5-10-2012, 19:14
Funkcije korteksa
Čelni reženj. Glavna funkcija čelnih režnjev je nadzor nad prostovoljnimi gibi, koordinacija motoričnih mehanizmov izraznega govora, pa tudi zagotavljanje "ustvarjalnega" ali abstraktnega razmišljanja.
Čelni reženj lahko razdelimo na pet regij:
- motor,
- premotor,
- čelni motorični vid,
- Brocino motorično področje govora
- in frontalno asociacijsko območje
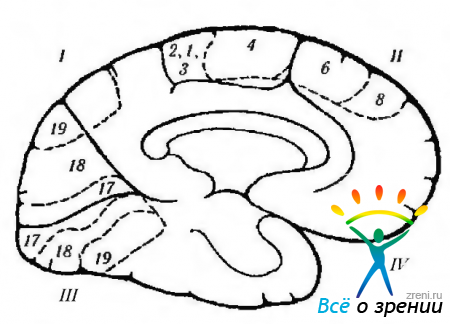
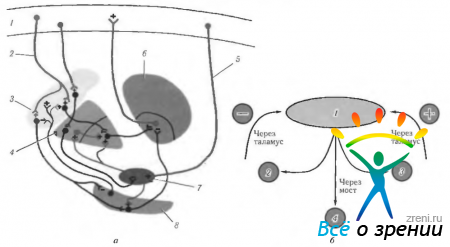
riž. 4.1.12. Shematski prikaz funkcij korteksa veliki možgani, lokalizirane na medialni površini, in njihova kršitev med razne bolezni. ki ga spremlja vzbujanje ali uničenje skorje (po Crosby et al., 1962): 1 - parietalni reženj. Centralni girus (4). levo okcipitalni predel: -aleksija brez agrafije. Posteriorni korteks: -težave pri prevajanju pogleda; - Balintov sindrom (dvostranski), 11 - čelni reženj. Dodatno motorično polje (6): + dvig nasprotne roke, obračanje oči in glave k roki; + krčenje zenice; + zakasnitev govora. Prednje motorično očesno polje (8). Belt gyrus: + razširitev zenice; + solzenje. orbitalno površino. III - okcipitalni reženj. primarni vidni korteks. Kontralateralni vid: - desnostranska hemianopsija. IV-temporalni reženj: paraliza gibanja oči; ipsilateralna hemiplegija; hidrocefalus; decerebralna togost; pedunkularna halucinoza; kontralateralna hemianopija; parinaudov sindrom. Pod površjem: - prepoznavanje obraza. Opomba. Citoarhitektonska polja ustrezajo Brodmannovi klasifikaciji. Plus označuje rezultat električne stimulacije skorje, minus pa rezultat uničenja polja.
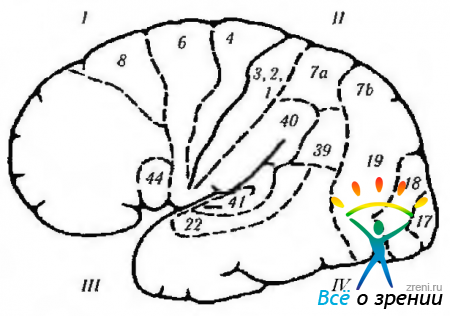
riž. 4.1.13. Bočna površina možganov (oznake so podobne tistim na prejšnji sliki): 1 - čelni reženj. Motorno polje (4). Premotorično polje (6). Motorično očesno polje (8): + kontralateralna sakada; + zapiranje nasprotnega očesa; + gomolateralna prijazna deviacija očesa. Motorično govorno polje (44): - motorična (motorična) afazija. 11 - parietalni reženj: - (- preprosta halucinoza; - iluzije; - metamorfopsija: - izguba pozornosti; - agnozija; - apraksija; - oslabitev optokinetičnega nistagmusa; - hemianopija spodnjega kvadranta. Supramarginalni girus (40): - amnezija; - žargon. afazija Angularni girus (39): - agrafija; - akalkulija; - izguba sposobnosti razlikovanja med levo in desno. desna stran; agnozija prstov. somatska občutljivost. III-temporalni reženj: + vidne halucinacije; + deja vu; + oslabljen občutek za vonj; + epileptični napadi; + avtomatizmi; - zgornja kvadrianopsija; -izguba spomina; - Klüver-Bucyjev sindrom. primarni slušni korteks. Sekundarni slušni korteks: -slušna afazija. Planum temporale: - disleksija. IV-okcipitalni reženj (17, 18, 19): + občutek svetlobe; -(-občutek barv; -(-občutek gibanja; + občutek linearnih slik. Vidna asociativna polja (18, 19): -(-kontralateralni gibi oči, kot je gladko sledenje; + vergenčni gibi; -Antonov sindrom
Primarno motorično področje(motorično področje; 4. polje po Brodmanu).
motorično področje b je pred osrednjim sulkusom, ki ločuje čelni reženj od temenskega režnja. Sestavljen je iz velikega števila Betzovih velikanskih piramidnih celic.
Predstavlja gibe, ne posameznih mišic. Celice, ki nadzorujejo gibe, so nameščene tako, da so gibi spodnje polovice telesa predstavljeni v zgornjem delu girusa, zgornje polovice pa v spodnjem. Nevronski aksoni se spuščajo skozi notranjo kapsulo in peclje možganov v obliki kortikobulbarnega in kortikospinalnega trakta.
Odstranitev motoričnega korteksa vodi do paralize.
Premotorično področje(polje b) leži neposredno pred primarnim motorični korteks, ki zaseda zadnji del treh vodoravno usmerjenih čelnih vijug.
Tvori vlakna frontopontocerebelarnih traktov in daje tudi številna vlakna bazalnim ganglijem.
Domneva se, da je premotorični korteks vključen v načrtovanje motoričnih dejanj. To dokazuje dejstvo, da so nevroni šestega polja vznemirjeni šele, ko razmišljajo o nameravanem dejanju. Stimulacija teh nevronov povzroči dvig rok nasprotna stran, glava in oči pa se obrnejo v smeri dvignjenega uda. Zenice se razširijo in govor je moten (glej sliko 4.1.12, 4.1.13).
Uničenje tega območja spremlja psevdocerebelarna ataksija nasprotne strani. Oslabljena je tudi sposobnost kopiranja naučenih gibov (apraksija). V tem primeru posamezni gibi niso poškodovani, vendar jih bolnik ne more izvesti v želenem zaporedju.
Prednje motorično vidno območje(polje 8 po Brodmanu).
To območje (polje 8 in delno 6 in 9) zavzema zadnji del medialne čelne vijuge (gyrus frontalis medius).
Prednje motorično vidno območje pomembna pri prostovoljnih gibih oči. Nekatera področja definirajo konvergenco, divergenco in akomodacijo.
Treba je opozoriti, da je mogoče med električno stimulacijo skoraj vseh predelov možganskih hemisfer zaznati gibanje oči, zenice in vek. Električna stimulacija frontalnega motoričnega očesnega področja vodi do sakadičnih gibov, v nasprotju z odzivom na stimulacijo polj 18, 19 in 22 (gladko sledenje in vergenčni gibi). Stimulacijo 8. polja pri opicah spremlja sočasna kontralateralna deviacija oči. V tem primeru se zenica razširi, glava in trup pa se obrneta v nasprotno smer. Poškodba čelnega režnja povzroči reverzibilno izgubo sposobnosti kontralateralnih sakadičnih gibov.
Ta predel skorje odlikujejo številne povezave.
Aferentna vlakna vstopijo v vidni frontalni korteks iz vidnega tuberkula, pa tudi iz zgornje temporalne, peristriatalne in parietalne (območje 7) skorje. Tu so bili najdeni nevroni, ki nadzorujejo gibanje oči.
Eferenti projiciran na bazalne ganglije, talamus, pretektalno regijo (vključno z jedrom optičnega trakta), zgornje kolikule kvadrigemina in del retikularna tvorba most.
Po drugi strani pa je treba opozoriti, da so frontalna vidna področja projicirana na kontralateralna frontalna področja in na kortikalna področja z iste strani. Omogočajo vizualno zaznavo (na 7. polju parietalni korteks in peristriatalni korteks).
Ko se projicira na srednji možgani vlakna prefrontalnega korteksa prečkajo sprednji del interne kapsule in se v rostralnem diencefalonu delijo na dorzalno in ventralno pot.
Dorzalna transtalamična pot prečka dorzomedialno in intralamilarno jedro optikusa talamusa in srednji del vzglavnika ter hkrati tvori ne veliko število sinaptične povezave. Sinapse najdemo tudi v pretektalnih jedrih in v zgornjih tuberkulah kvadrigemine.
Ventralna pedunculo-tegmentalna pot poteka v najbolj ventralnem delu možganskega debla, oskrbuje z vlakni jedra hipotalamusa in globlje - v zgornje tuberkuloze kvadrigemine. Istočasno imajo frontalna vidna območja topografsko projekcijo na zgornje tuberkule kvadrigemine. Pot se konča v retikularni tvorbi mostu.
Tretji prefrontalni ligament se pojavi v območju diencefalično-mezencefalnega stičišča, ki se nahaja v bližini rdečega jedra. Ta pot je projicirana na jedro medialnega longitudinalnega fascikulusa in intermediarno Cajalovo jedro na isti strani, pa tudi na Darkshevichevo jedro in rastralni del okulomotornega jedra na obeh straneh.
Različna področja frontalne vidne skorje nadzorujejo sakadične gibe oči. drugačna amplituda, kot tudi smer sakad. Zagotavljajo tako imenovano "vizualno pozornost", usmerjanje oči na "cilj".
Trenutno poteka preiskava vloga čelnega režnja pri delovanju vek. Prostovoljno zapiranje vek zahteva sprostitev dvigalke veke in kontrakcijo orbikularne očesne mišice. Obstaja polemika glede lokalizacije področij možganov, ki nadzorujejo voljne gibe vek. Ta mesta se lahko nahajajo v motoričnih premotornih ali frontalnih motoričnih vidnih področjih. Stimulacija tega področja vodi do zaprtja kontralateralne veke. Enostransko poškodbo nedominantnih čelnih režnjev lahko spremlja obojestransko neuspešno zapiranje vek.
Brocino motorično govorno področje(polja 44 in 45 po Brodmanu).
To območje se nahaja v posteriorno-spodnjem delu čelnega režnja. Prispeva k začetku govora. Poškodba območja Broca vodi v afazijo, ki je sestavljena iz težav pri povezovanju v določenem zaporedju besed, da se zagotovi soroden govor (ekspresivna afazija). Bolniki razumejo pisni in govorni jezik, vendar običajno ne morejo ničesar izgovoriti. Motena je tudi sposobnost pisanja, čeprav se roka lahko uporablja za druge dejavnosti.
frontalno asociacijsko območje. To pomembno področje v sprednji polovici čelnega režnja je odgovorno za sprožitev vseh vrst vedenjsko-intelektualnih, visceralnih in čustvenih funkcij. Spremembe v spominu, inteligenci in osebnosti se pojavijo z draženjem ali masivnim uničenjem čelnih režnjev. V teh primerih je to še posebej izrazito. ko je prizadeta dominantna polobla.
Olfaktorni trakt se nahaja pod orbitalno površino čelnega režnja in je edina senzorična pot, ki doseže korteks brez tvorbe sinaps v talamusu. Enostranski tumor orbitalne površine čelnega režnja (meningiom olfaktornega sulkusa ali meningiom manjšega krila) sphenoidna kost) lahko privede do stiskanja vohalnega trakta in optični živec. Posledično se razvije anosmija (pomanjkanje vonja), istostranska atrofija vidnega živca in otekanje glavice vidnega živca na nasprotni strani (sindrom Kennedyja Fosterja).
Poškodba struktur limbičnega sistema v čelnem režnju, zlasti cingularnega girusa, lahko povzroči motnje na čustveni plati življenja, pa tudi visceralne znake, vključno z brezvzročnim solzenjem in spremembami v odzivu zenic na svetlobne dražljaje.
parietalni reženj(glej sliko 4.1.12, 4.1.13). Na splošno je funkcija parietalnih režnjev povezana z občutljivostjo, spominom, povezanim z govorom in učenjem, pa tudi z orientacijo telesa v prostoru z obdelavo vizualnih informacij.
V sprednjem parietalnem režnju je girus, ki se nahaja vzporedno s precentralnim girusom ali motorično skorjo. To je postcentralni girus ali primarna somatosenzorična skorja (polja 3, 1 in 2) (glej sliko 4.1.13). Preostali del parietalnega režnja lahko razdelimo na dva lobula - zgornji in spodnji. Polji 5 in 7, ki se nahajata na površini zgornjega parietalnega lobula, sta povezani s korelacijo somatosenzoričnih informacij, kar omogoča zavestno oceno teže, površinskih lastnosti, velikosti in oblike predmeta.
Spodnji parietalni lobule(supramarginalni in kotni girus) je tesno povezan z govornimi mehanizmi, saj uničenje tega področja skorje na levi hemisferi vodi do motenj zaznavanja govora.
Vloga parietalnega režnja pri prostorski orientaciji še ni raziskana. Vendar pa je pri bolnikih z njegovim porazom opažena kršitev te funkcije.
Aferentna vlakna prehajajo skozi parietalni reženj in povezujejo področji 18 in 19 s čelnimi režnji in možganskim deblom. Te poti vključujejo zgornji in spodnji vzdolžni snop.
Predpostavlja se, da parietalni korteks (polje 7) zagotavlja funkcijo "pozornosti" na vizualne cilje.
Sedmo polje sprejema aferente od cingularni girus(g. cinguli), iz pretektalne regije in zgornjih tuberkulusov kvadrigemine skozi jedra talamusa. Najdemo tudi aferente, ki gredo od frontalnega vidnega področja in prestriatne skorje do skorje parietalnega lobula.
Eferenti so projicirani na pretektalno regijo, zgornje tuberkule kvadrigemine, sivo snov, ki se nahaja okoli silvijevega akvadukta, in na parietalne režnje obeh hemisfer možganov.
Ko se oči premikajo, se aktivirajo nevroni spodnjega temenskega režnja, kar kaže na njihovo vlogo pri nadzoru prostovoljnih gibov oči. Aktivnost se nadaljuje ves čas opazovanja tarče.
Celice 7. polja ne določajo sakadnih gibanj, saj se njihova aktivnost med sakadami ne poveča.
Stimulacija kotnega girusa(g. angularis) vodi pri poskusnih živalih do prijaznega odstopanja oči. To sta polji 38 in 39, ki potekata okoli zadnjega roba srednje temporalne vijuge (g. temporalis medius). Operativno odstranjevanje to področje skorje ne vodi do motenj gibanja oči. Dvostranska poškodba parietalne skorje lahko povzroči razvoj motorične apraksije očesa.
Poraz parietalnega režnja spremlja tudi kršitev vizualnih motoričnih mehanizmov, ki zagotavljajo optokinetični nistagmus, med katerim oči proizvajajo sakade za iskanje in fiksiranje cilja.
Drugi motorični znaki izoliranih lezij parietalnega režnja vključujejo prijazen odklon oči v nasprotni smeri od mesta poškodbe s prisilnim zapiranjem vek, nizko gibljivostjo bolnika in slabim nadzorom (nerazumevanjem) okončin nasprotne strani. Težave pri razumevanju narave delovanja udov vodijo do ustavne in okulomotorne apraksije. Bolniki s poškodbo parietalnega režnja imajo lahko težave pri premikanju pogleda z enega vidnega predmeta na drugega znotraj kontralateralne polovice vidnega polja.
Drug znak disfunkcije parietalnega režnja je nezmožnost fiksiranja vizualnega predmeta.
Poleg senzoričnih funkcij, ki so lokalizirane v posteriorni ustnici Rolandovega osrednjega sulkusa, je parietalni reženj mesto kompleksnega procesa prepoznavanja in razumevanja. Difuzna poškodba parietalnih režnjev se konča z agnozijo, vizualno nepazljivostjo, metamorfopsijo, fotopsijo, halucinacijami, iluzijami in aleksijo (glej sliko 4.1.12, 4.1.13).
več podrobnosti o posledicah disfunkcije parietalnega režnja so podani v nevrološki literaturi. Tukaj predstavljamo le nekatere izmed sindromov.
vidna agnozija, ki se pogosto razvije s kršitvijo krvnega obtoka v centralnem živčnem sistemu, je značilna kršitev procesa prepoznavanja. Agnozija je lahko popolna ali delna. Vizualna agnozija se diagnosticira v primerih, ko lahko pacient identificira predmet samo na otip. Prepoznavanje in razumevanje vizualnih podob ter barvnega pomena podobe sta pogosto motena.
Dvostranske poškodbe parietalnega režnja (pogosteje pri senilni demenci, mehčanje temporo-okcipitalnih in parietalnih območij skorje, poškodbe zadnjih delov obeh hemisfer, encefalitis) spremljajo vizualna agnozija in vizualna ataksija (Balintov sindrom).
Vizualna prostorska agnozija značilne so težave pri ocenjevanju velikosti predmeta in razdalje do njega. Opažen je tudi pojav nepravilne rešitve "sintetičnih" prostorskih problemov, kot je branje zemljevida. Simultana agnozija se zmanjša na dejstvo, da bolniki ohranijo sposobnost prepoznavanja posameznih predmetov, vendar nimajo sposobnosti zaznavanja skupine predmetov (slik) kot celote.
Asterognostija značilen po tem, da lahko pacient analizira predmete po njihovem videzu, vendar jih ne more otipljivo označiti. To se razvije kot posledica motenj kompleksne interakcije med taktilnimi in vizualnimi informacijami v parietalnih režnjih korteksa.
Difuzne lezije parietalnih režnjev lahko povzročijo iluzije in metamorfopsije. Ta izkrivljanja vključujejo mikropsijo, makropsijo. Bolniki opazijo premikanje nepremičnih ciljev, napačno ocenijo razdaljo do predmetov. TO posebne vrste iluzije vključujejo papinopsijo (kršitev vizualne percepcije v obliki ohranjanja ali ponovnega pojava vizualne podobe po izginotju predmeta iz vidnega polja), halucinacijske poliopije itd.
Iluzije se razvijejo kot posledica difuznih motenj skorje, halucinacije in fotopsije pa z razvojem tumorja ali toksične poškodbe skorje.
Značilnosti patologije parietalnega režnja so lahko odvisne tudi od tega, ali je patološko žarišče lokalizirano v dominantni (običajno levi) ali nedominantni polobli. Prevladujoči parietalni reženj je odgovoren za razvoj govornih motenj, kot so slušno senzorična afazija, vidna agnozija in agrafije.
Poškodba prevladujočega kotnega girusa(pogosteje kot posledica motenj krvnega obtoka v bazenu angularne arterije) razvijejo nistagmus, vizualno-agnostične motnje, agrafijo, akalkulijo (nezmožnost seštevanja ali odštevanja števil), digitalno agnozijo, nezmožnost razlikovanja med desno in levo stranjo ( Gerstmannov sindrom; sindrom angularne vijuge). Morda razvoj homonimne hemianopsije.
Če je nedominantni kotni girus poškodovan, je interakcija bolnika z okoljem motena. Posledično se razvijeta vidna dezorientacija in topografska agnozija.
Spredaj od kotnega gyrusa in čez supratemporalni sulkus je supramarginalni girus. Na dominantni hemisferi je to območje stičišče vidnih in slušnih komponent jezika.
Pogosto se konča poškodba posteriornih parietalnih asociacijskih območij razvoj apraksije(težave pri izvajanju "finih" specializiranih gibov). To je očitno posledica nerazumevanja potrebnega zaporedja gibov za izvedbo določene funkcije (tj. Pacientu ni jasno, kaj in v kakšnem zaporedju je treba izvesti). Bolnik nima sposobnosti risanja preprostega diagrama (konstruktivna apraksija). Manj resna oblika te patologije je taktilna agnozija (nezmožnost prepoznavanja predmeta na palpacijo). Včasih se ta bolezen imenuje asterognostija. Hkrati ni znakov izgube taktilne ali proprioceptivne občutljivosti, integracija vidnih in somatosenzoričnih informacij je motena. Apraksija in asterognostija se pogosto razvijeta po poškodbi leve hemisfere možganov. Asterognostija je običajno omejena na nasprotno stran telesa, medtem ko je apraksija običajno dvostranska.
Vidne in slušne informacije so tesno povezane s funkcijo jezika. Iz tega razloga poškodbe območij 39 in 40 ter delov območja slušne asociacije običajno povzročijo afazijo, znano kot Wernickejeva afazija(Wernicke). Če je poškodba omejena na angularni girus, se izgubi sposobnost razumevanja pisnega jezika (aleksija) in sposobnost pisanja (agrafija). Hkrati bolniki razumejo govor. To stanje se običajno razvije po poškodbi.
Integracija vizualnih in somatosenzoričnih informacij je pomembna za oblikovanje "podobe telesa", to je za razumevanje značilnosti položaja telesa v prostoru. Obsežna poškodba polj 7, 39 in 40 ene hemisfere lahko privede do razvoja "nerazumevanja" ali "zanemarjanja" nasprotne polovice telesa. Pacienti si ne morejo previti ali odvezati lastnih okončin. Prav tako ne prepoznajo lastnih udov.
Okcipitalni reženj(polja 17, 18 in 19). Vzdolž brazde (fissura calcarina) je primarno vidno območje (polje 17). Poleg njega je "ocenjevalno" vizualno območje (polje 18), za katerega se verjame, da pretvarja signale, ki jih prejme polje 17, v vizualne slike.
Območje 17 okcipitalnega korteksa je območje sprejema in obdelave vizualnih informacij in ima pomembne povezave z motoričnimi vizualnimi področji korteksa. Na tej ravni vidne skorje se oceni pomen videnega. Draženje tega področja povzroča halucinacije in sanjske slike.
Pozornost si zaslužijo tudi motorične povezave okcipitalnega režnja, saj sodelujejo pri optokinetičnem nistagmusu, akomodacijskem refleksu in regulaciji binokularnega vida.
Polji 18 in 19 sta soodvisni in imata več povezav z ostalimi deli možganov. Domneva se, da je področje 18 bolj zainteresirano za integracijo vizualnih informacij. prejme v 17. polju, polje 19 pa to informacijo prevede v kompleksnejšo mentalno in motorično aktivnost.
Nedavni anatomski in fiziološke raziskave razkrila, da obstajajo vsaj, 10 podregij polj 18 in 19, katerih nevroni imajo sposobnost odzivanja na vizualne dražljaje. Vsaka od teh podregij je povezana s 17. poljem. Vendar med seboj niso povezani. Poškodbe teh podregij ne vodijo v slepoto. Nekateri so kršeni vizualne funkcije, kot je analiza barv, zaznavanje celovitosti predmeta itd.
vidna asociacijska skorja, ki se nahaja na nekaterih področjih temporalnega režnja, sodeluje tudi pri obdelavi vizualnih informacij. To v največji meri velja za polje 21 (v srednji temporalni skorji). To polje zagotavlja prepoznavanje slike, njegova poškodba pa se konča z agnozijo.
Navedene lastnosti vidne skorje zagotavljajo tudi številne horizontalne in vertikalne povezave med nevroni v različnih plasteh vidne skorje. Zaradi njihove prisotnosti je opažena izjemna plastičnost vidne skorje pri obdelavi vizualnih informacij različnih vrst.
Strukturna organizacija vidne skorje in njene funkcije bodo podrobneje podane v naslednjem razdelku.
temporalni reženj(polja 41, 22).
Ta del možganov povezana z zaznavanjem slušnih občutkov, sodeluje pri govorni funkciji preko slušnega nadzora govora, igra pa tudi vlogo pri vrednotenju prostora in spominske funkcije. Že dolgo je ugotovljeno, da je transverzalni temporalni girus (polje 41) primarni slušni center. Malo polje (22). ki obdaja to vijugo, velja za "mentalno" ali "ocenjevalno", slušni center. Draženje večjega dela temporalnega režnja, zlasti vzdolž srednje temporalne vijuge, povzroči občutke, ki jih bolnik opisuje kot slušne. Bolniki s poškodbo "mentalnega polja" temporalnega režnja včasih izgubijo sposobnost ocenjevanja zvočnih tonov. Hkrati pacient ne prepozna motivov, glasbene zvoke pa zaznava kot naključen hrup.
Polje 22 leve poloble je povezano tudi z govorno funkcijo, saj poškodba tega področja vodi do izgube sposobnosti razumevanja pomena besed. Obstajajo tudi dokazi, da je temporalni reženj povezan z "vestibularno" občutljivostjo (občutek za ravnotežje), saj stimulacija posteriornega zgornjega temporalnega girusa pri zavestnem bolniku povzroči omotico in občutek vrtenja.
Globoke lezije v temporalnem režnju pogosto vključujejo najnižja optična sevalna vlakna iz lateralnega genikulatnega telesa. pri čemer okvara vida. Najpogostejša je hemianopija zgornjega kvadranta.
S porazom temporalnega ali psihomotornega režnja se razvije epilepsija, ki jo spremljajo vidne halucinacije, občutek deja vu in deja pense (»že videno« - fr.), pri katerem imata vizualna izkušnja in misli čuden prijateljski odnos, pojavijo se avtomatizmi, ki spremljajo amnezijo, kompulzivno vedenje, jezo in tesnobo.
Pri bolnikih z enostransko poškodbo primarnega slušna skorja rahla izguba sluha. V primeru kršitve slušnega prepoznavanja (zvočna agnozija) so prizadeta asociativna slušna polja. Bolniki običajno poročajo, da slišijo zvoke, vendar jih ne morejo identificirati.
Območje, ki sodeluje pri prepoznavanju obraza, se nahaja na ventralni površini temporalnega režnja. Poškodba tega območja (pogosteje s kršitvijo krvnega obtoka v območju bazilarne arterije možganov), ki se praviloma razširi na sosednji okcipitalni reženj (polja 20, 21), obe polobli, spremlja razvoj prozopagnozije(nezmožnost prepoznavanja obrazov). Na splošno tega stanja ne spremljajo drugi nevrološki znaki. Pacient zna brati in poimenovati predmete. V primerih, ko pacient sliši glas znane osebe, jo lahko takoj identificira.
Preostali del zgornjega temporalnega gyrusa zaseda območje 22 (slušna asociativna skorja), ki prejme veliko število aferentov iz polj 41 in 42, ki dajejo vlakna v parietalni in okcipitalni reženj. Območje 22 je vključeno tudi v jezikovne funkcije. Poškodba zgornjega dela 22. polja povzroča težave pri razumevanju jezika.
Temporalni režnji so še posebej pomembni pri spominski funkciji.. Na primer, stimulacija slušne asociacijske skorje pri bolnikih med nevrokirurškimi operacijami prikliče kompleksne spomine, tako slušne kot vidne. Pri obsežni poškodbi temporalnega režnja pride do motenj spomina.
limbični reženj(slika 4.1.14).
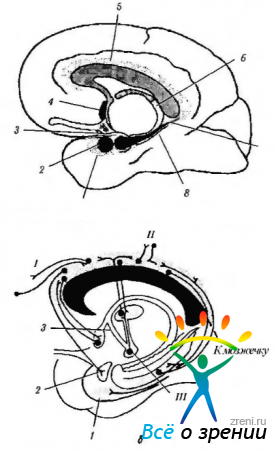
riž. 4.1.14. Limbični sistem (po Brodslu, 1992): a - lokacija struktur limbičnega sistema (1 - entorialno polje; 2 - amigdala; 3 - kavelj; 4 - septalno jedro; 5 - cingularni girus; b - forniks; 7 - hipokampus; 8 - parahipokampalni girus); b - povezave cingularnega gyrusa (I - frontalna asociativna skorja; II - parietalna in temporalna asociativna skorja; III - mastoidno telo). Cingulatni girus je povezan z asociativnimi polji možganske skorje in limbičnega sistema, kar zagotavlja interakcijo med njimi.
Limbični sistem je kompleks tvorb terminala, diencefalona in srednjih možganov. Pravzaprav " limbični sistem” sovpada s prej uporabljenim konceptom - “vohalni možgani”.
Če nova skorja (neokorteks) nadzoruje prostorsko-časovne odnose organizma z okoljem, odgovorna pa je tudi za formalno-logično mišljenje in stereognostične sposobnosti, potem je limbični sistem določa čustvene funkcije in motivacijo za delovanje kot tudi procese učenja in spomina. Prav tako nadzoruje homeostazo. Njegova pomembna funkcija je organizacija vedenjskih reakcij posameznika kot odziv na vplive zunanjega okolja in spremembe v notranjem okolju organizma. Te reakcije so namenjene ohranjanju posameznika in potekajo preko spodnjih centrov diencefalona.
Limbični reženj ima tudi nekaj opraviti s funkcijo vonja, saj draženje ali poškodbo hipokampalne regije spremlja občutek za vonj.
Nastane limbični reženj:
- subkavzalni predel (area subcallosa),
- cingularni girus (gyrus cinguli),
- isthmus cingulate gyrus
- parahipokampalna vijuga (gyrus parahippocampalis),
- hipokampus (amonov rog, zobati girus in osnova hipokampusa ali subikulum)
- in Broca diagonalni ligament.
Subkortikalne strukture limbičnega sistema vključujejo tonzile (corpus amygdaloideum), septalna jedra in sprednje talamično jedro.
Aferentne in eferentne povezave struktur limbičnega sistema, tako med seboj kot z drugimi deli možganov, so izjemno raznolike. Njegove vzajemne povezave s hipotalamusom so najbolj izrazite.
Hipotalamus in mamilarna telesa povezan s hipokampusom in septalnim predelom prek forniksa, z amigdalo prek terminalne strije in amigdalofugalne snope ter z vohalnimi možgani preko medialnega snopa prednji možgani. Limbični sistem je povezan s srednjimi možgani preko hipotalamusa in mamilarnih teles.
Limbični sistem komunicira z neokorteksom v frontalni in temporalni reženj. Temporalne regije so v glavnem odgovorni za prenos informacij iz vidne, slušne in somatske skorje v amigdalo in hipokampus. Čelni predeli služijo kot glavni del neokorteksa, ki uravnava limbične cone.
Bazalni gangliji predstavljajo subkortikalno kopičenje nevronov v obliki skupine jeder (sl. 4.1.15; 4.1.16).
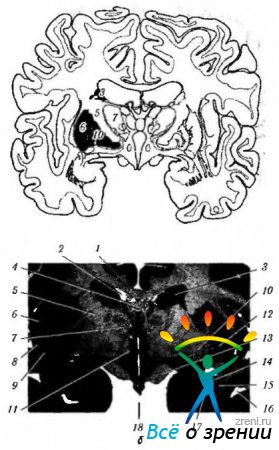
riž. 4.1.15. Bazalni gangliji: a - shematski prikaz lokacije struktur, povezanih z bazalnimi gangliji; b - čelni del možganov, ki poteka skozi strukture, povezane z bazalnimi gangliji (4 - corpus callosum; 2 - rog stranski ventrikel; 3- telo repnega jedra; 4-trezor; 5-notranja kapsula; 6 - otoček; 7 - vidni tuberkel; 8- lupina; 9 - ograja; 10 - bleda žoga; 11 - hipotuberozna regija; 12 - brezimna snov: 13 - vizualni trakt; 14 - zadnji spodnji del kavdatnega jedra; 15 - tonzila: 16 - temporalni rog lateralnega ventrikla; 17 - čelni rog lateralnega ventrikla; 18 - tretji prekat)
riž. 4.1.16. Najpomembnejše aferentne, eferentne in notranje povezave bazalnih ganglijev in njihova povezava z malimi možgani (razlaga v besedilu): a - (1-motorna skorja; 2 - eferentna vlakna od skorje do striatuma; 3 - striatum; 4 - bleda krogla; 5 - aferentna vlakna; 6 - vidni tuberkel; 7 - subtalamično jedro; 8 - substantia nigra); b - (1 - možganska skorja (motor); 2 - bazalni gangliji; 3 - mali možgani; 4 - motorični in interkalarni nevroni)
Nadzorujejo gibanje telesa. Ta funkcija bazalnih ganglijev je bila ugotovljena na podlagi kliničnih opazovanj. Ko so poškodovani, pride do kršitve motorična aktivnost tako v obliki nezmožnosti sprožitve potrebnih gibov kot nezmožnosti zatiranja gibov.
Pri Parkinsonovi bolezni, ki nastane ob poškodbi bazalnih ganglijev, imajo bolniki "neizrazit" obraz. To je povezano z oslabljeno motorično aktivnostjo obraznih mišic in nadzorom gibov oči, zlasti motnjami sakadičnih gibov. Prav zaradi tega si bomo podrobneje ogledali nevroanatomijo tega področja.
Obstajajo tri glavne kopičenja subkortikalnih jeder, ki se imenujejo striatum ( corpus striatum), ograjo (claustrum) in amigdalo (corpus amigdaloideum).
- Corpus striatum je sestavljen iz dveh delov - repnega jedra (nucleus caudatus) in lentiformnega jedra (nucleus lentiformis).
Repno jedro leži nad in medialno od lentikularnega jedra, ločeno od slednjega s plastjo bele snovi, imenovano notranja kapsula (capsula interna). Zadebeljeni sprednji del repnega jedra, njegova glava (caput nuclei caudati) tvori stransko steno sprednji rog lateralnega prekata, medtem ko se zadnji tanek del repnega jedra (corpus et cauda nuclei caudati) razteza nazaj po dnu osrednjega dela lateralnega ventrikla. Na medialni strani nucleus caudatus meji na vidni nasip, ločen od njega s trakom bele snovi (stria terminalis). Spredaj in spodaj glava repnega jedra doseže sprednjo perforirano snov (substantia perforata anterior). Na tej točki je glava povezana z lentikularnim jedrom (z delom, imenovanim putamen). Poleg te široke povezave obeh jeder na ventralni strani so še tanki trakovi sive snovi, prepredeni z belimi šopi notranje kapsule. Iz njih je nastalo ime "progasto telo" (corpus striatum).
Lentikularno jedro(nucleus lentiformis) leži lateralno od repnega jedra in vidnega nasipa, ločen od njih z notranjo kapsulo. Na vodoravnem delu hemisfere ima medialna površina lentiformnega jedra, obrnjena proti notranji kapsuli, obliko krhkosti z vrhom, usmerjenim proti sredini. Sprednja stran kota je vzporedna s kavdatnim jedrom, zadnja stran pa s talamusom. Bočna površina je rahlo konveksna in obrnjena proti stranski strani poloble v predelu insule. Spredaj in ventralno, kot je bilo že omenjeno, se lentiformno jedro združi z glavo kavdatnega jedra.
Na čelnem delu ima lentiformno jedro obliko klina, katerega vrh je obrnjen na medialno stran, osnova pa bočno. Lentikularno jedro je razdeljeno na tri segmente z dvema vzporednima belima plastema (laminae medullares). Bočno nameščeno temno sivo območje se imenuje lupina (putamen), oba medialna, svetlejša pa skupaj imenujemo bleda krogla (globus pallidus).
Razlikuje se že po svojem makroskopskem videzu, bleda žoga(globus pallidus) ima tudi histološko zgradbo, ki se bistveno razlikuje od zgradbe drugih delov striatuma.
Glede na vse te značilnosti ločimo bledo kroglico v posebno morfološko enoto, imenovano pallidum, medtem ko oznako striatum pustimo le za putamen in nucleus caudatus. Posledično izraz "lentikularno jedro" izgubi svoj prejšnji pomen in se lahko uporablja le v izključno topografskem smislu. Istočasno se namesto prejšnjega imena corpus striatum caudatus in lentikularna jedra imenujejo striopallidar sistem.
Striopalidarni sistem je glavni del ekstrapiramidnega sistema, poleg tega pa je najvišje regulacijsko središče avtonomne funkcije v povezavi s termoregulacijo in presnovo ogljikovih hidratov prevladuje nad podobnimi vegetativnimi centri v hipotalamusu.
- Ograja(claustrum) je tanka plošča sive snovi, ki se nahaja v predelu otoka, med njim in putamenom. Od slednjega je ločen s plastjo bele snovi, imenovano zunanja ovojnica (capsula externa).
- amigdala(corpus amygdaloideum) se nahaja pod putamenom na sprednjem koncu temporalnega režnja. Zdi se, da amigdala pripada subkortikalnim vohalnim centrom in limbičnemu sistemu. Konča se v snopu vlaken, ki prihajajo iz vohalnega režnja in sprednje perforirane snovi (substantia perforata anterior), ki je navedena v opisu vidnega hriba, imenovanega stria terminates.
Vloga amigdale pri ljudeh ni dobro razumljena. V bistvu je bila vloga amigdale razkrita na podlagi študije nevroloških bolnikov. Zahvaljujoč tem študijam je bilo ugotovljeno, da mehanska ali električna stimulacija amigdale pri ljudeh pogosto povzroči strah ali druge čustvene reakcije. Torej človeška amigdala sodeluje v procesih čustvenega zaznavanja. Dokazano je tudi, da z dvostransko poškodbo tonzil pride do izgube "čustvenega" spomina, pojava "prestrašenega" izraza obraza in kršitve integracije vohalnih in vizualnih informacij.
Znano je, da ima človek sposobnost določanja razpoloženja in čustvenega stanja drugega posameznika. IN Zadnje čase vpletenost amigdale v ta kompleks fiziološki proces. Pri določanju čustvenega stanja človeka naši možgani zajamejo in analizirajo subtilne spremembe v izrazu obraza, predvsem pa smer pogleda. Nevronski mehanizmi, na katerih temeljijo ti procesi, niso povsem jasni. Kljub temu je bilo ugotovljeno, da je "natančna" nastavitev pogleda, psihofiziološko ocenjena kot prisotnost "pozornosti", odločen vrh temporalni korteks in amigdala. V študiji bolnikov z avtizmom je bilo ugotovljeno, da imajo pogosto anomalijo amigdale ali njeno poškodbo. Neposredni mehanizmi, na katerih temelji družbena presoja drugih oseb na podlagi usmerjenosti pogleda, ostajajo večinoma neznani.
V zadnjem času se substantia nigra in subtalamična jedra pripisujejo tudi bazalnim ganglijem. Poleg tega se v substantia nigra razlikujeta 2 dela - retikularna (pars reticulata) in kompaktna (pars compacta).
Morfološke in elektrofiziološke metode so pokazale, da večina aferentni signali, ki prihajajo do bazalnih ganglijev, vstopijo v striatum (glej sliko 4.1.16). Ti signali prihajajo iz številnih virov, glavni pa so:
- vsa področja korteksa hemisfere;
- intralamelarna jedra talamusa
- substantia nigra (po dopaminergični poti).
Iz notranjosti blede oble izvira najpomembnejše od vseh eferentne poti bazalni gangliji. Ta pot se konča predvsem v vidnem tuberkulu in strehi srednjih možganov.
Tako bazalni gangliji igrajo vlogo vmesnega člena v verigi, ki povezuje motorična področja korteksa z vsemi drugimi njegovimi področji. Njihova glavna funkcija je »načrtovanje« telesne dejavnosti.
IN Zadnja leta ugotovili, da bazalni gangliji skupaj z nekaterimi kortikalnimi območji določajo tudi nekatere kognitivne funkcije, vključno s pozornostjo, spominom. Koncept "pozornosti" je v najširšem pomenu besede "proces izbire". Glede na to, da je proces orientacije v prostoru in izbira želenega cilja, pri katerem sodelujejo sakadični gibi oči, povezana z bazalnimi gangliji, postane jasna pomembna vloga bazalnega kompleksa v procesih orientacije skozi sakade. V tem primeru ima posebno mesto pri nadzoru sakad očesa substantia nigra, zlasti njen retikularni del. Tako anatomsko kot elektrofiziološko je dokazana prisotnost njegove povezave z zgornjimi tuberkulami kvadrigemine. Glavna funkcija nevronov v črni substanci je zaviranje aktivnosti nevronov zgornjega kolikulusa kvadrigemine, ki sodelujejo pri nastajanju sakad.
Pri motorični aktivnosti oči sodeluje tudi caudatus nucleus, ki ima povezave s substantia nigra in zunanjimi tuberkulami quadrigemina. Vzbujanje kavdatnega jedra vodi do sakadičnih gibov očesa v nasprotni smeri od vzbujenega jedra.
Subtalamično jedro sodeluje tudi pri gibanju oči. Na to jedro je projicirana bleda kroglica, skorja čelnega predela. Vlakna odhajajo iz subtalamičnega jedra v substantia nigra, zunanji in notranji segment globus pallidus.
Funkcija bledega globusa pri okulomotorični aktivnosti je manj jasna. To je posledica dejstva, da ima globus pallidus veliko povezav med različnimi jedri bazalnih ganglijev in malo povezav z živčni centri ki se nahaja zunaj bazalnih ganglijev. Okulomotorični nevroni so bili najdeni samo v dorzalnem delu globusa pallidusa, tj. v delu, ki prejema vhode iz caudatus nucleusa.
Vloga bazalnih ganglijev pri gibanju oči potrjena s številnimi kliničnimi opazovanji. Pri Parkinsonovi bolezni, ki jo spremlja degeneracija bazalnih ganglijev in sive snovi srednjih možganov ter prekinejo frontomezencefalne poti, se razvije progresivna supranuklearna paraliza. Pri takih bolnikih se zmanjša hitrost sakadičnih gibov oči. Sakade so običajno hipometrične, kar je povezano s kršitvijo funkcije "sakadnega spomina". Pogosto očesnega gibanja ni mogoče dokončati v celoti. Navpični gibi oči so prizadeti močneje kot vodoravni. Bolniki kažejo tudi aksialno togost in demenco. Podobne spremembe so bile ugotovljene tudi pri Huntingtonovi bolezni.
corpus callosum(corpus callosum). Corpus callosum je velik snop vlaken (sestavljen iz približno 250 milijonov vlaken), ki povezuje obe polobli (slika 4.1.17).
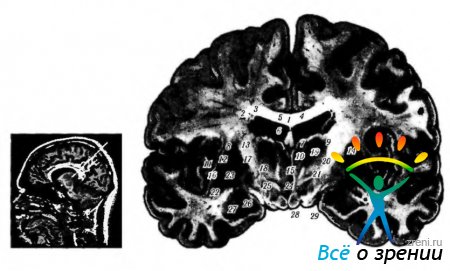
riž. 4.1.17. Lokalizacija jeder talamusa: a - jedrska paramagnetna resonanca; b - čelni del možganov (1 - telo corpus callosum; 2 - telo kaudatnega jedra; 3 - osrednji del lateralnega ventrikla; 4 - prozorni septum; 5 - horoidni pleksus stranski ventrikel; 6 - trezor; 7 - sprednja skupina jeder vidnega hriba; 8- zunanja kapsula; 9 - stranska skupina jeder talamusa; 10- medialna jedra talamusa: - lupina; 12- stranska medularna plošča; 13 - notranja kapsula; 14 - retikularno jedro talamusa; 15 - medtalamična konica; 16 - stranski del blede kroglice; 17 - medialna medularna plošča; 18 - mamilotalamični snop; 19 - HI polje postrvi; 20 - gopa incerta; 21 - H2 Postrvi; 22 - ograja; 23-medialni del blede kroglice; 24 - tretji ventrikel; 25 - jedra hipotalamusa; 26-vidni trakt; 27 - amigdala; 28-telo bradavice; 29-baza možganskih nog)
Glavna funkcija tega žarka je prenos informacij iz ene hemisfere v drugo.
Glavne funkcije corpus callosum so:
- Korelacija slik na desni in levi polovici vidnega polja.
- Integracija občutkov, ki prihajajo iz parnih okončin, potrebnih za učenje motorične koordinacije.
- Integracija pozornosti in aktivacijskih procesov v hemisferah.
Pri bolnikih, pri katerih je bil corpus callosum prerezan v terapevtske namene, je prišlo do disfunkcije možganov v Vsakdanje življenje ni bilo opaziti. Razkrili so se le skozi posebni testi. Drugi bolniki včasih razvijejo precejšnje vedenjske motnje. Najpogosteje se to kaže z »besedno slepoto«, »besedno naglušnostjo« in izgubo koordinacije gibov desnega in levega uda.
Nadaljevanje v naslednjem članku: Anatomija možganov? 3. del
In na očeh celotne prestolnice
Petelin je plapolal s pletilke,
odletel do kočije
In sedel na kraljevo krono,
Presenečen, kljuvan v krono
In poskočil ... in hkrati
Dadon je padel s kočije -
Enkrat je zadihal in umrl.
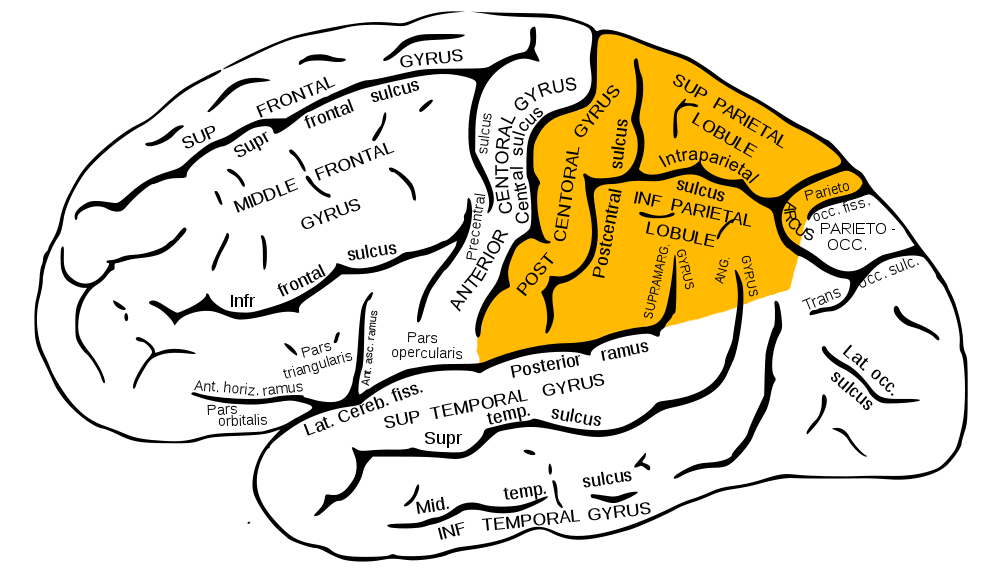
Zdi se, da ste že razumeli, o čem bomo razpravljali v naslednjem gradivu cikla "Kako delujejo možgani". Govorili smo že o čelnih režnjih, temporalnih in parietalnih, zdaj pa se obrnemo na parietalne režnje. V latinščini so tudi Lobus parietalis, ki ga poznajo zdravniki.
Parietalni režnji prikazani rumeno
Parietalni reženj se nahaja tik nad zatilnico in je "sestavljen" iz treh zavojev: enega navpičnega- zadnji osrednji (najstarejši odsek) in dva vodoravna - zgornji parietalni (novi) in spodnji parietalni (novejši).
Tako kot struktura čelnih režnjev so deli človeškega telesa "projicirani" na osrednjem sprednjem girusu parietala: spodnja tretjina je obraz, srednja tretjina je roka in trup, zgornja je noga. Ne pozabite, da je delež "podvojen", zato je ena njegova polovica odgovorna za eno (nasprotno) polovico telesa.
Struktura parietalnih režnjev
Poleg tega so v zgornjem parietalnem girusu centri, odgovorni za kompleksne vrste globoka občutljivost: mišično-sklepni, dvodimenzionalno-prostorski čut (definicija številk, črk, številk, narisanih s svinčnikom ali dr. top predmet na človeški koži), občutek teže in volumna gibanja, občutek za prepoznavanje predmetov na dotik.
V spodnjem temenskem režnju so centri za praxis, to so gibi, ki so postali "avtomatski" v procesu ponavljanja in vadbe, ki se razvijajo v procesu učenja in nenehne vadbe, na primer hoja, prehranjevanje, oblačenje itd. tako naprej
Parietalni reženj je vključen v procesiranje in zavestno zaznavanje somatosenzoričnih (iz receptorjev v mišicah, koži, sklepih in notranji organi) informacije, ki vplivajo na prostovoljna gibanja.
Lezije zgornjega parietalnega lobula spremlja razvoj kršitve sposobnosti prepoznavanja predmetov z dotikom z zaprtimi očmi. Bolniki opisujejo posamezne lastnosti predmeta, vendar ne morejo sintetizirati njegove podobe.
S porazom spodnjega parietalnega lobula je moten občutek telesne sheme. Človek se ne more zavedati, kje je desna in kje leva stran, ne prepozna svojih prstov. Druga vrsta motnje je nezavedanje svoje okvare (pacient trdi, da premika paralizirane okončine). Pri teh bolnikih se lahko razvije psevdopolimelija.— občutek dodatnih okončin ali delov telesa. Takšni bolniki lahko samostojno odstranijo "motečo" okončino ali prispevajo k njeni amputaciji.
S poškodbo skorje kotnega gyrusa pacient izgubi občutek za prostorsko dojemanje sveta okoli sebe, položaj lastnega telesa in medsebojne povezave njegovih delov. To spremljajo različni psihopatološki simptomi: depersonalizacija, derealizacija. Opazovati jih je mogoče pod pogojem popolnega ohranjanja zavesti in kritičnega mišljenja.
Z lezijami parietalnega režnja prevladujoče hemisfere oseba razvije disleksijo - nezmožnost branja, razlikovanje med desno in levo, pa tudi diskalkulijo - nezmožnost računanja. Omeniti velja, da je pogosto diskalkulija samostojna bolezen in ne posledica nevroloških ali psihološke težave. Poleg teh težav je možna tudi apraksija - kršitev ali nezmožnost izvajanja nekega namenskega dejanja (dobro, na primer, vzemite kozarec in popijte) ob ohranjanju elementarnih komponent dejanja.
Anastazija Šešukova
Okcipitalni reženj zavzema zadnje dele hemisfer. Na konveksni površini hemisfere okcipitalni reženj nima ostrih meja, ki bi ga ločevale od parietalnega in temporalnega režnja, z izjemo zgornjega dela parietalno-okcipitalnega sulkusa, ki se nahaja na notranji površini hemisfere. ločuje parietalni reženj od okcipitalnega režnja. Brazde in zavoji zgornje stranske površine okcipitalnega režnja so nestabilni in imajo spremenljivo strukturo. Na notranji površini okcipitalnega režnja je žleb, ki ločuje klin (trikotni reženj okcipitalnega režnja) od lingvalnega gyrusa in occipitotemporal gyrusa.
Funkcija okcipitalnega režnja je povezana z zaznavanjem in obdelavo vizualnih informacij, organizacijo kompleksnih procesov vizualne percepcije. V tem primeru je zgornja polovica mrežnice projicirana v območju klina, ki zaznava svetlobo iz spodnjih vidnih polj; v predelu lingularnega gyrusa je spodnja polovica mrežnice, ki zaznava svetlobo iz zgornjih vidnih polj.
Otok
Otoček ali tako imenovana zaprta lobula se nahaja globoko v stranskem žlebu. Otoček je od sosednjih sosednjih delov ločen s krožnim žlebom. Površina otočka je z vzdolžnim osrednjim žlebom razdeljena na sprednji in zadnji del. V otočku je projiciran analizator okusa.
limbični korteks
Na notranji površini hemisfer nad corpus callosum je cingularni gyrus. Ta girus z ožino za corpus callosum prehaja v girus blizu morskega konjička - parahipokampalni girus. Cingulatni girus skupaj s paragitpokampalnim girusom sestavlja obokani girus.
Notranje in spodnje površine hemisfer so združene v tako imenovano limbično (marginalno) skorjo, skupaj z amigdalo iz skupine subkortikalnih jeder, vohalnega trakta in čebulice, območij čelnega, temporalnega in parietalnega režnja možganov. skorjo, pa tudi s hipotuberozno regijo in retikularno tvorbo debla. Limbična skorja je združena v en sam funkcionalni sistem - limbično-retikularni kompleks. Glavna funkcija teh delov možganov ni toliko zagotavljanje komunikacije z zunanjim svetom, temveč uravnavanje tonusa možganske skorje, nagonov in čustvenega življenja. Uravnavajo kompleksne, večplastne funkcije notranjih organov in vedenjske odzive. Limbično-retikularni kompleks je najpomembnejši integrativni sistem telesa. Pri oblikovanju motivacije je pomemben tudi limbični sistem. Motivacija (ali notranja motivacija) vključuje najkompleksnejše instinktivne in čustvene reakcije (prehrambene, obrambne, spolne). Limbični sistem sodeluje tudi pri uravnavanju spanja in budnosti.
Limbična skorja opravlja tudi pomembno funkcijo voha. Vonj je zaznavanje kemikalij v zraku. Vohalni možgani oseba zagotavlja občutek vonja, pa tudi organizacijo kompleksnih oblik čustvenih in vedenjskih reakcij. Vohalni možgani so del limbičnega sistema.
Vohalni možgani so sestavljeni iz dveh delov - perifernega in osrednjega. Periferni del predstavljajo vohalni živec, vohalne čebulice, primarni vohalni centri. Osrednji del vključuje girus morskega konjička - hipokampus, zobat in obokan girus.
Vohalni receptorski aparat se nahaja v nosni sluznici. Preko sistema živčnih prevodnikov se informacije iz receptorjev prenašajo v kortikalni del vohalnega analizatorja (slika 8).
riž. 8. Vohalni analizator (diagram):
1 - vohalni epitelij, bipolarne vohalne celice; 2 - vohalna čebulica; 3 - vohalni trakt; 4 - primarni vohalni centri; 5 - vidni tuberkel; 6 - kortikalni vohalni center; 7 - corpus callosum
Kortikalni del olfaktornega analizatorja se nahaja v cingularnem girusu, girusu morskega konjička in v kavlju morskega konjička, ki skupaj tvorita zaprto obročasto območje. Periferni del olfaktornega analizatorja je povezan s kortikalnimi regijami obeh hemisfer.
Fiziološki mehanizem zaznavanja vonjav z vohalnim analizatorjem ni povsem jasen. Obstajata dve glavni hipotezi, ki pojasnjujeta naravo tega procesa z različnih stališč. Po eni od hipotez se interakcija med molekulami odorantov in kemoreceptorji dogaja kot ključ in ključavnica, tj. vrsta molekule ustreza posebnemu receptorju. Druga hipoteza temelji na predpostavki, da imajo molekule dišeče snovi določen nihajni val, na katerega so "nastavljeni" vohalni receptorji. Molekule, ki imajo podobne vibracije, bi morale imeti skupno valovanje in posledično oddajati podobne vonjave.
Izraz "vohalni možgani" v zvezi s človeško fiziologijo je nekoliko poljuben in ne razkriva v celoti njegove večplastne in univerzalne funkcije. "Umestitev" osrednje vezi vohalnih možganov v možganske hemisfere ni naključna in je posledica ogromne "informacijske" vloge, ki jo je imel občutek za vonj v procesu evolucije pri prilagajanju zunanjemu okolju in uravnavanju kompleksnih vedenjske reakcije. Pridobivanje hrane, izbira osebka nasprotnega spola, skrb za potomce, celovitost ozemlja, organiziranje skupinskih skupnosti znotraj vrste - vse te vsakodnevne funkcije pri mnogih živalih potekajo z neposrednim sodelovanjem fino zgrajenega sistema vohalne recepcije in , ki temelji na tem, sposobnost številnih živali, da pošiljajo subtilne diferencirane specifične dišeče snovi- signali-obveščevalci.
Univerzalne oblike vedenjskih reakcij pri živalih, ki se kažejo v vsakodnevni skrbi za življenjski prostor, za potomce, ustvarjajo vtis, da so obdarjene z razumom. Navidezna inteligenca je preprosto rezultat odzivanja na zunanje dražljaje. Vendar ti dražljaji sami in odzivi nanje popolnoma ustrezajo biološkim potrebam živali.
V življenju ljudi je voh izgubil biološko informacijsko vrednost, ki jo je imel pri živalih. Človeški vohalni sistem je zasnovan tako za opravljanje ozke, "lastne" funkcije kot za nekakšno "polnjenje" čustev. Moč vpliva vonjav na čustveno sfero, da so najpomembnejši »prehranjevalni substrat čustev«, je znana že v pradavnini zgodovine človeštva.
Človekov čut za vonj je lahko različen. Praviloma so te razlike nepomembne, v nekaterih primerih pa je lahko ostrina vonja zelo visoka (degustatorji v parfumski industriji).
Ker ima vohalni analizator pomembno vlogo pri uravnavanju čustev, se njegov osrednji del nanaša na limbični sistem, ki ga figurativno imenujemo "skupni imenovalec" za številne čustvene in viscerosomatske reakcije telesa.
Velike hemisfere možganov
so največji del možganov. Pokrivajo male možgane in možgansko deblo. Možganske hemisfere predstavljajo približno 78% celotne mase možganov.
V procesu ontogenetskega razvoja organizma se iz končnega možganskega mehurja nevralne cevi razvijejo možganske hemisfere, zato ta del možganov imenujemo tudi telencefalon.
Možganske hemisfere delimo na srednja črta globok navpični razpor na desni in leva polobla. V globini srednjega dela sta obe hemisferi povezani z velikim oprijemom - corpus callosum. Vsaka polobla ima režnjeve: čelni, parietalni, časovno, okcipitalni.
Režnji možganskih hemisfer so med seboj ločeni z globokimi brazdami. Najpomembnejši so trije globoki utori: osrednji (Rolandov), ki ločuje čelni reženj od temenskega; stranski (Sylvian), ki ločuje temporalni reženj od parietalnega, in parietalno-okcipitalni, ki ločuje parietalni reženj od okcipitalnega na notranji površini poloble.
Vsaka hemisfera ima zgornjo stransko (konveksno) - konveksilno, spodnjo - bazalno in notranjo - medialno površino. Vsak reženj hemisfere ima možganske vijuge, ki so med seboj ločene z brazdami. Zgoraj je polobla prekrita z lubjem - tanko plastjo sive snovi, ki jo sestavljajo živčne celice.
Korteks- evolucijsko najmlajša tvorba centralnega živčnega sistema. Pri človeku doseže najvišji razvoj. Možganska skorja je zelo pomembna pri uravnavanju vitalne aktivnosti telesa, pri izvajanju kompleksnih oblik vedenja in oblikovanju nevropsihičnih funkcij.
Pod lubjem je belo snov hemisfere, sestavljena je iz procesov živčnih celic - prevodnikov. Zaradi nastajanja možganskih vijug se skupna površina možganske skorje znatno poveča. Skupna površina hemisferne skorje je 1200 cm2, pri čemer se 2/3 njene površine nahaja v globini brazd, 1/3 pa na vidni površini hemisfer. Vsak del možganov ima drugačen funkcionalni pomen.
Korteks je sestavljen iz 4 režnjev, ki so med seboj ločeni z brazdami. Glavni utori, ki ločujejo čelni, parietalni in temporalni reženj, so Rolandov in Sylviev.
Režnji možganske skorje :
čelni (čelni ),
parietalni (parietalni ),
zatilnica (okcipitalni ),
časovno (časovno ),
V zvezi z delnicami govorimo o lokalnih sistemih možganov.
LOKALNI MOŽGANSKI SISTEMI:
Okcipitalni predeli možganov opravljajo funkcije organiziranja vizualne percepcije. Primarne cone okcipitalnega korteksa - elementarne funkcije vizija.
Sekundarni odseki okcipitalnega korteksa - optiko-gnostične funkcije.
Temporalne regije možganov»odgovoren« za organizacijo slušnega zaznavanja. Primarne cone temporalnega korteksa so osnovne funkcije sluha.
Sekundarne cone temporalnega korteksa - akustično-gnostične funkcije.
Senzomotorični in premotorični deli možganov- organizacija gibanja. 1. Postcentralni deli možganov - aferentna organizacija gibov. 2. Cone premotorične skorje - eferentna organizacija (programiranje) gibanja.
3. Motorične cone korteksa - motorični analizator, izvajanje motoričnega programa. Čelni režnji možganov (prefrontalne regije) izvajajo regulacijo duševne dejavnosti, tj. regulacija stanj aktivnosti, prostovoljnih gibov in dejanj, kognitivnih procesov ter čustvene in osebne sfere, so na splošno odgovorni za mišljenje, intelektualno dejavnost. Parietalne regije možganov Predstavljajo jih primarne cone (vse vrste kožno-kinestetične občutljivosti), sekundarne cone (vidno-prostorske predstave, predstave o telesni shemi, somatognoza, stereognoza) in terciarne cone.
Čelni reženj zaseda sprednje dele hemisfer. Od parietalnega režnja je ločen s centralnim sulkusom, od temporalnega režnja pa z lateralnim sulkusom. V čelnem režnju so štirje girusi: en navpični - precentralni in trije vodoravni - zgornji, srednji in spodnji čelni girus.
Zvitki so med seboj ločeni z brazdami. Na spodnji površini čelnih režnjev se razlikujeta neposredni in orbitalni girus. Neposredni girus leži med notranjim robom poloble, vohalnim utorom in zunanjim robom poloble.
V globini vohalne brazde ležita vohalni bulbus in vohalni trakt. Čelni reženj osebe predstavlja 25-28% korteksa, povprečna masa čelnega režnja je 450 g.
Funkcija čelnih režnjev je povezana z organizacijo prostovoljnih gibov, motoričnih mehanizmov govora, regulacijo kompleksnih oblik vedenja in miselnih procesov. Več funkcionalno pomembnih centrov je koncentriranih v vijugah čelnega režnja. Sprednji osrednji girus je "reprezentacija" primarne motorične cone s strogo določeno projekcijo delov telesa. Obraz se "nahaja" v spodnji tretjini gyrusa, roka v srednji tretjini, noga notri zgornja tretjina. Trup je predstavljen v zadnjih delih zgornjega čelnega gyrusa. Tako je oseba projicirana z glavo navzdol in navzdol v sprednjem osrednjem girusu.
Sprednji osrednji girus skupaj s sosednjimi posteriornimi deli čelnih vijug ima funkcionalno zelo pomembno vlogo. Je središče prostovoljnih gibanj. V globinah skorje osrednjega gyrusa iz tako imenovanega piramidalne celice -centralni motorični nevron- začne se glavna motorična pot - piramidna ali kortikospinalna pot. Periferni procesi motoričnih nevronov izhajajo iz skorje, se zberejo v en močan snop, prehajajo skozi osrednjo belo snov hemisfer in vstopijo v možgansko deblo skozi notranjo kapsulo; na koncu možganskega debla se delno križajo (prehajajo z ene strani na drugo) in se nato spustijo v hrbtenjača. Te veje se končajo v sive snovi hrbtenjača. Tam pridejo v stik s perifernim motoričnim nevronom in nanj prenašajo impulze iz centralnega motoričnega nevrona. Impulzi prostovoljnega gibanja se prenašajo po piramidni poti.
V zadnjih delih zgornjega čelnega gyrusa je tudi ekstrapiramidni center skorje, ki je anatomsko in funkcionalno tesno povezan s formacijami tako imenovanega ekstramiramidnega sistema. Ekstrapiramidni sistem- motorični sistem, ki pomaga pri izvajanju poljubnega gibanja. To je sistem "zagotavljanja" poljubnih gibov. Človeški ekstrapiramidni sistem, ki je filogenetsko starejši od piramidnega sistema, zagotavlja avtomatsko regulacijo "naučenih" motoričnih dejanj, vzdrževanje splošnega mišičnega tonusa, "pripravljenost" perifernega motoričnega aparata za gibanje, prerazporeditev mišičnega tonusa med gibanjem. Poleg tega sodeluje pri ohranjanju normalne drže.
V zadnjem delu srednjega čelnega gyrusa je čelni okulomotorni center, ki nadzoruje prijazno, sočasno vrtenje glave in oči (središče vrtenja glave in oči v nasprotni smeri). Draženje tega centra povzroči obračanje glave in oči v nasprotno smer. Funkcija tega centra je zelo pomembna pri izvajanju tako imenovanih orientacijskih refleksov, ki so zelo pomembni za ohranjanje življenja živali.
V zadnjem delu spodnjega čelnega gyrusa je motorično središče govora(Brockovo središče).
Prejema tudi čelna skorja možganskih hemisfer Aktivno sodelovanje pri oblikovanju mišljenja, organizaciji namenskih dejavnosti, dolgoročnem načrtovanju.
temporalni reženj zavzema spodnjo stransko površino hemisfer. Temporalni reženj je ločen od čelnega in parietalnega režnja s stranskim žlebom.
Na zgornji stranski površini temporalnega režnja so trije zavoji - zgornji, povprečje in nižje. Zgornji temporalni girus se nahaja med silvijsko in zgornjo temporalno brazdo, srednji girus je med zgornjim in spodnjim temporalnim sulkusom, spodnji girus pa med spodnjim temporalnim sulkusom in prečno možgansko razpoko. Na spodnji površini temporalnega režnja se razlikujejo spodnji temporalni girus, lateralni okcipitotemporalni girus, hipokampalni girus (noge morskega konjička).
Funkcija temporalnega režnja povezana z zaznavanjem slušnih, okusnih, vohalnih občutkov, analizo in sintezo govornih zvokov, spominskimi mehanizmi. Glavno funkcionalno središče zgornje stranske površine temporalnega režnja se nahaja v zgornjem temporalnem girusu. Tu je slušno ali gnostično središče govora (Wernickejevo središče).
V zgornjem temporalnem girusu in na notranji površini temporalnega režnja je slušno projekcijsko območje korteksa. Vohalno projekcijsko območje se nahaja v hipokampalnem girusu, zlasti v njegovem sprednjem delu (tako imenovani kavelj). Poleg vohalnih projekcijskih con so tudi okusne.
Temporalni režnji igrajo pomembno vlogo pri organizaciji kompleksa miselni procesi, zlasti spomin.
parietalni reženj zavzema zgornje stranske površine poloble. Od čelnega parietalnega režnja, spredaj in ob strani, ga omejuje osrednji sulkus, od temporalnega od spodaj - stranski sulkus, od okcipitalnega - z namišljeno črto, ki poteka od zgornjega roba parietalno-okcipitalnega sulkusa do spodnji rob poloble.
Na zgornji stranski površini parietalnega režnja so trije zavoji: ena navpična - posteriorna osrednja in dve vodoravni - zgornja parietalna in inferiorna parietalna. Del spodnjega parietalnega vijuga, ki ovija posteriorni del lateralnega sulkusa, imenujemo supramarginalni (supramarginalni), del, ki obkroža zgornji temporalni girus, nodalno (kotno) območje.
Parietalni reženj, tako kot čelni reženj, predstavlja pomemben del možganske hemisfere. V filogenetskem smislu se v njem razlikuje stari del - zadnji osrednji girus, nov - zgornji parietalni girus in novejši - spodnji parietalni girus. Funkcija parietalnega režnja je povezana z zaznavanjem in analizo občutljivih dražljajev, orientacijo v prostoru. Več funkcionalnih centrov je koncentriranih v vijugah parietalnega režnja.
V posteriornem osrednjem girusu so centri občutljivosti projicirani s projekcijo telesa, podobno kot v sprednjem osrednjem girusu. V spodnji tretjini gyrusa je projiciran obraz, v srednji tretjini - roka, trup, v zgornji tretjini - noga. V zgornjem parietalnem girusu so centri, ki so odgovorni za kompleksne vrste globoke občutljivosti: mišično-sklepni, dvodimenzionalno-prostorski občutek, občutek teže in volumna gibanja, občutek za prepoznavanje predmetov na dotik.
Tako je kortikalni del občutljivega analizatorja lokaliziran v parietalnem režnju.
Praxis centri se nahajajo v spodnjem parietalnem režnju. Praxis razumemo kot namenske gibe, ki so postali avtomatizirani v procesu ponavljanj in vaj, ki se razvijajo v procesu učenja in nenehne vaje v življenju posameznika.
Hoja, prehranjevanje, oblačenje, strojno pisanje, različne vrste delovna aktivnost (na primer gibi voznika za vožnjo avtomobila, košnja itd.) so praksa.
Praxis- najvišja manifestacija motorične funkcije človeka. Izvaja se kot rezultat skupnih dejavnosti različnih ozemelj. možganska skorja.
Okcipitalni reženj zaseda zadnje predele hemisfer. Na konveksni površini hemisfere okcipitalni reženj nima ostrih meja, ki bi ga ločevale od parietalnega in temporalnega režnja, z izjemo zgornjega dela parietalno-okcipitalnega sulkusa, ki se nahaja na notranji površini hemisfere. ločuje parietalni reženj od okcipitalnega režnja.
Brazde in zavoji zgornje stranske površine okcipitalnega režnja so nestabilni in imajo spremenljivo strukturo.
Na notranji površini okcipitalnega režnja je žleb, ki ločuje klin (trikotni reženj okcipitalnega režnja) od lingvalnega gyrusa in occipitotemporal gyrusa.
Funkcija okcipitalnega režnja je povezana z zaznavanjem in obdelavo vizualne informacije, organizacija kompleksnih procesov vizualne percepcije. V tem primeru je zgornja polovica mrežnice projicirana v območju klina, ki zaznava svetlobo iz spodnjih vidnih polj; v predelu lingularnega gyrusa je spodnja polovica mrežnice, ki zaznava svetlobo iz zgornjih vidnih polj.
Otok, oziroma t.i zaprt lobulus, se nahaja v globini stranske brazde. Otoček je od sosednjih sosednjih delov ločen s krožnim žlebom.
Površina otočka je z vzdolžnim osrednjim žlebom razdeljena na sprednji in zadnji del. Na otoku je predvideno analizator okusa.
Vohalni analizator
Živčne celice, ki zaznavajo draženje vonja, se nahajajo v sluznici zgornjih delov nosne votline. Od tu gredo aksoni teh celic v lobanjsko votlino in vstopijo v vohalne čebulice. Iz njih se pošiljajo živčna vlakna temporalni reženj (notranja površina) kje so živčne celice vohalni analizator.
Analizator okusa
Ta analizator se začne v živčnih končičih brbončic jezika, ki jih v njih predstavljajo brbončice. Živčna vlakna, ki zapuščajo brbončice, gredo v možgane in se končajo, tako kot živci vohalnega analizatorja, pri notranja površina temporalni reženj.
corpus callosum- ločna tanka plošča, filogenetsko mlada, povezuje srednje površine obe polobli. Podolgovati srednji del corpus callosum zadaj prehaja v odebelitev, spredaj pa se ločno ukrivlja in ukrivlja navzdol.
Corpus callosum povezuje filogenetsko najmlajše dele hemisfer in ima pomembno vlogo pri izmenjavi informacij med njimi.
MOŽGANSKO DEBLO ALI MOŽGANSKO DEBLO -
tradicionalno razločen sistem možganskih regij, ki je razširjena tvorba, ki nadaljuje hrbtenjačo.
Možgansko deblo vedno vključuje medullo oblongato, pons in srednje možgane. Pogosto vključuje male možgane, včasih diencefalon.
Medula -
oddelek za možgane. Obstaja tudi tradicionalno ime bulbus (čebulica, zaradi oblike tega oddelka).
Medulla oblongata vstopi v možgansko deblo.
Zunaj, na ventralni (obrazni) strani, so piramide (vsebujejo kortikospinalni trakt - pot od korteksa do motoričnih nevronov hrbtenjače) in olive (vsebujejo jedra spodnje olive, ki so povezana z vzdrževanjem ravnovesja) . Na hrbtni strani: tanki in klinasti snopi, ki se končajo s tuberkulami tankih in klinastih jeder (preklop informacij o globoki občutljivosti spodnje oziroma zgornje polovice telesa), spodnja polovica romboidne jame, ki je dno četrtega ventrikla, in vrvna telesa, ki ga ločujejo, ali spodnji kraki malih možganov.
V notranjosti so tudi jedra od VIII do XII (in eno od jeder VII) kranialnih živcev, del retikularne tvorbe, medialna zanka in druge ascendentne in descendentne poti.
Ima obliko prisekanega stožca.
Zahvaljujoč raziskavam znanstvenikov, kot sta R. Magnus in I. F. Klein, je bilo ugotovljeno, da obstaja kompleksen sistem refleksnih centrov v podolgovati meduli, ki zagotavljajo določen položaj v telesu zaradi statičnih in statično-kinetičnih refleksov. Ti refleksi so pravzaprav mehanizmi za prerazporeditev mišičnega tonusa tako, da se ohrani udobna drža za žival (posturalno-tonični refleksi) ali vrnitev v dano iz neudobne (popravljalni refleksi) in ravnotežje se ohranja tudi med pospeševanjem (stato-kinetični refleksi) . Izvajanje teh refleksov poteka s sodelovanjem takšnih formacij debla, kot so retikularna tvorba, rdeče jedro in vestibularna jedra.
Retikularna tvorba - je tvorba, ki poteka od hrbtenjače do talamusa v rostralni (korteksni) smeri. Retikularna formacija poleg sodelovanja pri procesiranju senzoričnih informacij deluje aktivacijsko na možgansko skorjo in tako nadzoruje delovanje hrbtenjače. Prvič mehanizem delovanja retikularne formacije na mišični tonus je ugotovil R. Granit: pokazal je, da je retikularna formacija sposobna spremeniti aktivnost γ-motoričnih nevronov, zaradi česar ihaksoni (γ-eferenti) povzročajo krčenje mišičnih vreten in posledično povečanje v aferentnih impulzih iz mišičnih receptorjev. Ti impulzi, ki vstopajo v hrbtenjačo, povzročijo vzbujanje α-motoričnih nevronov, kar je vzrok za mišični tonus.
Ugotovljeno je bilo, da pri opravljanju te funkcije retikularne formacije sodelujeta dve skupini nevronov: nevroni retikularne formacije ponsa in nevroni retikularne formacije podolgovate medule. Obnašanje nevronov retikularne formacije medulla oblongata podobno kot se obnašajo nevroni v retikularni formaciji mostu: povzročijo aktivacijo α-motoričnih nevronov mišic upogibalk in s tem zavirajo aktivnost α-motoričnih nevronov mišic iztegovalk. Nevroni retikularne formacije mostu delujejo ravno nasprotno, vzdražijo α-motorične nevrone mišice iztegovalke in zavirajo aktivnost α-motoričnih nevronov mišice upogibalke. Retikularna tvorba je povezana z malimi možgani (del informacij iz nje gre v nevrone medule oblongate (iz jeder plutastih in sferičnih malih možganov) in iz šotora v nevrone mostu) in z možgansko skorjo, iz katere sprejema informacije. To nakazuje, da je retikularna formacija zbiralnik nespecifičnega senzoričnega toka, ki je verjetno vključen v regulacijo mišične aktivnosti.
pomemben funkcionalni pomen retikularno, ali mrežna tvorba možganskega debla, ki se razvije v povezavi s pojavom sistema vagusa, vestibularnega in trigeminalni živci.
Retikularno tvorbo sestavljajo živčne celice različnih velikosti in oblik ter gosta mreža živčna vlakna, ki potekajo v različnih smereh in se nahajajo predvsem v bližini ventrikularnega sistema. Retikularna tvorba ima primarni pomen v kortikalno-subkortikalnih odnosih. Nahaja se v srednjih etažah. medulla oblongata,hipotalamus, siva snov tegmentum, pons.
Retikularni formaciji se približujejo številne kolaterale iz vseh aferentnih (senzoričnih) sistemov. Skozi te kolaterale vsako draženje s periferije, ki se usmeri v določene predele korteksa po specifičnih poteh živčnega sistema, doseže tudi retikularno formacijo. Nespecifični ascendentni sistemi (t.j. poti iz retikularne tvorbe) zagotavljajo vzbujanje možganske skorje, aktiviranje njegove aktivnosti.
Skupaj z naraščajočimi nespecifičnimi sistemi skozi možgansko deblo prehajajo padajoči nespecifični sistemi, ki vplivajo na spinalne refleksne mehanizme.
Retikularna tvorba je tesno povezana z limbičnim sistemom, pa tudi z možgansko skorjo. Zaradi tega se tvori funkcionalna povezava med višjimi deli centralnega živčnega sistema in deblom. možgani. Ta sistem se imenuje limbično-retikularni kompleks ali limbično-retikularna os. Ta kompleksen strukturni in funkcionalni kompleks zagotavlja integracijo najpomembnejših funkcij, pri izvajanju katerih sodelujejo različni deli možganov.
Znano je, da budnost korteksa zagotavljajo specifični in nespecifični sistemi. Aktivacijska reakcija se vzdržuje s stalnim dovajanjem impulzov iz receptorjev slušni, vizualni, olfactory, gustatory in kožno-kinestetični analizatorji. Ti dražljaji se prenašajo po specifičnih aferentnih poteh v različne dele korteksa. Od vseh prijavljenih talamus, nato pa številni kolaterali retikularne formacije odidejo v skorjo možganskih hemisfer aferentnih poti, kar zagotavlja njegovo naraščajočo aktivacijsko aktivnost.
Po drugi strani pa retikularna tvorba prejema impulze iz mali možgani, subkortikalna jedra, limbični sistem ki zagotavljajo čustveno prilagodljive vedenjske odzive, motivacijske oblike vedenja. Vendar pa je stopnja zagotavljanja adaptivnih brezpogojnih refleksnih reakcij s strani nespecifičnega sistema pri ljudeh in živalih različna. Če imajo pri živalih podkortikalne tvorbe in limbični sistem vodilno vlogo pri zadovoljevanju vitalnih potreb organizma za njegovo preživetje, okolju, potem je pri človeku zaradi prevlade skorje aktivnost globokih struktur možganov (subkortikalnih tvorb, limbičnega sistema, retikularne tvorbe) v večji meri kot pri živalih podrejena možganska skorja. Retikularna formacija ima pomembno vlogo pri uravnavanju mišičnega tonusa. Mišični tonus uravnavata dve vrsti retikulospinalnih poti. Hitro prevodni retikulospinalni trakt uravnava hitre gibe; počasi vodena retikulospinalna pot - počasni tonični gibi.
Retikularna formacija podolgovate medule je vpletena v pojav decerebratne rigidnosti. Pri prerezu možganskega debla nad podolgovato medullo se zmanjša aktivnost nevronov, ki imajo zaviralni učinek na motorične nevrone hrbtenjače, kar povzroči močno povečanje tonusa skeletnih mišic.
Funkcije podolgovate medule
Zaščitni refleksi (na primer kašljanje, kihanje).
Vitalni refleksi (npr. dihanje).
uravnavanje žilnega tonusa.
Refleksni centri podolgovate medule:
zaščitno (kašljanje, kihanje itd.)
centri za nadzor tona skeletna mišica ohraniti držo osebe.
skrajšanje ali podaljšanje časa spinalnih refleksov
prebavo
srčna aktivnost
Pons
Pons Varolii (po imenu Constanzo Varolia), ali most - del možganov, je skupaj z malimi možgani del zadnjih možganov. pripada možganom,
V osnovi mostu so padajoče poti: kortikospinalni piramidalni trakt, kortikobulbarni, kortikomostni trakt.
srednji možgani(lat.Mesencephalon) - del možganov, starodavno vidno središče. Vključeno v možgansko deblo.
funkcije srednjih možganov
1. motor,
2. čutila (vid, sluh),
3. Uravnavanje dejanj žvečenja in požiranja,

