Pôvod mozgu Saveliev Sergey Vyacheslavovič
§ 37. Asociačné centrum mozgu plazov
Vzhľadom na všeobecný plán budovy nervový systém, mali by sme sa osobitne zaoberať novými princípmi organizácie a fungovania mozgu, prvýkrát implementovanými u plazov. Nervový systém archaických amniotov sa stal logický vývoj budovy úspešného obojživelného dizajnu. Mozog obojživelníka však prakticky vykonával funkciu zložitého reflexného aparátu a jeho intelektuálne schopnosti zostali nevyužité. O evolúcii obojživelníkov rozhodovali svaly, zuby, lineárne rozmery a mierky rozmnožovania.
Zo všetkých cicavcov je človek najrozvinutejší a vyvinutý mozog. U všetkých stavovcov sa však mozog skladá z 3 hlavných štruktúrnych jednotiek: mozgové hemisféry, mozgový kmeň a mozoček. Tvorba siete je hlavnou oblasťou výmeny informácií, ktorá je zdrojom mozgovej energie. V tejto sieti nervové vlákna každý neurón môže byť prepojený s inými zosieťovanými formátmi, ktoré neustále prijímajú správy zo zmyslových orgánov, ktoré zase vysielajú elektrickú aktivitu cez mozgovú kôru.
Udržiava vedomie. Ak zošitý pás zníži ich aktivitu, človek môže zaspať alebo trpieť halucináciami, vidieť, počuť a mať nereálne pocity. Výsledkom zmyslovej deprivácie, keď je človek niekoľko hodín zavretý v tmavej, tichej a prázdnej miestnosti, je zdanlivá nálada, tranz alebo kóma.
Došlo k elementárnemu rozvoju potravinových zdrojov, kde nebol priestor ani biologická nutnosť pre rozvoj komplexného správania. So stopami tohto obdobia evolúcie stavovcov čelíme pokusu o rozvoj podmienené reflexy u rôznych predstaviteľov moderných obojživelníkov. Extrémne nízka schopnosť učenia a nedostatok dlhodobej pamäte na akumuláciu individuálnych skúseností ukazujú, že starí obojživelníci vo svojom vývoji nikdy nečelili zložitým behaviorálnym úlohám.
Rysy vývoja zmyslových orgánov a znaky komplexného správania plazov sú založené na znakoch štrukturálnej organizácie mozgu. Mozog plazov sa líši od mozgu obojživelníkov v množstve aj množstve kvalitatívne. Pred príchodom amniot sa stratégie správania alebo reakcie na konkrétny stimul zvolili podľa princípu dominancie (pozri obr. III-6, f). Tento princíp spočíva v tom, že mnohé primárne vodné stavovce alebo obojživelníky nemajú výrazné veľké asociačné centrum mozgu (pozri obr. III-6, e). K voľbe formy správania dochádza na základe porovnania aktivít približne ekvivalentných častí mozgu obsluhujúcich rôzne zmyslové orgány. Rozhodujúcu úlohu zohráva úroveň excitácie mozgových analytických centier jedného z analyzátorov. Hlavnou oblasťou pre rozhodovanie sa stáva reprezentácia zmyslového orgánu, ktorý dosiahol v mozgu najväčšiu excitáciu. Po výbere jednej z inštinktívnych reakcií dochádza k jej behaviorálnej implementácii. Tento proces je riadený tým istým jednoduché porovnanie dominancie. Ak v priebehu reakcie vznikne nové podráždenie, ktoré zmení pomer vzruchov zmyslových orgánov, potom sa behaviorálna realizácia inštinktívneho procesu zastaví. Každá konkrétna situácia je iná ako predchádzajúca, no zapája sa do nej rovnaký súbor zmyslov. Ak sa najväčšia excitácia dosiahne v tom istom zmyslovom systéme, potom sa správanie zachová, a ak v inom, potom sa zmení. Keďže absolútne identické podmienky sa v prirodzenom živote prakticky nikdy nevyskytujú, správanie aj tých najprimitívnejších anamnií bude nekonečne rôznorodé. V dôsledku toho bude správanie každého jednotlivca individuálne s pomerne vysokou dynamickou prispôsobivosťou.
Prvé známky asociatívneho centra sa objavili v mozgu obojživelníkov. V nich by sa takýmto centrom mohol stať stred alebo diencephalon. Boli na to všetky dôvody. V diencefale sú neuroendokrinné centrá, ktoré riadia sexuálne správanie, migráciu a energetickú bilanciu anamniotického organizmu. Prostredníctvom aktivácie centier diencefala sa spúšťajú inštinktívne programy správania, ktoré riadia prácu iných častí mozgu. Zdalo by sa, že diencephalon by sa mohol stať analytickým centrom správania anamnie a potom amniotov. V tomto prípade by však systém implementácie behaviorálnych reakcií spočíval nielen v práci nervového systému. Zakaždým by každá udalosť správania viedla k stimulácii práce neuroendokrinných centier. Hormonálna regulácia správania sa vykonáva po dlhú dobu a neurologická - rýchlo. Pri rýchlej zmene foriem správania by nastal konflikt medzi inertnými hormonálnymi a dynamickými nervovými programami správania. U hmyzu bol tento konflikt vyriešený v prospech neurohormonálnych centier a čisto inštinktívneho správania.
Pomerne nestabilná situácia sa vyvinula v prípade primárnych vodných stavovcov a obojživelníkov. Na jednej strane je úloha hormonálno-inštinktívnej regulácie správania u obojživelníkov veľmi dôležitá a jednoznačne dominuje pri výbere stratégií správania. Na druhej strane neuromorfologický substrát je dostatočne vyvinutý na miernu individualizáciu správania pri realizácii týchto stratégií. Vznikol originálny systém hormonálne dominantného výberu foriem správania zo štandardnej inštinktívnej množiny. U obojživelníkov je stratégia správania určená neurohormonálnym stavom jednotlivca. Pri realizácii zvolenej formy správania sa prispôsobuje konkrétnym podmienkam pomocou porovnávania dominancie, ktoré bolo popísané vyššie. V takejto schéme kontroly správania anamnios nie je miesto pre asociatívne centrum. Mohol by byť potrebný len vtedy, keď by bola potrebná rýchla adaptačná individualizácia správania. Takáto situácia môže nastať len vtedy, keď dôsledná realizácia inštinktívnych foriem správania bude priamo závisieť od neustále sa meniacej situácie.
V takýchto nestabilných podmienkach životné prostredie sa ukázali ako archaické plazy. Zjavne sa zvýšili požiadavky na rýchlu individualizáciu správania a pamäte a implementácia princípu dominantného hormónu výberu zo štandardného inštinktívneho súboru správania sa stala neúčinnou. Vznikol úplne nový typ rozhodovania, ktorý sa zachoval v mozgoch moderných plazov patriacich do vzdialených systematických skupín. Všetky spája jedna zásadne nová kvalita mozgu – výrazné asociatívne centrum (pozri obr. III-7).
Hlavné asociačné centrum plazov sa vytvorilo v streche stredného mozgu (pozri obr. III-5, c; III-6; III-7, b). Vznikol na základe viacerých zmyslových orgánov, ktoré mali zastúpenie v tejto časti mozgu. Hlavná časť strechy stredného mozgu je obsadená reprezentáciou vizuálny systém. zrakové nervy po prechode cez chiazmatu sa krížia a stúpajú na strechu stredného mozgu. Axóny gangliových buniek sietnice končia na neurónoch strechy stredného mozgu, ktoré sú organizované do stratifikovaných štruktúr (pozri obr. III-5, c; III-7, b). Existuje jasné topografické spojenie medzi určitou oblasťou sietnice a strechou stredného mozgu. Zároveň tvar obrazu a vzájomného usporiadania jeho prvky. Po dlhú dobu bola predná časť strechy stredného mozgu považovaná výlučne za mozgové centrum. vizuálny analyzátor. Funkčno-morfologické štúdie však ukázali, že to tak ani zďaleka nie je.
Spolu s reprezentáciou zrakového systému prichádzajú na strechu stredného mozgu informácie o somatickej (kožnej) citlivosti, motorickom analyzátore, vestibulárnych a sluchových signáloch (pozri obr. III-6, e). sluchový analyzátor u plazov výrazne zvyšuje svoje zastúpenie v tomto centre. Výsledkom je, že u mnohých plazov v zadnej časti strechy stredného mozgu sa objavujú nenápadné párové výbežky - zadné alebo dolné tuberkulózy. Strecha stredného mozgu sa stáva nie homogénnou anatomickou formáciou, ako pri anamnii, ale kvadrigeminou. Sústreďuje zastúpenie takmer všetkých hlavných vzdialených a kontaktných analyzátorov. Aj čuchové ústrojenstvo má svoje zastúpenie v streche stredného mozgu. S výnimkou čuchového ústrojenstva sú takmer všetky zmyslové projekcie do strechy plazieho stredného mozgu topologického charakteru. To znamená, že informácie z každej konkrétnej oblasti tela sú prezentované striktne určitej oblasti strecha stredného mozgu. Zachováva sa princíp mapy tela, ktorá sa bodovo prenáša do mozgu.
V streche stredného mozgu plazov sa teda sústreďujú najrôznejšie informácie o stave ich vlastného organizmu a okolitého sveta, ktoré sa kombinujú podľa topologického princípu. Zvážte, čo sa stane v streche stredného mozgu, ak predná pravá končatina plaza jednoducho stojí na nezvyčajnom povrchu. Pri hodnotení takejto situácie v streche stredného mozgu, komparatívna analýza somatické, senzomotorické, sluchové a zrakové informácie. Je to jednoduché, pretože všetky signály sú sústredené v jednom strede a často na sebe, ako vo vrstvenom koláči. Stredný mozog vedie komplexná analýza veľa faktorov jedného javu, čo umožňuje zvoliť najvhodnejšiu reakciu. Na to je ideálna vrstvená štruktúra strechy stredného mozgu.
Extrémne zjednodušenie reálnej situácie, môžeme povedať, že v streche stredného mozgu, zastúpenie rôznych zmyslové systémy umiestnené na podmienených "podlahách" usporiadaných v horizontálnej rovine. Každé poschodie zaberá akási mapa. Môže ísť o momentálnu informačnú mapu receptorových signálov z povrchu tela, obraz na sietnici alebo akustické pole. Všetky tieto mapy sú na svojich „podlahách“ orientované tak, že odrážajú približne rovnaký smer v priestore. Sluchový signál z prednej pravej končatiny leží pod jej vizuálnou mapou a nad somatickým signálom z kože nohy. Špecializované "poschodia" sú navzájom korelované pomocou vertikálnych odkazov, ktoré umožňujú rýchle posúdenie konkrétnu situáciu a urobiť správne rozhodnutie. Táto schéma strechy stredného mozgu nám umožňuje pochopiť reflexnú rýchlosť mozgu plazov. Zrejme práve táto rýchlosť sa stala hlavným dôvodom evolučného úspechu archaických plazov.
Vznik dokonalého centra reflexného rozhodovania u plazov mal niekoľko dôležitých dôsledkov. Na jednej strane je možnosť rýchleho výberu riešení nezmyselná, ak celková rýchlosť metabolizmu zostane na rovnakej úrovni. V dôsledku toho bol vývoj stredného mozgu sprevádzaný zvýšením metabolizmu. Na druhej strane, zväčšenie veľkosti strechy stredného mozgu vytvorilo potrebný bunkový substrát pre rozvoj pamäte. Individuálna skúsenosť zvieraťa sa stala základom pre porovnávanie udalostí oddelených v čase. Je ťažké preceňovať túto udalosť. Prvýkrát vznikli skutočné základy pre individualizáciu správania na základe porovnávania rôznych udalostí. Treba poznamenať, že tieto neurobiologické výhody plazieho mozgu sa zvyčajne ani nezohľadňujú pri rekonštrukcii ranej evolúcie amniotov (Carroll, 1982).
Archaické plazy v nadväznosti na rozvoj centra reflexnej analýzy dostali materiálny substrát na zapamätanie si rôznych udalostí. Sprístupnila sa im reprodukcia individuálnej skúsenosti, ktorá slúži ako základ pre učenie. Obojživelníky už nemohli konkurovať archaickým plazom, ktorí sa sami učia. Hormonálno-inštinktívne princípy správania obojživelníkov, rýb a bezstavovcov z nich urobili potravu pre plazy s vyvinutým reflexne asociatívnym stredným mozgom.
Všetky tieto výhody štruktúry mozgu plazov nemohli vzniknúť samy osebe. Pre takú hlbokú kvalitatívnu reštrukturalizáciu mozgu museli vzniknúť mimoriadne drsné a mimoriadne podmienky. Archaické plazy sa museli ocitnúť v zvláštnom prostredí s veľmi vysokými nárokmi na analytické vlastnosti mozgu a individuálnu pamäť.
Z knihy Nálezy zvyškov vrchnojurských plazov v bridlicovej bani Saveljevskij autora Z knihy Nálezy zvyškov vrchnojurských plazov v bridlicovej bani Saveljevskij autora Žuravlev Konstantin IvanovičNálezy pozostatkov plazov V roku 1931, hneď v prvých mesiacoch ťažby bridlíc, keď sa ešte ťažilo povrchovými jamami, sa podarilo získať niekoľko nálezov pozostatkov ichtyosaura. Medzi týmito nálezmi je séria 13 chvostových stavcov nájdených vo vrstve ropnej bridlice na základni
autora§ 36. Celkový plán stavby nervovej sústavy plazov S rozsiahlym rozvojom krajiny sa mozog plazov zmenil. Jeho morfologické preskupenia však na prvý pohľad neboli také zásadné ako u obojživelníkov. Povrchová anatomická analýza ukazuje, že starovek
Z knihy Pôvod mozgu autora Saveliev Sergej Vjačeslavovič§ 38. Podmienky pre vznik mozgu plazov Asociačné mozgové centrum nemôže vzniknúť náhodou. Energetické náklady na udržiavanie asociatívneho centra mozgu a náklady na zmenu stratégií správania sú vždy veľmi vysoké (pozri kapitolu I). Na to musí byť dôvod
Z knihy Pôvod mozgu autora Saveliev Sergej Vjačeslavovič§ 40. Adaptívne žiarenie archaických plazov Malé dravé a všežravé plazy, ktoré sa vyvinuli v agresívnom prostredí rastlinných zvyškov, neboli najväčšími živočíchmi tej doby. Ich telesná veľkosť na konci karbónu zvyčajne nepresahovala niekoľko desiatok
Z knihy Biologická chémia autora Lelevič Vladimír ValeryanovičAktívne miesto enzýmu Oblasť molekuly enzýmu, ktorá špecificky interaguje so substrátom, sa nazýva aktívne miesto. Aktívne miesto je jedinečná kombinácia aminokyselinových zvyškov v molekule enzýmu, ktorá poskytuje priame
autoraKde je ťažisko systému Zem-Mesiac? Ťažisko sústavy Zem-Mesiac, takzvané barycentrum, sa nachádza vo vzdialenosti 4672 kilometrov od stredu Zeme smerom k Mesiacu, teda v hĺbke približne 1700 kilometrov pod povrchom Zeme. Presne povedané, podľa
Z knihy najnovšia kniha faktov. Zväzok 1. Astronómia a astrofyzika. Geografia a iné vedy o Zemi. Biológia a medicína autora Kondrashov Anatolij PavlovičČo je to centrum potešenia a kde sa nachádza v tele? Jednou z častí mozgu je hypotalamus, ktorý je súčasťou diencefala a nachádza sa pod zrakovými tuberkulami (talamus). Hypotalamus, v ktorom sú umiestnené autonómne centrá
autora Kondrashov Anatolij Pavlovič Z knihy Najnovšia kniha faktov. Zväzok 1 [Astronómia a astrofyzika. Geografia a iné vedy o Zemi. Biológia a medicína] autora Kondrashov Anatolij Pavlovič Z knihy Farmaceutická a potravinová mafia od Browera LouisaCentrum pre farmakologický dohľad „Paríž, 4. februára 1977. NCPC, založené v januári 1974, sa stretlo na Valnom zhromaždení 2. februára spolu s delegátmi Národnej charty lekárov a farmaceutov, ku ktorým sa pripojila skupina centier pre kontrolu z intoxikácie a
Z knihy V divočine času autora Čiževskij Nemec MichajlovičKlasifikácia a stručný popis hlavných skupín obojživelníkov, plazov, vtákov a cicavcov TRIEDA Obojživelníky alebo obojživelníky Podtrieda Apsidospondylus Nadrad Labyrintodonty Skupina obojživelníkov, ktorá vyhynula najneskôr v období triasu a vrchol dosiahla na konci paleozoika.
autora Kandel Eric Richard Z knihy Hľadanie pamäti [Vznik novej vedy o ľudskej psychike] autora Kandel Eric Richard Z knihy Psychopati. Spoľahlivý príbeh o ľuďoch bez ľútosti, bez svedomia, bez výčitiek svedomia Autor: Keel Kent A.Mendota Juvenile Rehabilitation Center Začiatkom 90. rokov zachvátila Spojené štáty skutočná epidémia tínedžerského násilia. Počet trestných činov spáchaných mladistvými sa medzi rokmi 1980 a 1993 takmer zdvojnásobil. Zdalo sa, že to nemôže nič zastaviť
Z knihy Čítanie medzi riadkami DNA [Druhý kód nášho života alebo Kniha, ktorú si každý musí prečítať] autor Shpork PeterZ Berlína do centra budúcej revolúcie Neurazí sa, keď si ho pomýlia so študentom alebo doktorandom. A to sa 32-ročnému genetikovi stáva neustále. Alexander Meissner - sivomodré oči, tmavé blond nedbalo vyčesané vlasy, trojdňové strnisko - nielen mladé,
Názov materiálu: Základy zoológie a zoogeografie
všeobecné charakteristiky
Vzhľadom na vzhľad krku u plazov je hlava pohyblivo spojená s telom a môže sa pohybovať nielen, ako u obojživelníkov, vo vertikálnom smere, ale aj v horizontálnom smere.
Telo je predĺžené (iba u korytnačiek je široké), skôr pružné; chvost veľkej väčšiny druhov je dlhý, schopný rôznych pohybov; nohy sú silne vyvinuté (u niektorých druhov jašteríc a u všetkých hadov sú nohy znížené v dôsledku prechodu na iný aktívny spôsob pohybu). Priemerná veľkosť tela je oveľa väčšia ako u obojživelníkov a niektoré druhy dosahujú obrovské veľkosti.
Kryty. Koža je pokrytá zrohovatenými pevnými a zároveň ľahkými šupinami ektodermálneho pôvodu, ktoré chránia telo nielen pred mechanickému poškodeniu ale aj z vysychania. Keratinizovaná vrstva kože sa počas línania pravidelne odlupuje. U korytnačiek a krokodílov sa osifikácia vyvíja v mezenchymálnej vrstve kože, zatiaľ čo u ostatných plazov chýbajú. Korytnačky sú dobre chránené pred nepriateľmi mušľami. Koža moderných plazov, na rozdiel od obojživelníkov, je suchá, bez žliaz.
Farba vonkajších krytov je rôznorodá a u mnohých druhov má ochrannú hodnotu, pretože vďaka nej sú zvieratá na pozadí prostredia sotva viditeľné.
Nervový systém. Vďaka mobilnejšiemu a komplexnejšiemu životnému štýlu je nervová sústava plazov lepšie vyvinutá ako u obojživelníkov (obr. 95).
Ryža. 95. Mozog jašterice, bočný pohľad:
1 - hemisféra predný mozog; 2 - čuchový lalok; 3 - diencephalon; 4 - hypofýza; 5 - stredný mozog; 6 - optický chiasm (chiasm); 7 - cerebellum; 8 - miecha
Predný mozog je výrazne veľké veľkosti než posledne menované (najmä v dôsledku nárastu striata), objavujú sa na povrchu hemisfér nervové bunky- rudiment mozgovej kôry. Preto plazy rýchlo nadväzujú nové spojenia s vonkajším prostredím ako ich predkovia. Úloha tejto časti mozgu ako koordinátora vitálnej činnosti celého organizmu sa zvyšuje. Štruktúra diencefala a stredného mozgu sa stáva zložitejšou, zvyšuje sa ich prepojenie s predným mozgom a ostatnými časťami centrálneho nervového systému. Cerebellum je v dôsledku komplikovanosti spôsobov pohybu a potreby udržania rovnováhy tela oveľa lepšie vyvinutý ako u obojživelníkov. Zväčšila sa aj medulla oblongata, ktorá tvorí vertikálny ohyb charakteristický pre plazy. Miecha je dlhá; jeho štruktúra sa stala zložitejšou. To sa prejavilo v ostrejšom oddelení vonkajšej vrstvy (biela, t.j. dráhy) od vnútornej (sivá, t.j. nervové bunky) vrstvy.
Zo zmyslových orgánov u plazov (s výnimkou hadov a niektorých iných druhov) sú oči lepšie vyvinuté ako u obojživelníkov: sú pohyblivejšie. Ich prispôsobenie sa videniu na rôzne vzdialenosti (u mnohých druhov dosť veľké) je zabezpečené nielen ako u rýb a obojživelníkov pohybom šošovky, ale ako u vyšších stavovcov aj zmenou jej zakrivenia. Okrem očných viečok sa tu nachádza mliečna membrána. Mnohé druhy majú parietálny orgán spojený s diencefalom a schopný vnímať svetelné podnety. V orgáne sluchu sa vyskytli niektoré progresívne zmeny: dolný vak sa mierne zvýšil; sluchová kostička lepšie prenášať vibrácie ušný bubienok do vnútorného ucha; okrem oválneho okna sa objavilo druhé okno - okrúhle, ktoré zvyšuje pohyblivosť exolymfy obklopujúcej labyrint; Bubienok sa nenachádza na povrchu hlavy, ale v malej priehlbine. Orgány čuchu sú vyvinuté celkom dobre, ich vnútorný povrch sa zväčšil. Chuťové orgány u väčšiny druhov sú slabo vyvinuté. Jazyk zohráva v kontakte dôležitú úlohu.
Kostra. Je takmer úplne osifikovaná v dôsledku nahradenia hrubých vláknitých kostného tkaniva, charakteristický pre obojživelníky, tenký vláknitý, zvýšila sa jeho pevnosť, zlepšili sa nosné a ochranné funkcie (obr. 96).
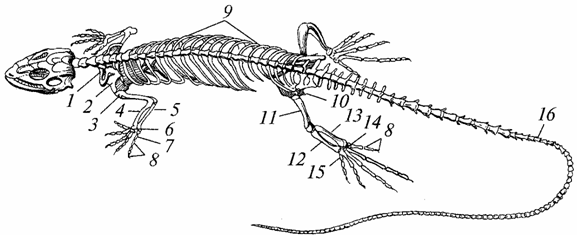
Ryža. 96. Kostra jašterice:
7 - kľúčna kosť; 2 - lopatka; 3 - rameno; 4 - polomer; 5 - ulna; b - zápästie; 7 - metakarpus; 8 - falangy prstov; 9 - rebrá; 10 - panva; 11 - stehno; 12 - veľký holennej kosti; 13 - fibula; 14 - tarzus; 15 - metatarsus; 16 - chvostové stavce
Lebka zväčšená; kapacita sekcie mozgu bola väčšia ako kapacita obojživelníkov. Bočný povrch lebka starých plazov bola pevná, neskôr sa v nej objavili časové jamy - jedna alebo dve, čo znížilo jej hmotnosť a zväčšilo povrch na pripevnenie žuvacie svaly. Dolné čeľuste, podobne ako u obojživelníkov, sú zavesené na mozgovej oblasti lebky pomocou štvorcovej kosti a tiež pozostávajú z niekoľkých kostí (zubných, kĺbových atď.).
Telo prvého stavca (atlas) dorástlo v podobe zuba k telu druhého stavca (epistrofia). Prstencový atlas dostal možnosť otáčať sa okolo epistrofického zuba, vďaka čomu bolo možné otáčať hlavou, čo malo za následok veľký význam na získavanie potravy (hlavne iných zvierat) a na ochranu pred nepriateľmi. Atlas je spojený s okcipitálnou oblasťou lebky pomocou jedného kondylu, čo tiež prispieva k zvýšeniu pohyblivosti hlavy. Chrbtica je rozdelená do piatich častí: krčná, hrudná, drieková, sakrálna a chvostová. Stavce hrudný sú spojené s hrudnou kosťou dobre vyvinutými rebrami, v dôsledku čoho sa vytvára hrudník, ktorý u obojživelníkov chýba. Skrátené rebrá vybiehajú z bedrových stavcov a nedosahujú hrudnú kosť. Existujú dva sakrálne stavce (obojživelníky majú jeden). V kaudálnej oblasti je veľa stavcov (s výnimkou korytnačiek).
Kosti nôh sú lepšie vyvinuté a odolnejšie ako kosti obojživelníkov. Medzi kosťami zápästia a tarzu sú pohyblivé kĺby. Preto predlaktie a proximálny rad zápästia, ako aj predkolenie a proximálny rad tarzu fungujú ako integrálne formácie. Za zmienku stojí najmä spevnenie pásov končatín: ramena - vzhľadom na spojenie s hrudnou kosťou a hrudnou chrbticou, panvy - vďaka tomu, že sa spolieha na dve sakrálne stavce a obe jeho polovice tvoria dve zrasty – ischiálnu a pubickú.
Svalový systém. Prešla diferenciáciou, vyvinuli sa nové svalové skupiny, na ktorých sa otáča hlava, pohyby krku, ktoré vznikli u plazov, rozšírenie a zúženie hrudník a iné.Výrazne sa zväčšili svaly nôh, vyvinuli sa flexory a extenzory prstov. Hmotnosť a pevnosť celého systému sa zvýšila.
Zažívacie ústrojenstvo. Je diferencovanejší ako u obojživelníkov (obr. 97). Zuby sú sústredené najmä na čeľustiach, ich počet je veľký, no sú rovnakého typu a slúžia najmä na ulovenie koristi. Slinné žľazy sú lepšie vyvinuté ako u obojživelníkov (u mnohých druhov sú sekréty týchto žliaz jedovaté). U plazov začína proces tvorby tvrdého podnebia, ktoré je vyvinuté najmä u krokodílov. Vďaka vytvoreniu tvrdého podnebia je ústna dutina oddelená od nosohltanovej dutiny, čo uľahčuje dýchanie aj vstrebávanie potravy. Hltan, pažerák a žalúdok sú dobre vyvinuté (najmä u predátorov, ktorí jedia stavovce). Pečeň (so žlčníkom) a pankreas sú dokonalejšie ako u obojživelníkov. Tenké črevo výrazne predĺžené, hrubé črevo je krátke a končí sa kloakou. Moderné plazy sa živia hlavne zvieratami, počet bylinožravých druhov je zanedbateľný.
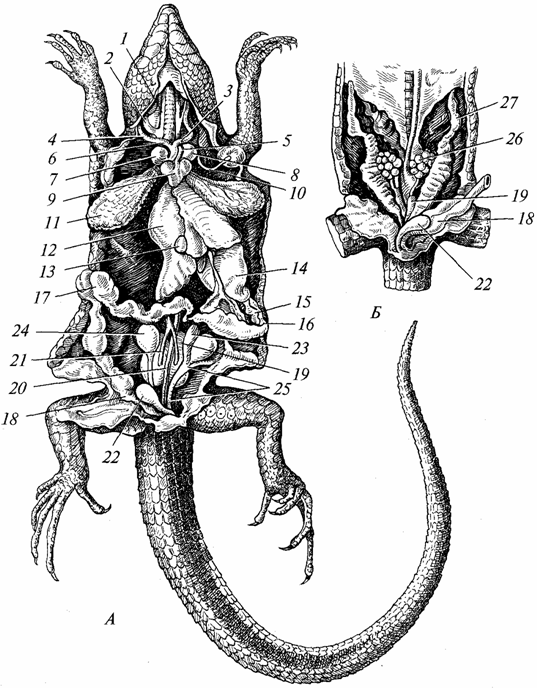
Ryža. 97. Vnútorné orgány jašterice; A - mužský; B - žena:
/ - vonkajšie a 2 - vnútorné krčné žily; 3 - ľavé a 4 - pravé krčné tepny; 5 - ľavý a 6 - pravý aortálny oblúk; 7 - pravá a 8 - ľavá predsieň; 9 - srdcová komora; 10 - ľavá podkľúčová tepna;11 - pľúca; 12 - pečeň; 13 - žlčníka; 14 - žalúdok; /5 - pankreas; 16 - dvanástnik; 17 - hrubé črevo;18 - konečník; 19 - oblička; 20 - dorzálna aorta; 21 - eferentné žily obličiek; 22 - močový mechúr; 23 - príloha semenníka; 24 - semenník; 25 - trubica na osivo; 26 - vaječník; 27 - vajcovod
Dýchací systém. Dýchacími orgánmi plazov sú pľúca (pozri obr. 97), pretože ich koža je nevhodná na výmenu plynov; sliznica ústna dutina, ktorý zohráva významnú úlohu pri dýchaní obojživelníkov, na túto funkciu využíva niekoľko druhov plazov. Z tohto dôvodu a najmä v súvislosti so zvýšenou aktivitou plazov v porovnaní s obojživelníkmi sa stavba pľúc resp. dýchacieho traktu komplikované. Pľúca sú väčšie orgány a sú umiestnené hlbšie v hrudnej dutine ako u obojživelníkov. Ich vnútorný povrch sa výrazne zväčšil v dôsledku vývoja mnohých priečnikov s hustejšou sieťou. cievy. Vzduch vstupuje do pľúc, prechádza cez choanae, hltan, hrtan, dlhú priedušnicu, ktorej lúmen je podopretý chrupavkovitými krúžkami. Pri prechode priedušnicou sa vzduch zbavuje prachu, jeho teplota sa blíži telesnej teplote. U obojživelníkov bola priedušnica v plienkach. Zadný koniec priedušnice je rozdelený na dva priedušky, v stenách ktorých sú tiež chrupavkové krúžky. Realizácia nádychu a výdychu je účinnejšia ako u obojživelníkov vďaka vývoju hrudníka u plazov, ktorého objem sa s nádychom zväčšuje a s výdychom zmenšuje. Rovnaký mechanizmus sa zachoval aj u vyšších stavovcov – vtákov a cicavcov.
Obehový systém. Srdce, podobne ako srdce obojživelníkov, je trojkomorové, ale zospodu v komore sa objavuje zárodok septa. U krokodílov je už plná a ich srdce sa stáva štvorkomorovým (dve predsiene a dve komory). Následne je u plazov zosilnená separácia venóznych a arteriálnych krvných prúdov, ale úplná izolácia oboch prúdov ešte nie je úplná ani u krokodílov, ako je zrejmé z nasledujúceho popisu (obr. 98).
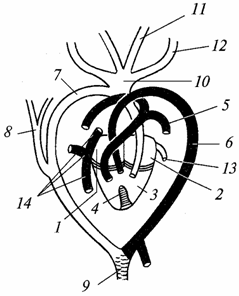
Ryža. 98. Srdce a hlavné cievy jašterice (svetlé - cievy s arteriálnou krvou, tieňovanie - so zmiešanou krvou; čierne - s venóznym):
1 - pravé átrium; 2 - ľavej predsiene; 3 - komora; 4 - neúplná priehradka rozdeľujúca komoru na ľavú a pravú polovicu; 5 - pľúcna tepna; 6 - ľavý oblúk aorty; 7 - oblúk pravej aorty; 8 - podkľúčová tepna; 9 - dorzálna aorta; 10 - spoločný kmeň krčné tepny; 11 - ľavá vnútorná krčnej tepny, 12 - ľavá vonkajšia krčná tepna; 13 - pľúcna žila; 14 - späť vena cava a predné vena cava (jugulárne) žily
Arteriálna krv vstupuje do ľavej predsiene - pochádza z pľúc. Venózna krv vstupuje do pravej predsiene – odoberá sa zo všetkých častí tela. Ďalej krv z oboch predsiení vstupuje do komory: v pravej polovici - venóznej, v ľavej - arteriálnej. Kvôli prítomnosti prepážky, hoci neúplnej, sa krv v komore mieša menej ako u obojživelníkov. Neexistuje žiadny arteriálny kužeľ a hlavné arteriálne kmene odchádzajú priamo zo srdca.
Od pravá strana komory odchádzajú pľúcne tepny(bývalý štvrtý pár branchiálnych artérií), ktorý vedie venóznu krv do pľúc. Bývalý tretí pár branchiálnych ciev atrofoval. Druhý pár ciev pozostáva z dvoch aortálnych oblúkov: pravý oblúk začína z ľavej strany komory a vedie arteriálnu krv, ľavý oblúk začína z pravej strany komory (ale bližšie k jej ľavej strane) a nesie zmiešanú krv . Z pravého oblúka aorty vybiehajú krčné tepny (bývalý prvý pár žiabrových ciev) k hlave. Zásobujú mozog, obzvlášť citlivý na nedostatok kyslíka, arteriálnou krvou. Oba aortálne oblúky sa spájajú a vytvárajú aortu, ktorá sa rozvetvuje do mnohých tepien. Telo plazov je teda zásobované hlavne zmiešanou krvou, ale je nasýtené viac kyslíkom ako obojživelníky, čo prispieva k intenzívnejšiemu metabolizmu.
vylučovací systém. Veľa sa zmenilo v dôsledku zvýšenej rýchlosti metabolizmu plazov, čo viedlo k zvýšeniu množstva produktov disimilácie a ich úplnému prispôsobeniu sa suchozemskému životnému štýlu, v dôsledku čoho bolo potrebné hospodárnejšie využívať vstupnú vodu. telo. Preto sa u plazov vyvinuli zložitejšie vylučovacie orgány nazývané panvové alebo sekundárne obličky (metayaephros). Všetky tubuly týchto obličiek nemajú lieviky a začínajú Bowmanovými kapsulami obklopujúcimi Malpighove telá so zvýšeným počtom kapilár. Tubuly panvových obličiek sú veľmi dlhé a husto prepletené kapilárami. V Bowmanových kapsulách z produktov disimilácie rozpustených vo vode a niekt organickej hmoty(napr. glukóza) sa tvorí primárny moč. Ten pri prechode cez dlhé obličkové tubuly vracia krvi cez kapiláry opletené ich stenami významnú časť vody a ďalších látok potrebných pre telo. Sekundárny moč, ktorý sa potom vytvorí, neodchádza cez vlčie kanály, ale cez novo vyvinuté močovody do kloaky a hromadí sa v močového mechúra. Procesy tvorby primárneho a sekundárneho moču sa tiež uskutočňujú v obličkách trupu, ale s návratom menšieho množstva vody do krvi. Primárne vodné stavovce, najmä obojživelníky, ktorých koža je holá, často trpia nadbytkom vody, a nie jej nedostatkom. Treba tiež poznamenať, že u plazov je konečným produktom degradácie bielkovín kyselina močová, na odstránenie ktorých z tela močom sa minie málo vody. U obojživelníkov a mnohých ďalších primárnych vodných stavovcov vedie rozklad bielkovín k tvorbe močoviny, ktorej odstránenie vyžaduje veľa vody.
Embryá plazov majú základ obličky hlavy a vyvinutú obličku trupu, ktorá funguje u dospelých (u mnohých druhov jašteríc) až do puberty. Tieto skutočnosti potvrdzujú kontinuitu troch typov obličiek počas evolúcie stavovcov.
Rozmnožovanie. Mužský reprodukčný systém plazov zahŕňa dva semenníky okrúhleho tvaru ležiace po stranách driekovej chrbtice, dva prívesky semenníkov vytvorené z predných častí obličiek trupu, dva semenné kanáliky (Wolffove kanály) a kopulačné orgány vyvinuté zo steny. z kloaky. Dámske reprodukčný systém pozostáva z dvoch vaječníkov, ktoré silne rastú počas obdobia rozmnožovania, a dvoch vajcovodov (Müllerových kanálov), ktoré ústia do kloaky. Hnojenie je len vnútorné. Semeno vnášajú samce do kloaky samice pomocou kopulačných orgánov. Zrelé vajcia, ktoré obsahujú veľa žĺtka, spadajú do lievikov vajcovodov. K spojeniu ženských a mužských gamét dochádza v hornej časti vajcovodov, kam po pohlavnom styku prenikajú spermie. Zygoty, ktoré sa pohybujú pozdĺž vajcovodov, sú v strednej časti obklopené proteínovou škrupinou a na zadnej strane sú pokryté škrupinou.
U niektorých druhov jašteríc a hadov sa pozoruje partenogenetické rozmnožovanie samíc. V týchto druhoch zvyčajne nie sú žiadni muži.
rozvoj. U plazov je vývoj priamy; plazy nemajú larvy. Oplodnené vajíčka zvyčajne kladú plazy do pôdy a niektoré druhy - pod hnijúce rastlinné zvyšky. U mnohých druhov prebieha vývoj zygotov v pohlavnom trakte samice a hneď po uvoľnení vyvinutých vajíčok sa z nich vyliahnu formované živočíchy, t.j. pre takéto druhy je charakteristická ovoviviparita. U niekoľkých druhov sa pozoruje skutočný živý pôrod: cievy žĺtkového vaku sú úzko spojené s cievami vajcovodu a embryo dostáva významnú časť živiny z tela matky. Oviparous sa častejšie pozoruje u druhov bežných v severných a horských oblastiach. Je zrejmé, že ovoviviparita je ochrana vyvíjajúcich sa embryí pred ochladením a inými nepriaznivými podmienkami. Neexistuje jediný druh plazov, u ktorých sa vajíčka vyvíjajú vo vode. Dokonca aj pri dirigovaní plazov najviacživot vo vode (všetky krokodíly, veľa korytnačiek, niektoré hady), kladenie vajíčok a vývoj prebieha len na súši. Morské hady, ktoré nikdy neopúšťajú vodu, sú živorodé.
Vajcia a embryá plazov majú množstvo úprav pre vývoj na súši. Po prvé, vajcia sú pokryté škrupinami a inými membránami, ktoré ich chránia pred vysychaním a poškodením. Po druhé, embryá vyvíjajú špeciálne embryonálne membrány (serózne, amnionové a alantois), ktoré sú veľmi dôležité pre zabezpečenie embryonálny vývoj zvieratá na suchej zemi. Seróza a amnion sa vyvíjajú z rovnakého dvojitého záhybu tela embrya, ktorý pochádza z ektodermu a mezodermu. Tento záhyb sa objaví z dvoch protiľahlé strany vo forme rudimentov, ktoré rastú k sebe a uzatvárajú sa nad embryom. Vonkajšia časť záhybu sa mení na serózu (jej vonkajšia vrstva je ektodermálna, vnútorná je mezodermálna) a vnútorná časť sa mení na amnion (jej vonkajšia vrstva je mezodermálna, vnútorná je ektodermálna). Dutina tvorená amniónom je naplnená vodnou kvapalinou, vďaka čomu dochádza k vývoju vo vlhkom prostredí a embryo je chránené pred vysychaním a účinkami rôznych trasení, obzvlášť nebezpečných na vzduchu.
Pôvod. Prvé plazy, ktoré vznikli v druhej polovici karbónu, žili v mokrých biotopoch pôdy a pravdepodobne sa množili a vyvíjali vo vode. V tom čase už bola krajina obývaná nižšími suchozemskými rastlinami, rôznymi bezstavovcami, začalo sa dobývanie vzduchu hmyzom. Plazy, ktoré sa z vodných plôch mohli dostať ďalej ako obojživelníky, mali bohatú potravu a nestretli sa s vážnymi nepriateľmi. Koncom karbónu a opakovane počas permu sa klíma stávala suchšou, čo spôsobilo úhyn mnohých obojživelníkov a naopak prispelo k progresívnemu rozvoju a širšiemu rozšíreniu plazov na súši. Po určitom čase prešli na rozmnožovanie a vývoj mimo vodných plôch, najskôr vo vlhkejších suchozemských biotopoch a potom na suchších miestach. Počas druhohôr sa rozšírili po celej zemeguli a mnoho rôzne skupiny. Spolu s predátormi "bolo veľa bylinožravých druhov, objavili sa plazy prispôsobené na let a plazy žijúce v moriach. Ich telesná veľkosť bola rôzna, ale väčšinou išlo o veľké zvieratá a niektoré boli gigantické - až 20 m alebo viac na dĺžku. Podnebie počas takmer celej éry druhohôr bolo teplo, veľmi priaznivé pre tieto živočíchy, ktoré nemajú dostatočne vysoké konštantná teplota telo a strácajú svoju aktivitu, keď sa prostredie ochladí.
Na konci druhohôr, glóbus prebiehali silné horotvorné procesy sprevádzané prudkou zmenou podmienok existencie (teplota, vlhkosť atď.). To viedlo k hromadnému vyhynutiu druhohorných plazov, ktorí nedokázali vydržať boj o existenciu v takých pre nich ťažkých abiotických podmienkach prostredia, komplikovaných konkurenciou s cicavcami a vtákmi. Zároveň v kenozoickej ére vznikali a rozmnožovali sa nové skupiny plazov – z nich potom pochádzali jašterice a hady. Zo starých skupín prežil iba jeden veľmi primitívny druh primárnych jašterov, malý počet krokodílov a korytnačiek. Moderné plazy sú rozšírené najmä v teplých krajinách, najmä v subtropických a tropických krajinách, v krajinách s miernym podnebím je ich málo a na ďalekom severe chýbajú.

