry naznačuje, že sivá hmota je v zásade jednotné vzdelávanie, ktorého všetky časti majú rovnaké schopnosti na realizáciu akýchkoľvek funkcií mozgových hemisfér. Do určitej miery je to pravda: experimenty ukazujú, že u laboratórnych zvierat je možné zničiť významnú časť novej kôry bez toho, aby došlo k trvalému poškodeniu ich normálna činnosť. Dôkazy o zranení a morbídnej zmene potvrdzujú, že to platí aj pre ľudský mozog. Zároveň je jasné, že určité oblasti kôry sú normálne spojené s výkonom celkom špecifických funkcií. Vyššie sme spomenuli oblasti paleopalia a archipallia, určené najmä na analýzu čuchových informácií a zachované v podobe hruškovitého laloku a hipokampu. V neopálovej kôre prebieha aj diferenciácia jednotlivých oblastí. Predná časť hemisfér obsahuje motorickú oblasť. nachádza sa tu čelný lalok okrem iného zvláda komunikáciu zvierat vrátane akustickej; u ľudí sa spája s rečou, teda druhým signálny systém. Zadná strana hemisfér je spojená s vnímaním vnemov. v tylový a temporálnych lalokov existujú oblasti, ktoré ovládajú zrak a sluch, resp. Ďalej vpred, v blízkosti motorickej oblasti, sú oblasti, ktoré vnímajú hmatové a proprioceptívne signály. U primátov centrálny sulcus (sulcus centralis), ktorý prechádza vrcholom hemisféry od mediálneho k laterálnemu povrchu, ohraničuje (aj keď nie celkom presne) motorickú oblasť od senzorickej. Pozdĺž predného okraja centrálneho sulku sú špecifické motorické oblasti umiestnené v lineárnom poradí a slúžia každej časti tela a končatín. Pozdĺž zadného okraja centrálneho sulku sú v rovnakom poradí umiestnené oblasti zmyslového vnímania zodpovedajúcich častí tela.
U mnohých cicavcov teda takmer celý povrch neokortexu zaberajú oblasti viac či menej úzko spojené s určitými senzorickými, resp. motorické funkcie. Hoci centrálny sulcus môže chýbať, vo väčšine prípadov majú placenty podobné lineárne usporiadanie senzorických a motorických oblastí proti sebe. U vačkovcov (a medzi placentami, u xenartrov) je „značenie“ oblastí tela približne rovnaké, ale zmyslové oblasti nie sú oddelené od motorických oblastí, ale sú nimi rozptýlené. Ale napríklad u ľudí tieto špecifické funkčné oblasti zaberajú relatívne málo miesta na povrchu neokortexu. Medzi nimi vznikli rozsiahle zóny šedej hmoty (jedna taká veľká plocha zaberá najviac frontálny lalok), ktoré nie sú spojené so špecifickými senzorickými alebo motorickými funkciami. Preto sa tieto oblasti často označujú ako „prázdne miesta“, hoci, ako ukazuje poškodenie týchto oblastí, práve v nich sa nachádzajú naše vyššie mentálne schopnosti vrátane príležitostí na učenie, iniciatívu, predvídavosť a úsudok. Sú však oblasti, ktoré možno odstrániť bez vážnych následkov na intelektuálnu činnosť. 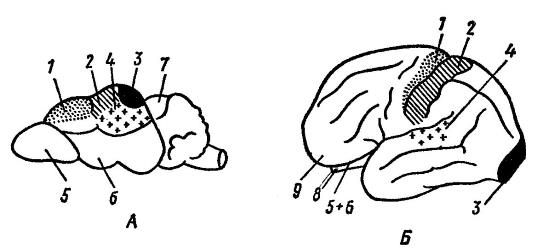
Funkčné centrá mozgovej kôry piskora ( sorex sp.) (A) a ľudský ( Homo sapiens
) (B) (podľa Naumova a Kartasheva, 1979):
1 - motorické centrum; 2 - centrum kožno-svalovej citlivosti; 3 - vizuálne centrum; 4 - sluchové centrum; 5 - čuchová žiarovka; 6 - čuchové laloky; 7 - strecha stredného mozgu; 8 - cerebellum; 9 - čelný lalok. Na evolúciu mozgu má veľký vplyv vonkajšie prostredie a motorická (potravinová, obranná) činnosť. Zároveň je vývoj rôznych častí mozgu determinovaný najmä spôsobmi hľadania potravy: u psa ( canis lupus), ktorý v tomto procese využíva čuch, je čuchový región rozvinutejší; u mačky ( Felis silvestris), hľadanie potravy pomocou zraku - zraku; pri makakoch ( Macaca mulatta), ktorý využíva zrak a sluch – zrakový a sluchový.
Zvyčajne sa predpokladá, že veľkosť mozgových hemisfér určuje rozdiely v mentálnych schopnostiach rôznych cicavcov. V istom zmysle je to pravda, ale s výraznými výhradami. Väčší mozog sa skladá z viac nervové bunky. Ak je oblasť existujúceho povrchu kôry akýmkoľvek spôsobom spojená s inteligenciou, potom je zrejmé, že z dvoch variantov mozgu rovnakej veľkosti bude ten s rozbrázdeným povrchom rozvinutejší a mozog s hladkým povrchom budú menej vyvinuté. Veľkosť samotného zvieraťa ovplyvňuje aj objem mozgu. To sa deje len preto, že mozog musí mať väčšie oblasti, aby slúžil väčším zmyslovým a motorickým spojeniam. Nárast veľkosti mozgu však nie je úplne úmerný telesnej hmotnosti, takže veľké zvieratá majú tendenciu mať relatívne menšie mozgy bez zjavnej straty inteligencie. Absolútna veľkosť mozgu teda nie je bezpodmienečným kritériom inteligencie. Určite tomu nasvedčuje fakt, že mozog veľryby môže byť objemovo päťkrát väčší ako ľudský mozog.
Porovnanie mozgu niektorých cicavcov:
1 - kôň; 2 - pes; 3 - klokan; 4 - osoba; 5 - slon. Percento mozgu v celkovej telesnej hmotnosti sa nazýva index cefalizácie. U veľkých hmyzožravcov je to asi 0,6%, u malých - do 1,2%, u veľkých veľrýb - asi 0,3% a u malých - do 1,7%. Väčšina primátov má cefalizačný index 1-2%. U ľudí dosahuje 2-3% a niektoré malé širokonosé opice majú mozog, ktorého hmotnosť je až 7% telesnej hmotnosti. Zároveň sa u moderných plazov a vtákov index cefalizácie pohybuje od 0,05 do 0,5%.
Nižšie je uvedená hmotnosť mozgu niektorých cicavcov (hmotnosť zvieraťa je uvedená v zátvorkách):
virginská vačica ( Didelphis virginiana) - 7,6 g (5 kg);
koala ( Phascolarctos cinereus) - 19,2 g (8 kg);
krovitý slon ( Loxodonta africana) - 6000 g (5000 kg);
obyčajný ježko ( Erinaceus europaeus) - 3,3 g (1 kg);
domáca myš ( Mus sval) - 0,3 g (0,02 kg);
šedá krysa ( Rattus norvegicus) - 2 g (0,3 kg);
veverička obyčajná ( Sciurus vulgaris) - 7 g (0,4 kg);
Európsky králik ( Oryctolagus cuniculus) - 11 g (3 kg);
domáci kôň ( Equus ferus) - 530 g (500 kg);
čierny nosorožec ( Diceros bicornis) - 500 g (1200 kg);
jeleň bielochvostý ( Odocoileus virginianus) - 500 g (200 kg);
žirafa ( Žirafa camelopardalis) - 680 g (800 kg);
domáce ovce ( Ovis orientalis) - 140 g (55 kg);
domáci býk ( Bos primigenius) - 490 g (700 kg);
ťava dvojhrbá ( camelus bactrianus) - 762 g (700 kg);
hroch ( Hroch obojživelník) - 580 g (3500 kg);
delfín bielohlavý ( Delphinus delphis) - 815 g (60 kg);
narval ( Monodon monoceros) - 2997 g (1578 kg);
vorvaň ( Physeter macrocephalus) - 8028 g (35833 kg);
modrá veľryba ( Balaenoptera musculus) - 3636 g (50900 kg);
domáca mačka ( Felis silvestris) - 25 g (3 kg);
Lev ( panthera leo) - 270 g (250 kg);
líška obyčajná ( Vulpes vulpes) - 53 g (4,5 kg);
domáci pes ( canis lupus) - 64 g (10 kg);
ľadový medveď ( Ursus maritimus) - 500 g (700 kg);
mrož ( Odobenus rosmarus) - 1130 g (700 kg);
kosmáč Geldi ( Callimico goeldii) - 7 g (0,2 kg);
kapucínka bieločelá ( Cebus albifrons) - 57 g (1 kg);
opica rhesus ( Macaca mulatta) - 88 g (6,5 kg);
pavián ( Papio cynocephalus) - 200 g (25 kg);
strieborný gibon ( Hylobates moloch) - 112 g (6,5 kg);
kalimantan orangutan ( Pongo pygmaeus) - 413 g (50 kg);
západná gorila ( gorila gorila) - 506 g (126 kg);
šimpanz obyčajný ( Pan troglodyty) - 430 g (55 kg);
rozumný človek ( Homo sapiens) - 1400 g (72 kg).
Z uvedených príkladov je zrejmé, že u menších cicavcov je mozog takmer vždy relatívne väčší a so zväčšovaním veľkosti tela zvieraťa sa relatívna veľkosť mozgu zmenšuje. Toto je obzvlášť výrazné u príbuzných druhov cicavcov – napríklad u mačky ( Felis silvestris) a lev ( panthera leo). Veľmi presvedčivé v tomto zmysle aj psy rôznych plemien. Ak telesné hmoty najmenší a najväčší veľké plemená sú približne v pomere 1:33, potom hmotnosti mozgu u tých istých plemien súvisia ako 1:3. 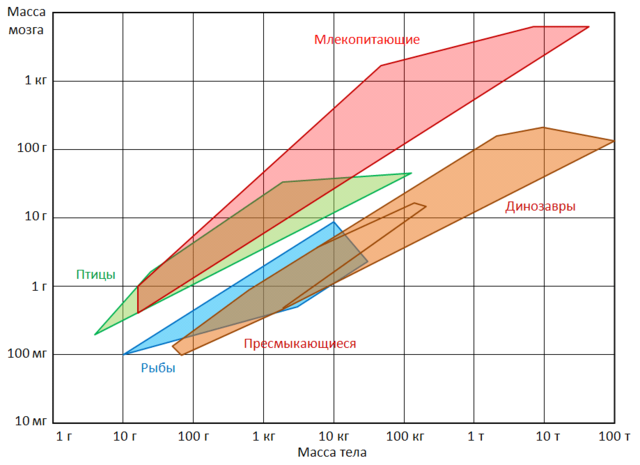
Rozsahy hmotnosti mozgu a tela pre niektoré skupiny stavovcov. U domestikovaných zvierat, zbavených potreby získať potravu a brániť sa pred nepriateľmi, je veľkosť mozgu výrazne znížená. Napríklad objem mozgu vlka ( canis lupus) je o 30 % väčší ako pes rovnakej veľkosti. Je zaujímavé, že tieto zmeny sa netýkajú len tradične domácich zvierat, ale aj zástupcov voľne žijúcich druhov, ktoré sú už nejaký čas chované v zajatí. Áno, líšky Vulpes vulpes), narodené v prírode, ale od prvých dní života v zajatí majú menší mozog ako ich príbuzní žijúci v prírodných podmienkach. Rozdiely zároveň dosahujú 20 %, čo približne zodpovedá rozdielu v objeme mozgu medzi divými a skutočnými domácimi zvieratami. Zmršťovanie mozgu, aj keď menej výrazné (približne 5 %), bolo zistené u vlkov chovaných v zajatí ( Canis), fretky ( Mustela), potkany ( Rattus). Zároveň pokles nezachytáva všetky časti mozgu, ale iba tie oblasti, ktoré sú spojené s prácou zmyslových orgánov. Najpozoruhodnejšia vec je, že u prepustených domácich zvierat sa zvyšuje hmotnosť mozgu. Napríklad divé mačky majú asi o 10% väčší mozog ako ich domáce náprotivky. Významný nárast v mozgu sa zistil aj u divých králikov ( Oryctolagus cuniculus) na Kerguelenských ostrovoch. divoké somáre ( Equus asinus) V Južná Amerika mal 15% veľký mozog než domáce. Je tiež zaujímavé, že mozog neandertálca ( Homo neanderthalensis) a paleolitický Homo sapiens ( Homo sapiens) boli o niečo väčšie ako mozog moderného človeka.
Zistilo sa, že u mnohých cicavcov sa pozoruje motorická asymetria, t.j. prevládajúce používanie pravej alebo ľavej polovice tela. Napríklad pri štúdiu nezlomených koní ( Equus ferus) zaznamenával, na ktorej nohe zvieratá začínajú chodiť, na ktorej strane radšej obchádzajú prekážky a na ktorej strane radšej ležia v sene. Výsledkom bolo, že väčšina kobýl bola pravákov a väčšina žrebcov bola ľavákov. Približne 10 % koní nedalo prednosť ani pravej, ani ľavej končatine. Podľa pozorovaní asi 90 % mrožov ( Odobenus rosmarus) vyhrabávať mäkkýše z morského bahna svojimi pravými plutvami. Uspávanie mláďat, asi 80 % samíc šimpanzov ( Panvica) a gorily ( Gorila) pritlačte hlavu na ľavú stranu hrudníka (približne rovnaké percento je pozorované u žien). Potkany ( Rattus), vedúci hľadania potravy pomocou vibrissae umiestnených na pravá strana náhubky, sú väčšou korisťou ako ich ľaváci príbuzní.

