Velike hemisfere velike poloble
možgani, parne tvorbe, ki jih corpus callosum združuje v tako imenovani telencefalon. Površina hemisfere ki ga predstavljajo številni veliki ali majhni globoki zavoji. Obstajajo režnji: čelni, parietalni, temporalni, otočni, okcipitalni. Siva snov možganov je sestavljena iz živčne celice- nevroni, tvorijo možgansko skorjo in subkortikalne ganglije (vozlišča). belo snov ki ga tvorijo procesi nevronov, ki tvorijo možganske poti.
VELIKE POLOKRLEVELIKE HEMISFERE možganov, parne tvorbe, ki jih združuje corpus callosum (cm. corpus callosum) v tako imenovani. terminalni možgani. Površino možganskih hemisfer predstavljajo številni veliki ali majhni globoki vijugi. Obstajajo režnji: čelni, parietalni, temporalni, otočni, okcipitalni. Siva snov možganov, sestavljena iz živčnih celic - nevronov, tvori možgansko skorjo in subkortikalne ganglije. (cm. GANGLION)(vozlišča). Belo snov tvorijo procesi nevronov, ki sestavljajo možganske poti.
enciklopedični slovar. 2009 .
Oglejte si, kaj so "velike hemisfere" v drugih slovarjih:
Možgani so seznanjene tvorbe, ki jih corpus callosum združuje v tako imenovani. terminalni možgani. Površino možganskih hemisfer predstavljajo številni veliki ali majhni globoki vijugi. Obstajajo režnji: čelni, parietalni, temporalni, otočni ... Veliki enciklopedični slovar
Možgani, seznanjene tvorbe, združene s corpus callosum v ti. terminalni možgani. Površino B. p. predstavljajo številni. b. ali m.globoke vijuge. Obstajajo režnji: čelni, parietalni, temporalni, otočni, okcipitalni. Siva v ...... Naravoslovje. enciklopedični slovar
VELIKE MOŽGANSKE POLOBE- višji deli možganov, sestavljeni iz površinske plasti možganske skorje in globokih delov podkorteksa; pokrivajo male možgane in možgansko deblo. B. p. g. m. deljeno s srednja črta na desni in levi hemisferi, ki v globini ... Psihomotorika: Referenca slovarja
Arktični ocean, v nasprotju z južnim, predstavlja povsem sredozemski značaj. Ima naravne meje na precejšnji razdalji in se le na treh mestih neposredno zlije z vodami Atlantika in Pacifika ... ...
Arktični ocean je v nasprotju z južnim povsem sredozemskega značaja. Ima naravne meje na precejšnji razdalji in se le na treh mestih neposredno zlije z vodami Atlantika in Pacifika ... ... Enciklopedični slovar F.A. Brockhaus in I.A. Efron
Imenuje se tudi primerjalna morfologija in preučuje vzorce strukture in razvoja organov s primerjavo različne vrsteŽiva bitja. Podatki primerjalne anatomije so tradicionalna osnova biološke klasifikacije. Pod morfologijo... Enciklopedija Collier
Prerez možganov odraslega moškega. Človeški možgani (latinsko encephalon) so približno ... Wikipedia
Veda, ki preučuje strukturo telesa posameznih teles, tkiva in njihova razmerja v telesu. Za vsa živa bitja so značilne štiri lastnosti: rast, metabolizem, razdražljivost in sposobnost samorazmnoževanja. Kombinacija teh znakov ... ... Enciklopedija Collier
Živali (Mammalia), razred vretenčarjev, največ znana skupinaživali, vključno z več kot 4600 vrstami svetovne favne. Vključuje mačke, pse, krave, slone, miši, kite, ljudi itd. V procesu evolucije so sesalci izvajali najširše ... ... Enciklopedija Collier
I Medicina Medicinski sistem znanstvena spoznanja in praktične dejavnosti za krepitev in ohranjanje zdravja, podaljševanje življenja ljudi, preprečevanje in zdravljenje bolezni ljudi. Za izpolnitev teh nalog M. preučuje strukturo in ... ... Medicinska enciklopedija
Človeški možgani so izjemno kompleksen sistem. Zahvaljujoč temu organu so ljudje dosegli stopnjo razvoja, ki jo opazimo zdaj. Kaj on predstavlja?
evolucijski razvoj
V sodobnem šolskem tečaju biologije se teme obravnavajo od preprostih do zapletenih. Najprej pogovarjamo se o celicah, praživalih, bakterijah, rastlinah, glivah. Kasneje pride do prehoda na živali in človeka. Do neke mere to odraža hipotetični potek evolucije. Če upoštevamo strukturo, na primer, črvov, je enostavno videti, da je veliko preprostejša kot pri ljudeh ali višjih živalih. Toda ti organizmi imajo nekaj pomembnega - ganglij, ki opravlja funkcije možganov.
prednji možgani
Če nekoga prosite, naj nariše vsebino človeške lobanje, bodo poloble najverjetneje shematično prikazane. To je res eden najbolj vidnih in največjih delov. Toda sprednji možgani vsebujejo tudi medullo oblongato. Na splošno je njihova struktura precej zapletena. In če upoštevamo podrobnejšo delitev, potem lahko celo poimenujemo vse oddelke prednji možgani:
- hipokampus;
- bazalni gangliji;
- veliki možgani.
Seveda obstaja še podrobnejša delitev, ki pa praviloma zanima le strokovnjake. No, za tiste, ki si šele širijo obzorja, bo veliko bolj zabavno izvedeti, kaj vse ti oddelki počnejo. Kakšne so torej funkcije prednjih možganov? In zakaj obstajajo razlike med mišljenjem desničarjev in levičarjev?
Funkcije
Sprednji možgani vključujejo nazadnje razvite dele. In to pomeni, da ima oseba zaradi njih lastnosti, ki jih ima. In če se diencefalon v glavnem ukvarja z uravnavanjem metabolizma, primitivnih refleksov in potreb, pa tudi preprostih motorična aktivnost, potem so hemisfere tisti kraj, kjer se rojevajo zavestne misli, kjer se informacije učijo in shranjujejo ter ustvarjajo nekaj novega.
Hemisfere so tudi pogojno razdeljene na več delov-območij: parietalno, čelno, posteriorno in temporalno. In tukaj so celice, ki se med drugim ukvarjajo z analizo informacij, ki prihajajo od zunaj: vizualni, slušni, vohalni, okusni in taktilni centri.
Najbolj zanimivo je, da se s funkcionalnega vidika leva in desna polobla razlikujeta. Seveda obstajajo primeri, ko je ob poškodbi enega dela možganov njegove naloge prevzel drugi, to pomeni, da obstaja določena zamenljivost, vendar je v običajnem primeru lahko situacija naslednja: leva polobla se ukvarja z analizo intonacije govora druge osebe, desna pa z razlago pomena povedanega. Zato levičarji in desničarji, ki imajo bolj razvite različne dele, razmišljajo nekoliko drugače.
Tudi funkcije prednjih možganov vključujejo spomin, različne reakcije na zunanje dražljaje, načrtovanje in gradnjo prihodnjih scenarijev in situacij. Tu je tudi govorni center. Tu poteka vsa višja živčna dejavnost: ustvarjalnost, razmišljanja, ideje.
Zanimivo je tudi, da se prednji del možganov aktivno razvija ne samo v predporodnem obdobju, ampak tudi v prvih nekaj letih življenja. Vsaka nova veščina in spretnost, naučena beseda, katera koli pomembna informacija- vse to tvori novo nevronske povezave. In takšna karta je unikatna za vsako osebo.

- Miselne sposobnosti niso odvisne od mase možganov, ampak so v korelaciji s takšno vrednostjo, kot je število konvolucij.
- Hitrost signalov med nevroni doseže 288 kilometrov na uro. S starostjo se ta številka zmanjšuje.
- Možgani porabijo največ energije med človeški organi- približno 20%. To je ogromna številka, glede na to, da je njegova masa glede na telo le 2%. Prav tako je za njegovo normalno delovanje potrebna zadostna količina tekočine v telesu.
- Trditev, da možgani porabijo le 10% svojih virov, je mit. Res ne more delovati toliko centrov hkrati, a tako ali drugače so vključeni vsi.
besedilna_polja
besedilna_polja
puščica_navzgor
Diencephalon skupaj z možganskim deblom je pokrit od zgoraj in s strani velike poloble - terminalni možgani. Hemisfere so sestavljene iz subkortikalnih vozlov (bazalnih ganglijev) in imajo votline -. Zunaj so poloble pokrite (s plaščem).
Bazalni gangliji ali bazalni gangliji
besedilna_polja
besedilna_polja
puščica_navzgor
Bazalni gangliji oz subkortikalni vozli, (nuclei basales)- tvorbe so filogenetsko starejše od skorje. Bazalni gangliji so dobili ime po tem, da ležijo tako rekoč na dnu možganskih hemisfer, v njihovem bazalnem delu. Sem spadajo kavdatno in lentikularno jedro, združeno v striatum, ograjo in amigdalo.
Repno jedro
besedilna_polja
besedilna_polja
puščica_navzgor
Repno jedro (nucleus caudatus) podolgovat v sagitalni ravnini in močno ukrivljen (sl. 3.22; 3.32; 3.33). Njegov sprednji, odebeljeni del - glavo- se nahaja pred talamusom, v stranski steni sprednjega roga stranski ventrikel, od zadaj se postopoma oži in preide v rep. Repno jedro pokriva vidni tuberkel spredaj, od zgoraj in s strani.
riž. 3.22.
1 - kaudatno jedro;
2 - stebri loka;
3 - epifiza;
4 - vrh in
5 - spodnji kolikul;
6 - vlakna srednjega cerebelarnega peclja;
7 - pot zgornjega cerebelarnega peclja (razrezana);
8 - jedro šotora;
9 - črv;
10 - sferično,
11 - pluta in
13 - zobno jedro;
12 - skorja hemisfer malih možganov;
14 - zgornji cerebelarni pecelj;
15 - trikotnik povodca;
16 - vzglavnik talamusa;
17 - vidni tuberkel;
18 - posteriorna komisura;
19 - tretji ventrikel;
20 - sprednje jedro vidnega hriba
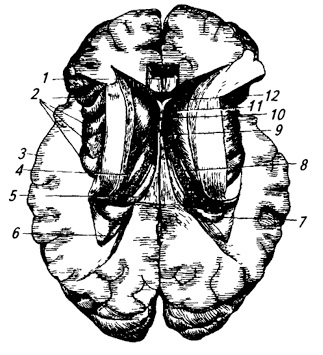 riž. 3.32.
riž. 3.32. riž. 3.32. Možgani - vodoravni prerez skozi stranske ventrikle:
1 - corpus callosum;
2 - otoček;
3 - lubje;
4 - rep kavdatnega jedra;
5 - lok;
6 - zadnji rog lateralnega ventrikla;
7 - hipokampus;
8 – horoidni pleksus;
9 - interventrikularna odprtina;
10 - prozorna pregrada;
11 - glava repnega jedra;
12 – sprednji rog stranski ventrikel
Jedro v obliki leče
besedilna_polja
besedilna_polja
puščica_navzgor
Jedro v obliki leče (nucleus lentiformis) ki se nahaja zunaj vizualne gomile, na ravni otoka. Oblika jedra je podobna trikotni piramidi, z osnovo obrnjeno navzven. Jedro je jasno razdeljeno s plastmi bele snovi na temneje obarvan stranski del - lupina in medialno bleda žoga, sestavljen iz dveh segmentov: notranjega in zunanjega (sl. 3.33; 3.34).
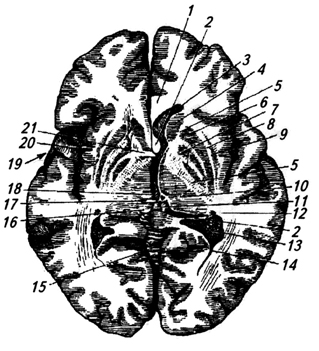 riž. 3.33.
riž. 3.33. riž. 3.33. Horizontalni prerez možganskih hemisfer na ravni bazalnih ganglijev:
1 - corpus callosum;
2 - trezor;
3 - sprednji rog lateralnega ventrikla;
4 - glava repnega jedra;
5 - notranja kapsula;
6 - lupina;
7 - bleda žoga;
8 - zunanja kapsula;
9 - ograja;
10 - talamus;
11 - epifiza;
12 - rep kavdatnega jedra;
13 - horoidni pleksus stranskega prekata;
14 - zadnji rog lateralnega ventrikla;
15 - cerebelarni vermis;
16 - kvadrigemina;
17 - posteriorna komisura;
18 - votlina tretjega prekata;
19 - jama stranske brazde;
20 - otoček;
21 - sprednja komisura
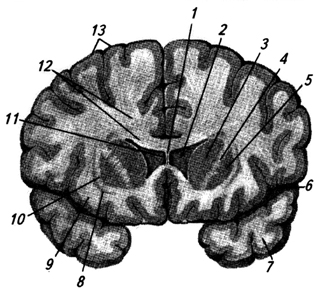 riž. 3.34.
riž. 3.34. riž. 3.34. Frontalni prerez skozi možganske hemisfere na ravni bazalnih ganglijev:
1 - corpus callosum;
2 - stranski prekat;
3 - kavdatno jedro (glava);
4 - notranja kapsula;
5 - jedro lečaste oblike;
6 - stranska brazda;
7 - temporalni reženj;
8 - ograja;
9 - otoček;
10 - zunanja kapsula;
11 - prozorna pregrada;
12 - sijaj corpus callosum;
13 - možganska skorja
školjka
besedilna_polja
besedilna_polja
puščica_navzgor
 riž. 3.35.
riž. 3.35. školjka (putamen) genetsko, strukturno in funkcionalno blizu repnemu jedru.
Obe formaciji imata bolj zapleteno strukturo kot bleda žoga. Do njih pridejo vlakna predvsem iz možganske skorje in talamusa (slika 3.35).
riž. 3.35. Aferentne in eferentne povezave bazalnih ganglijev:
1 - precentralni girus;
2 - lupina;
3 - zunanji in notranji segmenti blede kroglice;
4 - lečasta zanka;
5 - retikularna tvorba;
6 - retikulospinalni trakt,
7 - rubrospinalni trakt;
8 - cerebelarno-talamični trakt (iz dentatnega jedra malih možganov);
9 - rdeče jedro;
10 - črna snov;
11 - subtalamično jedro;
12 - Zona incerta;
13 - hipotalamus;
14 - ventrolateralno,
15 - intralaminarna in centromediana jedra talamusa;
16 - III prekat;
17 - kavdatno jedro
bleda žoga
besedilna_polja
besedilna_polja
puščica_navzgor
Globus pallidus (globus pallidus) je v glavnem povezan s prevajanjem impulzov po številnih padajočih poteh do spodnjih možganskih struktur - rdečega jedra, črne snovi itd. Vlakna iz nevronov pallidusa gredo v ista talamična jedra, ki so povezana z malimi možgani. Iz teh jeder vodijo številne poti v možgansko skorjo.
Bleda kroglica sprejema impulze iz kavdatnega jedra in putamena.
striatum ( corpus striatum) (striatum), ki združuje kavdatno in lentikularno jedro, spada med eferentne ekstrapiramidni sistem. Dendriti striatnih nevronov so prekriti s številnimi bodicami. Zaključujejo vlakna iz nevronov skorje, talamusa in substancije nigre (slika 3.35). Po drugi strani striatni nevroni pošiljajo aksone v intralaminarna, sprednja in lateralna jedra talamusa. Od njih gredo vlakna v skorjo in tako se sklene povratna zanka med kortikalnimi nevroni in striatumom.
V procesu filogeneze so bila ta jedra zgrajena nad jedri srednjih možganov. S prejemanjem impulzov iz talamusa striatum sodeluje pri izvajanju tako zapletenih avtomatskih gibov, kot so hoja, plezanje, tek. V jedrih striatuma so zaprti loki najkompleksnejših brezpogojnih, tj. prirojeni, refleksi. Ekstrapiramidni sistem je filogenetsko starejši od piramidnega sistema. Pri novorojenčku le-ta še ni dovolj razvita in se impulzi dovajajo mišicam iz subkortikalnih ganglijev preko ekstrapiramidnega sistema. Zaradi tega je za gibanje otroka v prvih mesecih življenja značilna generalizacija, nediferenciacija. Z razvojem možganske skorje aksoni njihovih celic rastejo do bazalnih ganglijev in aktivnost slednjih začne uravnavati skorja. Subkortikalni gangliji niso povezani le z motoričnimi reakcijami, ampak tudi z vegetativne funkcije- te so najvišje subkortikalni centri avtonomni živčni sistem.
amigdala
besedilna_polja
besedilna_polja
puščica_navzgor
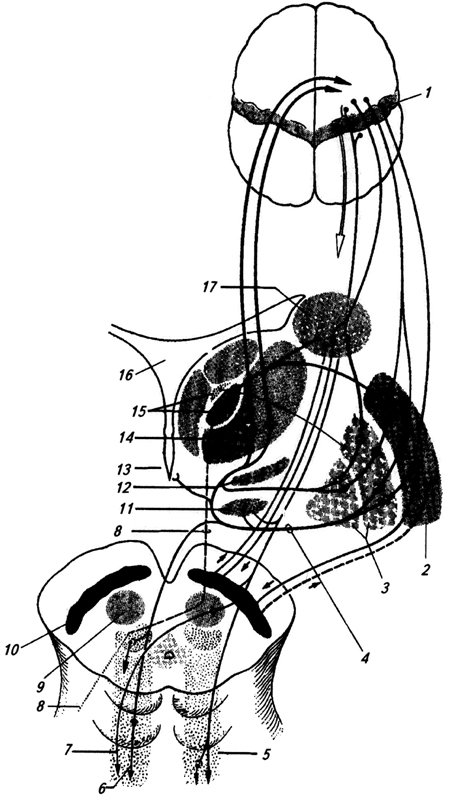
amigdala (corpus amygdaloideum) (amigdala) - kopičenje celic v beli snovi temporalnega režnja. S pomočjo sprednja komisura povezuje se z istoimenskim telesom na drugi strani. Amigdala sprejema impulze iz različnih aferentnih sistemov, vključno z vohalnim sistemom, in je povezana s čustvenimi reakcijami (slika 3.36).
 riž. 3.36.
riž. 3.36. riž. 3.36. Možganske strukture, povezane z amigdalo: aferentne (A) in eferentne (B) povezave amigdale:
1 - jedra talamusa;
2 - periaqueductal siva snov;
3 - parabrahialno jedro;
4 - modra točka;
5 - šivalna jedra;
6 - jedro ene poti;
7 - dozalno jedro X živca;
8 - temporalna skorja;
9 - vohalna skorja;
10 - vohalna žarnica;
11 - čelna skorja;
12 - cingularni girus;
13 - corpus callosum;
14 - vohalno jedro;
15 - anteriorno-ventralno in
16 - dorsomedialna jedra talamusa;
17 - osrednji,
18 - kortikalna in
19 - bazolateralno jedro amigdale;
20 - hipotalamus;
21 - retikularna tvorba;
22 - pregrada;
23 - črna snov;
24 - ventromedialno jedro hipotalamusa; XXIII, XXIV, XXVIII - kortikalna polja
Možganske hemisfere so največje regije možganov. Človeška polobla veliki možgani so bili v primerjavi z ostalimi deli maksimalno razviti, kar v veliki meri razlikuje možgane človeka in živali. levo in desna polobla možganov so ločeni drug od drugega z vzdolžno razpoko, ki poteka vzdolž srednje črte. Če pogledate površino možganov od zgoraj in od strani, lahko vidite reži podobno vdolbino, ki se začne 1 cm posteriorno od srednje točke med sprednjim in zadnjim polom možganov in gre globlje. To je osrednja (Rolandova) brazda. Pod njim vzdolž stranske površine možganov prehaja drugi veliki stranski (Sylvian) žleb. Funkcije hemisfera prednji možgani - tema članka.
1 106654
Fotogalerija: Funkcije možganske hemisfere prednjih možganov
Možganski režnji
Velike hemisfere so razdeljene na režnje, katerih imena so podana po kosteh, ki jih pokrivajo: čelni režnji se nahajajo pred Rolandovim in nad Silvijevim žlebom.
Parietalni reženj leži za osrednjim in nad zadnjim delom stranskega sulkusa; sega nazaj do parieto-okcipitalnega sulkusa – vrzel, ki ločuje parietalni reženj iz zatilnice, ki tvori zadnji del možganov.
Temporalni reženj je območje pod Silvijevim sulkusom in meji na okcipitalni reženj.
Ker možgani hitro rastejo že pred rojstvom, začne možganska skorja povečevati svojo površino, tvoriti gube, kar povzroči nastanek značilnega videz možganom podobna oreh. Te gube so znane kot vijuge, utori, ki jih ločujejo, pa se imenujejo brazde. Nekateri utori se pri vseh ljudeh nahajajo na istem mestu, zato se uporabljajo kot mejniki za razdelitev možganov na štiri režnje.
Razvoj konvolucij in brazd
Brazde in vijuge se začnejo pojavljati v 3-4 mesecu razvoja ploda. Do te točke ostane površina možganov gladka, kot možgani ptic ali dvoživk. Tvorba zložene strukture zagotavlja povečanje površine možganske skorje v omejenem volumnu lobanje. Različna področja možganske skorje opravljajo določene, visoko specializirane funkcije. Možgansko skorjo lahko razdelimo na naslednja področja:
Motorične cone - sprožijo in nadzorujejo gibe telesa. Primarno motorično področje nadzoruje prostovoljne gibe na nasprotni strani telesa. Neposredno pred motoričnim območjem skorje je tako imenovana premotorična skorja, tretje področje - dodatno motorično območje - pa leži na notranji površini čelnega režnja.
Senzorični predeli možganske skorje zaznavajo in povzemajo informacije iz senzoričnih receptorjev po celem telesu. Primarno somatosenzorično področje sprejema informacije z nasprotne strani telesa v obliki impulzov iz senzoričnih receptorjev za dotik, bolečino, temperaturo ter položaj sklepov in mišic (proprioceptivni receptorji).
Površina človeškega telesa ima svoje »predstave« v senzoričnih in motoričnih predelih možganske skorje, ki so organizirani na določen način. Kanadski nevrokirurg Wilder Penfield, ki je deloval v petdesetih letih prejšnjega stoletja, je ustvaril nekakšen zemljevid senzorične cone možganske skorje, ki zaznava informacije iz različnih delov telesa. Je v okviru svojih raziskav izvajal poskuse, v katerih je osebi pod lokalna anestezija opišite svoje občutke v trenutku, ko je stimuliral določena področja površino možganov. Penfield je ugotovil, da stimulacija postcentralnega girusa povzroča taktilne občutke na določenih področjih na nasprotni strani telesa. Druge študije so pokazale, da je obseg motoričnega korteksa, ki je odgovoren za različne dele človeškega telesa, bolj kot od moči in volumna odvisen od stopnje zahtevnosti in natančnosti izvedenih gibov. mišična masa. Možgansko skorjo sestavljata dve glavni plasti: siva snov - tanka plast živčnih in glialnih celic debeline približno 2-A mm in bela snov, ki jo tvorijo živčna vlakna (aksoni) in glialne celice.
Površina možganskih hemisfer je prekrita s plastjo sive snovi, katerih debelina se v različnih delih možganov giblje od 2 do 4 mm. Sivo snov tvorijo telesa živčnih celic (nevroni) in glialne celice, ki opravljajo podporno funkcijo. V večjem delu možganske skorje je pod mikroskopom mogoče najti šest ločenih plasti celic.
Nevroni možganske skorje
- Piramidalne celice so dobile ime zaradi oblike telesa nevrona, ki spominja na piramido; njihovi aksoni (živčna vlakna) izhajajo iz možganske skorje in prenašajo informacije v druge dele možganov.
- Nepiramidalne celice (vse ostale) so zasnovane za zaznavanje in obdelavo informacij iz drugih virov.
Debelina šestih plasti celic, ki sestavljajo možgansko skorjo, se močno razlikuje glede na področje možganov. Nemški nevrolog Korbinian Brodmann (1868-191) je raziskoval te razlike tako, da je obarval živčne celice in jih pregledal pod mikroskopom. Rezultat Brodmannovega znanstvenega raziskovanja je bila razdelitev možganske skorje na 50 ločenih delov na podlagi določenih anatomskih kriterijev. Poznejše študije so pokazale, da imajo tako identificirana "Brodmannova polja" posebno vlogo. fiziološka vloga in imajo edinstvene načine interakcije.
Možgani (nadaljevanje)
Največjo velikost in kompleksnost pri sesalcih dosežejo sprednji ali končni možgani (telencephalon), sestavljeni iz dveh možganskih hemisfer (hemispheri cerebri). Očitno so hemisfere nastale predvsem (in morda izključno) v povezavi z vohalno recepcijo. Vonjave v življenju višjih primatov, vključno s človekom, ne pomenijo toliko. Vendar pa za več zgodnje faze V evoluciji je bil do prednikov vretenčarjev voh glavni kanal, po katerem so živali prejemale informacije o svetu okoli sebe. Zato je povsem naravno, da so vohalni centri v možganih služili kot osnova, na kateri so se kasneje razvile kompleksne strukture. živčni mehanizmi. Že v zgodnjih fazah evolucije tetrapodov se hemisfere spremenijo v velika in pomembna središča korelacije senzoričnih signalov. V času, ko so se pojavili sesalci, je močno razširjena površina hemisfer postala prevladujoč asociativni center, kraj lokalizacije višje duševne dejavnosti. Pri različnih predstavnikih razreda se razmerje med maso hemisfer prednjih možganov in maso celotnih možganov razlikuje: pri ježu ( Erinaceus europaeus) je 48%, za beljakovine ( Sciurus vulgaris) - 53%, volk ( canis lupus) - 70%, v skupnem boku ( Delphinus delphis) - 75%, pri večini primatov - 75-80%, pri ljudeh - približno 85%. Pri pticah velike hemisfere po masi približno ustrezajo preostalim možganom ali so manjvredne od njih, včasih večkrat. Nenazadnje o izjemnem pomenu možganskih hemisfer priča dejstvo, da njihovo uničenje povzroči popolno funkcionalno odpoved sesalca.
Od spodaj se vohalne čebulice (bulbi olfactorii) prilegajo sprednjemu delu hemisfer. Te tvorbe so najbolj razvite pri živalih z dobrim vohom in so močno zmanjšane pri čisto vodnih oblikah. Glede na različno razvitost ločimo več vrst zgradbe možganov. Pri vrečarjih, žužkojedih, brezzobcih, mesojedih, glodalcih in nekaterih drugih so vohalne čebulice velike in dobro štrlijo, če gledamo možgane od zgoraj. Ta tip možganov s popolnim razvojem vohalnih režnjev se imenuje makromatični. Pri plavutonožcih, sirenah in mnogih primatih so čebulice slabo razvite; Te živali imajo mikrosmatične možgane. Nazadnje, za kite, tako imenovane. anosmatski možgani z zmanjšanimi vohalnimi žarnicami. Včasih je veljalo, da je sposobnost razlikovanja med kemičnimi signali pri kitih in delfinih popolnoma izgubljena, vendar se je izkazalo, da to ne drži povsem.
Površinske plasti hemisfer prednjih možganov sesalcev tvorijo palij ali možganski forniks (palij). Zgornja plast, sestavljena iz teles nevronov in ne-mesnatega živčna vlakna, se imenuje skorja (cortex cerebri) in je siva snov loka. Telesa nevronov se nahajajo v skorji v plasteh in tvorijo nekakšne zaslonske strukture. Ta organizacija možganov vam omogoča prostorsko prikazovanje zunanjega sveta na podlagi informacij, ki prihajajo iz čutil. Zaslonske strukture so značilne za najpomembnejše možganske centre sesalcev, medtem ko so pri drugih vretenčarjih manj pogoste, predvsem v centrih za vid. Pod skorjo je plast mieliniziranih procesov nevronov - bela snov forniksa. Živčna vlakna bele snovi tvorijo prevodne snope, ki spajkajo hemisfere - možganske komisure. Čeprav je glavnina sive snovi skoncentrirana v skorji, je prisotna tudi v subkortikalni plasti, kjer je predstavljena z razmeroma majhnimi skupki – jedri. Sem sodijo progasta telesca (corpora striata), ki ležijo pod votlino stranskega ventrikla in so dobila ime zaradi živčnih vlaken, ki jih prečkajo. Pod nadzorom korteksa striatum opravlja funkcijo uravnavanja stereotipnih, avtomatiziranih reakcij - brezpogojnih refleksov.
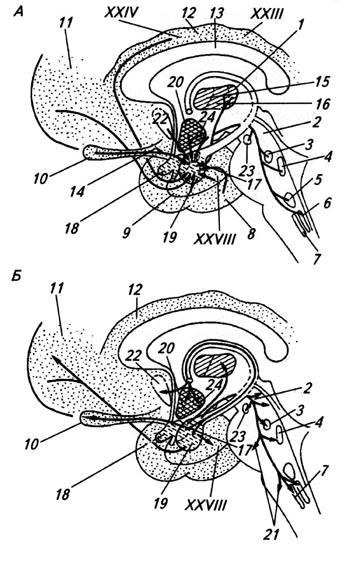
Sheme, ki odražajo progresivni razvoj hemisfer prednjih možganov. Stranski pogled na hemisfero z olfaktornim bulbusom. Različni oddelki, ki se med seboj citološko razlikujejo, se različno obarvajo.
A - hemisfera je samo vohalni reženj.
B - diferenciran je dorzalni del - arhipalij (= hipokampus) in ventralni del - bazalno jedro (striatum).
B - bazalna jedra so se preselila v notranjo regijo poloble.
G - pojavi se majhno območje, ki je neopalij.
E - paleopalij je potisnjen nazaj na medialno površino hemisfere, vendar je neopalij še vedno skromne velikosti, pod vohalnim žlebom pa ostanejo močno razvite vohalne regije.
E - primitivni olfaktorni predel je ohranjen le v ventralnem predelu, neopalij pa doseže izjemno močan razvoj. (Po Romer in Parsons, 1992.) Forniks možganov je začel nastajati že pri ribah. Zaradi progresivnega razvoja vonja senzorični sistem imajo paleopallium ali starodavni obok (paleopallium), ki popolnoma pokriva majhne poloble. Na stopnji rib z režnjami se v hrbtnem delu hemisfer, bližje telesni osi, pojavi arhipalij ali stari obok (archipallium). Pri dvoživkah in primitivnih plazilcih se še naprej razvija, zaradi česar je paleopalij potisnjen vstran in se ohrani le vzdolž stranske površine hemisfer. Hkrati ima paleopallium še naprej pretežno vohalni značaj in na najvišjih stopnjah evolucije tvori vohalne režnje hemisferične skorje. Archipallium je v določeni, čeprav majhni meri, korelativni center, ki prejema vzpenjajoča se vlakna iz diencefalona, pa tudi vlakna iz olfaktornega bulbusa in olfaktornega režnja; zdi se, da je povezano tudi s čustvenim vedenjem. Živčna pot od tega področja do hipotalamusa je glavni element snopa vlaken, ki se pri sesalcih imenuje forniks.
Pri dvoživkah se prvič pojavi zametek neopalija ali nov obok (neopallium). Pri plazilcih tvori neopalij že majhno območje med starim in starim obokom. To področje je že od vsega začetka svojega razvoja asociativno središče, ki tako kot bazalna jedra sprejema vlakna, ki preklapljajo senzorične signale iz možganskega debla nanj in, nasprotno, posredujejo ukaze neposredno motoričnim stebrom.
Pri monotremih je neopalij še vedno zagozden med paleopalij na zunanji strani in arhipalij na notranji strani. Pri vrečarjih raste preko strehe in stranskih sten hemisfer. Istočasno je arhipalij iztisnjen na medialno površino, paleopalij pa je omejen na ventrolateralni del hemisfere, ki se nahaja pod nosnim žlebom (fissura rhinalis) - žleb, ki predstavlja mejo med vohalnim in nevohalnim področjem korteksa. V placenti zaradi nadaljnjega zapletanja in širjenja neopalija hemisfere dosežejo takšno velikost, da presegajo volumen ostalih možganov skupaj. Hemisfere rastejo nazaj in na straneh, zaporedoma pokrivajo diencefalon, srednji možgani in del malih možganov. Parni ventrikli in starodavne strukture, ki so odgovorne predvsem za voh (vohalne žarnice, stare plasti forniksa in z njimi povezani živčni snopi in jedra), so v možganih posteljice potisnjeni vstran in deformirani. Tako je paleopalium ohranjen na ventralni površini hemisfere v obliki majhnega vonjalnega področja, imenovanega hruškasti reženj (lobus piriformis), arhipalij pa je v globini gube temporalnega režnja zvit v zvitek, imenovan hipokampus. Premoč možganskih hemisfer nad ostalimi deli možganov je opazna pri vseh sesalcih, še posebej pa je izrazita pri naprednejših oblikah, kot je človek. Tudi funkcionalno prevladujejo hemisfere. V nasprotju s sesalci se zapleti prednjih možganov pri pticah izražajo predvsem v rasti bazalnih jeder (nuclei basales), ne pa da lok ostane tanek. 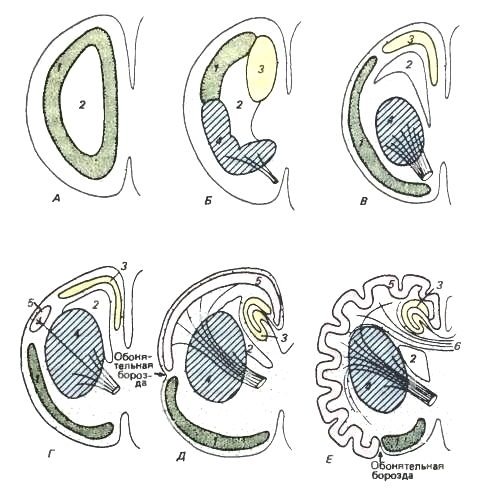
Shematski prerezi skozi levo hemisfero prednjih možganov. Barvne oznake so enake kot na prejšnji sliki.
1 - paleopalium; 2 - stranski prekat; 3 - arhipalij; 4 - bazalna jedra; 5 - neopalij; 6 - corpus callosum.
A je primitivna stopnja. Hemisfera je pravzaprav vohalni reženj. Šibko diferencirana siva snov se nahaja znotraj možganov.
B - stopnja, opažena pri sodobnih dvoživkah. Siva snov je še vedno daleč od zunanje površine, vendar je že razdeljena na paleopalij (= vohalni reženj), arhipalij (= hipokampus) in bazalna jedra (= striatum). Slednji prevzame pomen asociativnega centra, ki ima aferentne in eferentne povezave s talamusom (prikazano kot črte, ki simbolizirajo prerezane snope vlaken).
B - bolj progresivna stopnja, v kateri se bazalna jedra potopijo v hemisfero, medtem ko so se deli skorje premaknili nekoliko navzven.
G - stopnja, na kateri se nahajajo napredni plazilci. Pojavi se neopalij.
D - stopnja primitivnega sesalca. Neopalium se je povečal. Ima obsežne povezave z možganskim deblom. Arhipalij na medialni površini hemisfere se ovije kot hipokampus. Paleopalij je še vedno zelo razvit.
E - stopnja visoko organiziranega sesalca. Neopalium se izredno poveča in se zbere v gube. Paleopallium zavzema omejeno ventralno območje, ki predstavlja hruškast reženj. Razvije se corpus callosum - močan most, ki povezuje področja neopalija obeh hemisfer. (Po Romer in Parsons, 1992.) Lubje neopalija se imenuje novo skorja ali neokorteks (neocortex). Pri sesalcih služi kot središče višjega (pogojnega refleksa) živčna dejavnost usklajevanje dela drugih delov možganov. Od tu se impulzi pošiljajo v različne organe in tkiva telesa, tu pa se izvaja tudi regulacija. fizioloških procesov glede na okoljske razmere. Nova skorja je tista, ki kopiči sledi posameznih vzburjenj in njihovih kombinacij, zaradi česar se obogati delovni spomin, ki daje možnost izbire optimalnih rešitev v novih situacijah. Pogosto so te odločitve nove kombinacije predhodno znanih vedenjskih elementov, obstaja pa tudi razvoj in utrjevanje novih možnosti ukrepanja. Ko se razvija, nova skorja ne samo prevzame funkcije korelativnega in asociativnega centra za novo nastajajoče vrste višje živčne aktivnosti, ampak začne opravljati tudi številne funkcije, ki so prej pripadale središčem možganskega debla in bazalnih jeder. Hkrati se starodavni centri, ki nadzorujejo instinktivna dejanja, ne likvidirajo, temveč le podvržejo višjemu nadzoru.
V povezavi z razvojem neokorteksa streha srednjih možganov izgubi svoj prejšnji pomen in ostane le refleksno in prenosno središče. Slušni in drugi somatski senzorični impulzi se prenašajo naprej do talamusa, večina vidnih vlaken je tu prekinjena in vsi ti signali iz talamusa se po močnih živčnih snopih prenašajo v hemisfere. Podobne talamične povezave z bazalnimi jedri so nastale tudi pri nizko organiziranih skupinah vretenčarjev in so bile najbolj razvite pri pticah. Za razliko od ptic gre pri sesalcih večina vlaken skozi in skozi striatum in se razhaja na površino nove skorje. Tako se vanj steka celoten sklop senzoričnih podatkov, na podlagi katerih se v skorji sprejemajo ustrezne motorične »odločitve«.
Kot že omenjeno, se nekateri signali prenašajo iz skorje v male možgane skozi pons in zagotavljajo potrebne regulativne učinke. Neokorteks je povezan tudi s striatumom in celo s hipotalamusom – in s tem z avtonomnim živčni sistem. Vendar pa je glavnina motoričnih ukazov usmerjena po piramidni poti (tractus corticospinalis) – posebnem živčni snop, ki gre neposredno, brez preklapljanja, iz možganske skorje skozi srednje možgane v somatsko motorična področja možganskega debla in hrbtenjača. Istočasno se vlakna stranskega dela te poti križajo in inervirajo nasprotna stran telo (tj. levo vlakno inervira desna stran telo in obratno), ventralna vlakna pa ostanejo povezana s svojo stranjo telesa. Piramidna pot je prisotna le pri sesalcih, kar nazorno dokazuje dominanten položaj nove skorje pri njih. Ta struktura doseže svoj največji razvoj pri opicah in še posebej pri ljudeh, ki igrajo pomembno vlogo pri dvonožnem gibanju. Pri vrečarjih segajo piramidni aksoni le do torakalni, pri enojnem prehodu pa je piramidalna pot popolnoma odsotna. 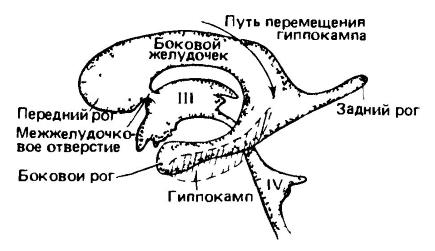
Prekati človeških možganov; stranski pogled z leve strani. Prekati so prikazani kot odlitki, medtem ko možganska tkiva niso prikazana. Z rastjo hemisfere prednjega možgana se stranski prekat razširi nazaj s tvorbo zadnjega roga v okcipitalnem režnju, v njegovem stranskem delu pa navzdol in naprej s tvorbo stranskega roga v okcipitalnem režnju. temporalni reženj. Podobni izrastki, usmerjeni nazaj in navzdol, so povzročili spremembe lokacije razne dele možgani. Hipokampus, ki se je razvil v dorzalnem položaju na medialni površini hemisfere, se je pri visoko razvitih sesalcih premaknil nazaj in navzdol v ventralni položaj. (Po Romerju in Parsonsu, 1992.) Ker je neokorteks tanka plast večplastnega celičnega materiala, pod katerim leži bela vlaknasta masa možganov, enostavno povečanje prostornine hemisfer ne more povzročiti sorazmernega širjenja skorje. Hkrati se lahko v naprednih oblikah površina lubja zaradi zlaganja znatno poveča. Gube, ki nastanejo na ta način, se imenujejo vijuge (gyri), globoke reže med njimi pa brazde (sulci). Oba vsebujeta skupne morfološke komponente. V najpreprostejšem primeru obstaja en globok silvijev žleb, ki ločuje čelni reženj (lobus frontalis) od temporalnega režnja (lobus temporalis). Nato se zgoraj in spredaj od Silvijeve brazde pojavi prečni Rolandov sulkus, ki od zgoraj ločuje čelni reženj od parietalnega (lobus parietalis). Pri primatih prečna brazda ločuje mali zadnjik okcipitalni reženj(lobus occipitalis). Poleg glavnih brazd se oblikujejo številne dodatne; njihovo število je še posebej veliko pri primatih in zobatih kitih. Prej je veljalo, da brazde v nekaterih primerih označujejo morfološke meje, ki ustrezajo določenim področjem skorje. Vendar pa so nadaljnje študije pokazale odsotnost fiksnega razmerja med porazdelitvijo zgibanja in strukturna enota korteksu (razen nosnega sulkusa in do neke mere centralnega sulkusa pri primatih, o katerem bomo govorili kasneje). Omeniti velja, da se je gubanje skorje razvilo v več evolucijskih deblih sesalcev precej neodvisno. Pri razmeroma primitivnih sesalcih, kot so monotremi, vrečarji in nekateri placentalni (žužkojedi, netopirji, glodavci, lagomorfi), je lubje skromneje razvito in ima gladko površino. 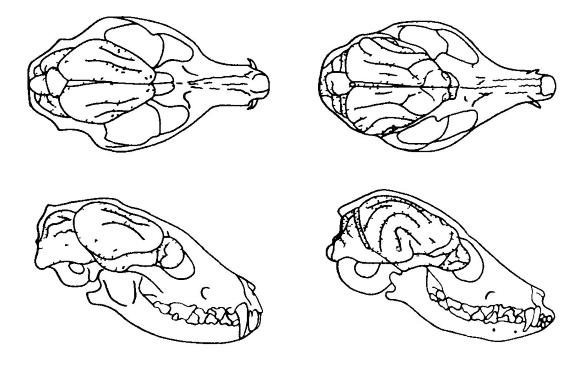
Lokacija možganov v lobanji fosilnega in živečega kanida. Opazno je povečanje velikosti in zapletov možganov, zlasti hemisfer prednjih možganov. Hesperocion ( Hesperocyon gregarius) (levo) je oligocenska oblika, ki je živela pred približno 30 milijoni let. Fenech ( Vulpes zerda) (desno) - moderna oblika podobne velikosti. (Po Romer in Parsons, 1992.) Za sivo snov neokorteksa je značilna kompleksna histološka zgradba. Pri placentnih sesalcih se razlikuje 6 plasti celic, ki ležijo ena nad drugo, in vlakna, ki vstopajo med njimi; to močno razlikuje neokorteks od preostalih delov skorje paleopalija in arhipalija, kjer lahko ločimo le 2 do 4 celične plasti. Po obstoječih ocenah pri sesalcih s še posebej veliki možgani, lahko število celic nove skorje doseže milijarde.
Bela snov, ki se nahaja pod sivo, poleg pahljače povezav, ki gredo od skorje do spodnjih delov možganov in nazaj, vključuje ogromno število prepletajočih se prečnih vlaken, ki povezujejo različne dele skorje. Tako nastala komisura je raztegnjena nazaj (glede na potek rasti hemisfer) in razdeljena na dve plošči, zraščeni vzdolž zadnjega roba. Spodnji, tanjši in s sprednjim robom odklonjen navzdol, je lok (fornix), komisura arhipalijske skorje (tj. hipokampusa). Zgornja, debelejša, vodoravno nameščena komisura pripada novi skorji in se imenuje corpus callosum (corpus callosum). Ta tvorba vam omogoča združevanje spomina obeh polobel in znatno poveča sposobnost možganov za učenje. Corpus callosum je zaradi znatnega razvoja nove skorje prisoten le v posteljicah, pri monotremih in vrečarjih so ga prikrajšani. Poleg tega imajo vsi sesalci sprednjo komisuro (commissura anterior), ki povezuje vohalne predele skorje. 
Slojevita razporeditev živčnih celic v skorji telencefalon sesalec (po Naumov in Kartashev, 1979.) Kompleksen sistem "prevodnikov", ki povezujejo vse dele korteksa, nakazuje, da je siva snov načeloma enotno izobraževanje, katerega vsi deli imajo enake sposobnosti za izvajanje katere koli funkcije možganskih hemisfer. Do neke mere je to res: poskusi kažejo, da je pri laboratorijskih živalih mogoče uničiti pomemben del nove skorje, ne da bi povzročili trajne poškodbe njihovih normalno aktivnost. Dokazi o poškodbah in morbidnih spremembah potrjujejo, da to velja tudi za človeški možgani. Hkrati je jasno, da so določeni predeli korteksa običajno povezani z izvajanjem precej specifičnih funkcij. Zgoraj smo omenili področja paleopalija in arhipalija, namenjena predvsem analizi vohalnih informacij in ohranjena v obliki hruškastega režnja oziroma hipokampusa. Diferenciacija posameznih območij poteka tudi v skorji neopalija. Sprednji del hemisfer vsebuje motorično področje. ki se nahaja tukaj Čelni reženj med drugim skrbi za komunikacijo živali, tudi akustično; pri ljudeh povezuje z govorom, torej drugo signalni sistem. Zadnji del hemisfer je povezan z zaznavanjem občutkov. v okcipitalnem in temporalni režnji obstajajo področja, ki nadzorujejo vid oziroma sluh. Še naprej, v bližini motoričnega področja, so območja, ki zaznavajo taktilne in proprioceptivne signale. Pri primatih osrednja brazda (sulcus centralis), ki prečka vrh poloble od medialne do lateralne površine, razmejuje (čeprav ne povsem natančno) motorično področje od senzoričnega. Vzdolž sprednjega roba osrednjega sulkusa so določena motorična področja nameščena v linearnem vrstnem redu, ki služijo vsakemu delu telesa in okončinam. Vzdolž zadnjega roba osrednjega sulkusa so v enakem vrstnem redu razporejena področja senzoričnega zaznavanja ustreznih delov telesa.
Tako je pri mnogih sesalcih skoraj vsa površina neokorteksa zasedena s področji, ki so bolj ali manj tesno povezana z nekaterimi senzoričnimi oz. motorične funkcije. Čeprav je osrednji sulkus morda odsoten, imajo v večini primerov placente podobno linearno razporeditev senzoričnih in motoričnih področij drug proti drugemu. Pri vrečarjih (in med placentalnimi pri ksenartrasih) je »označenost« telesnih področij približno enaka, le da senzorična področja niso ločena od motoričnih, ampak so prepredena z njimi. Toda na primer pri ljudeh ta specifična funkcionalna področja zasedajo relativno malo prostora na površini neokorteksa. Med njimi so nastala velika območja sive snovi (eno posebej veliko takšno območje zavzema večinačelni reženj), ki niso povezani s posebnimi senzoričnimi ali motoričnimi funkcijami. Zato se ta področja pogosto imenujejo "prazne lise", čeprav, kot kaže poškodba teh območij, se prav v njih nahajajo naše višje mentalne sposobnosti, vključno z možnostmi učenja, pobudo, predvidevanjem in presojo. Vendar pa obstajajo področja, ki jih je mogoče odstraniti brez resnih posledic za intelektualno dejavnost. 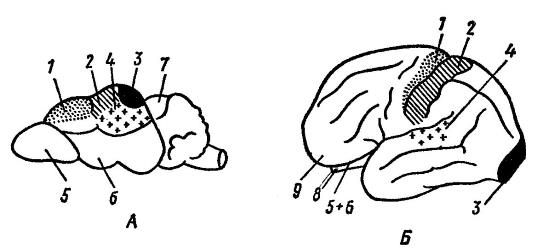
Funkcionalni centri možganske skorje rovke ( sorex sp.) (A) in človek ( Homo sapiens
) (B) (po Naumov in Kartashev, 1979):
1 - motorni center; 2 - središče kožno-mišične občutljivosti; 3 - vizualni center; 4 - slušni center; 5 - vohalna žarnica; 6 - vohalni režnji; 7 - streha srednjih možganov; 8 - mali možgani; 9 - čelni reženj. Na razvoj možganov močno vplivata zunanje okolje in motorična (prehrambena, obrambna) aktivnost. Hkrati je razvoj različnih delov možganov določen predvsem z načini iskanja hrane: pri psu ( canis lupus), ki pri tem procesu uporablja voh, je bolj razvit vohalni predel; pri mački ( Felis silvestris), iskanje hrane s pomočjo vida – vizualno; pri makaku ( Macaca mulatta), ki uporablja vid in sluh – vizualni in slušni.
Običajno se domneva, da velikost možganskih hemisfer določa razlike v duševnih sposobnostih različnih sesalcev. V nekem smislu je to res, vendar z velikimi zadržki. Večji možgani so sestavljeni iz večživčne celice. Če je območje obstoječe površine možganske skorje kakor koli povezano z inteligenco, potem je očitno, da bo od dveh različic možganov enake velikosti bolj razvita tista z brazdasto površino, možgani z gladko površino pa manj razviti. Velikost same živali vpliva tudi na volumen možganov. To se zgodi že zato, ker morajo imeti možgani večja območja, da služijo večjim senzoričnim in motoričnim povezavam. Vendar pa povečanje velikosti možganov ni povsem sorazmerno s telesno maso, tako da imajo velike živali običajno relativno manjše možgane brez očitne izgube inteligence. Tako absolutna velikost možganov ni brezpogojno merilo inteligence. Na to zagotovo nakazuje podatek, da so lahko kitovi možgani po prostornini petkrat večji od človeških.
Primerjava možganov nekaterih sesalcev:
1 - konj; 2 - pes; 3 - kenguru; 4 - oseba; 5 - slon. Odstotek možganov v celotni telesni masi se imenuje cefalizacijski indeks. Pri velikih žužkojedih je približno 0,6%, pri majhnih - do 1,2%, pri velikih kitovih - približno 0,3%, pri majhnih - do 1,7%. Večina primatov ima indeks cefalizacije 1-2%. Pri ljudeh doseže 2-3%, nekatere majhne opice s širokim nosom pa imajo možgane, katerih masa je do 7% telesne teže. Hkrati se pri sodobnih plazilcih in pticah indeks cefalizacije giblje od 0,05 do 0,5%.
Spodaj je masa možganov nekaterih sesalcev (masa živali je navedena v oklepajih):
virginijski oposum ( Didelphis virginiana) - 7,6 g (5 kg);
koala ( Phascolarctos cinereus) - 19,2 g (8 kg);
slon ( afriški loksont (Loxodonta africana).) - 6000 g (5000 kg);
navadni jež ( Erinaceus europaeus) - 3,3 g (1 kg);
hišna miška ( Mus mišica) - 0,3 g (0,02 kg);
siva podgana ( Rattus norvegicus) - 2 g (0,3 kg);
navadna veverica ( Sciurus vulgaris) - 7 g (0,4 kg);
evropski zajec ( Oryctolagus cuniculus) - 11 g (3 kg);
domači konj ( Equus ferus) - 530 g (500 kg);
črni nosorog ( Diceros bicornis) - 500 g (1200 kg);
belorepec ( Odocoileus virginianus) - 500 g (200 kg);
žirafa ( Giraffa camelopardalis) - 680 g (800 kg);
domače ovce ( Ovis orientalis) - 140 g (55 kg);
domači bik ( Bos primigenius) - 490 g (700 kg);
baktrijska kamela ( camelus bactrianus) - 762 g (700 kg);
povodni konj ( Hippopotamus amphibius) - 580 g (3500 kg);
belocevi delfin ( Delphinus delphis) - 815 g (60 kg);
narval ( Monodon monoceros) - 2997 g (1578 kg);
kit glavač ( Physeter macrocephalus) - 8028 g (35833 kg);
modri kit ( Balaenoptera musculus) - 3636 g (50900 kg);
domača mačka ( Felis silvestris) - 25 g (3 kg);
lev ( panthera leo) - 270 g (250 kg);
navadna lisica ( Vulpes vulpes) - 53 g (4,5 kg);
domači pes ( canis lupus) - 64 g (10 kg);
polarni medved ( Ursus maritimus) - 500 g (700 kg);
mrož ( Odobenus rosmarus) - 1130 g (700 kg);
marmozetka Geldi ( Callimico goeldii) - 7 g (0,2 kg);
beločela kapucinka ( Cebus albifrons) - 57 g (1 kg);
opica rezus ( Macaca mulatta) - 88 g (6,5 kg);
pavijan ( Papio cynocephalus) - 200 g (25 kg);
srebrni gibon ( Hylobates moloch) - 112 g (6,5 kg);
kalimantanski orangutan ( Pongo pygmaeus) - 413 g (50 kg);
zahodna gorila ( gorila gorila) - 506 g (126 kg);
navadni šimpanz ( Pan trogloditi) - 430 g (55 kg);
razumna oseba ( Homo sapiens) - 1400 g (72 kg).
Iz navedenih primerov je razvidno, da so pri manjših sesalcih možgani skoraj vedno relativno večji, z večanjem velikosti telesa živali pa se relativna velikost možganov zmanjšuje. To je še posebej izrazito pri sorodnih vrstah sesalcev – na primer pri mački ( Felis silvestris) in lev ( panthera leo). Zelo prepričljivi v tem smislu in psi različnih pasem. Če telesne mase najmanjših in največ velike pasme so približno v razmerju 1:33, potem so mase možganov pri istih pasmah povezane kot 1:3. 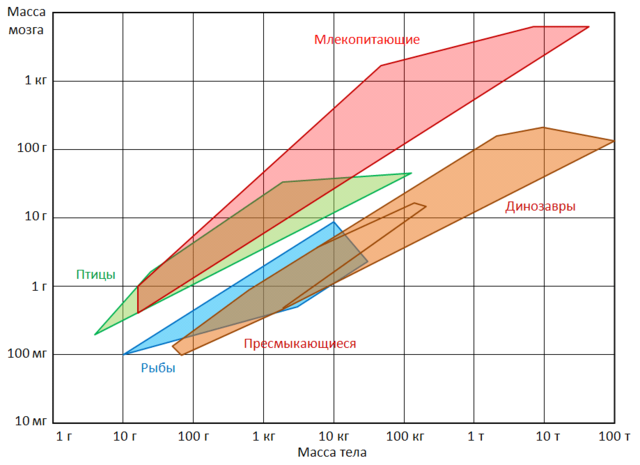
Razpon možganske in telesne mase za nekatere skupine vretenčarjev. Pri udomačenih živalih, ki jim je odvzeta potreba po hrani in obrambi pred sovražniki, se velikost možganov znatno zmanjša. Na primer, volumen možganov volka ( canis lupus) je 30 % večji od psa enake velikosti. Zanimivo je, da te spremembe ne vplivajo le na tradicionalno domače živali, temveč tudi na predstavnike prostoživečih vrst, ki so bile nekaj časa v ujetništvu. Ja, lisice Vulpes vulpes), rojeni v naravi, vendar že od prvih dni življenja v ujetništvu, imajo manjše možgane kot njihovi sorodniki, ki živijo v naravnih razmerah. Hkrati razlike dosežejo 20%, kar približno ustreza razliki v volumnu možganov med divjimi in pravimi domačimi živalmi. Zmanjšanje možganov, čeprav manj izrazito (približno 5 %), so ugotovili pri volkovih v ujetništvu ( Canis), dihurji ( Mustela), podgane ( Rattus). Hkrati pa zmanjšanje ne zajame vseh delov možganov, temveč le tista področja, ki so povezana z delom čutnih organov. Najbolj presenetljivo je, da se pri izpuščenih hišnih ljubljenčkih poveča teža možganov. Na primer, divje mačke imajo približno 10 % večje možgane kot njihove domače mačke. Znatno povečanje možganov je bilo ugotovljeno tudi pri divjih kuncih ( Oryctolagus cuniculus) na Kerguelenskih otokih. divji osli ( Equus asinus) V Južna Amerika imel 15% veliki možgani kot domače. Zanimivo je tudi, da so možgani neandertalca ( Homo neanderthalensis) in paleolitik Homo sapiens ( Homo sapiens) so bili nekoliko večji od možganov sodobnega človeka.
Ugotovljeno je bilo, da pri mnogih sesalcih opazimo motorično asimetrijo, to je prevladujočo uporabo desne ali leve polovice telesa. Na primer, ko preučujemo nezlomljene konje ( Equus ferus) zabeležili, po kateri nogi živali začnejo hoditi, po kateri strani najraje obidejo ovire in po kateri strani najraje ležijo v seniku. Posledično se je večina kobil izkazala za desničarje, večina žrebcev pa je bila levičarjev. Približno 10 % konj ni dalo prednosti ne desni ne levi ud. Po opazovanjih je približno 90% mrožev ( Odobenus rosmarus) z desno plavutjo izkopljejo mehkužce iz morskega mulja. Lulling dojenčki, približno 80% samic šimpanzov ( Pan) in gorile ( Gorilla) pritisnejo glavo na levo stran prsnega koša (približno enak odstotek je pri ženskah). podgane ( Rattus), ki vodi iskanje hrane s pomočjo vibris, ki se nahajajo na desna stran nagobčniki, so večji plen kot njihovi levičarski sorodniki.

