14.1. SPLOŠNE DOLOČBE
Končni možgani (telencephalon) ali veliki možgani (cerebrum), ki se nahaja v supratentorialnem prostoru lobanjske votline je sestavljen iz dveh velikih
hemisfere (možganska gemisferija),ločena z globoko vzdolžno režo (fissura longitudinalis cerebri), v katerega je potopljen srp veliki možgani (falx cerebri) ki predstavlja podvajanje trdne snovi možganske ovojnice. Velike hemisfere možganov predstavljajo 78% njihove mase. Vsak od hemisfere Ima režnja: čelni, temenski, temporalni, okcipitalni in limbični. Pokrivajo strukture diencefalona ter možganskega debla in malih možganov, ki se nahajajo pod cerebelarnim plaščem (subtentorialno).
Čeprav lahko človeški možgani kot celota opravljajo številne naloge in se osredotočajo na bolj specifične kot druge, je resnica taka, da je človeški mentalni aparat bolj specializiran, kar pomeni, da so deli vsake poloble osredotočeni na določene naloge. , obstaja bolj integralna uporaba obeh hemisfer več dejavnosti, kar imenujemo večopravilnost.
Segmentacija informacij
V moških možganih so informacije razdeljene v skupine ali segmente, ki niso nujno povezani. Pri ženskah je vse povezano kot sistem. Zato so ženske nagnjene k celovitosti. Zaradi teh razlik med možgani moških in žensk se moški ponavadi popolnoma osredotočijo na dejavnosti, ki jih izvajajo, ne da bi jih motili drugi vidiki. Po drugi strani pa ženske vsak dogodek na svetu gledajo kot popolno risbo, kjer vsak del medsebojno deluje.
Vsaka od možganskih hemisfer ima tri površine: zgornji stranski ali konveksni (slika 14.1a), - konveksen, obrnjen proti kostem lobanjskega oboka; notranja (slika 14.1b), ki meji na velik falciformni proces, in nižja ali bazalna (slika 14.1c), ki ponavlja relief dna lobanje (sprednja in srednja jama) in cerebelarni tenon. Na vsaki hemisferi ločimo tri robove: zgornji, spodnji notranji in spodnji zunanji ter tri pole: sprednji (čelni), zadnji (okcipitalni) in stranski (temporalni).
Občutki moškega in ženske
Tudi v čutenju in dojemanju sveta so razlike med moškim in žensko. Pri človeku je vid prevladujoča funkcija, pot, po kateri večina informacij vstopi v živčni sistem. Pri ženskah vsi sistemi delujejo skupaj kot pri slepcih, imajo zelo razvit sluh, voh in dotik, a so dodani vidu.
Tudi velikost možganov se med moškimi in ženskami razlikuje in je ponavadi nekoliko večja. To ne pomeni, da so pametnejši, saj znanstveniki verjamejo, da je namen te ekspanzije povezan z velikim obsegom moških mišic in ne z mentalnimi procesi. Moški imajo v telesu 20-krat več testosterona kot ženske. Zato se njegov živčni sistem kaže v bolj agresivnih in dominantnih odnosih in zato je seks bolj zaželen s fizičnega vidika. Ženske so bolj občutljive in njihov um išče spolni užitek, osredotočen na čutnost in naklonjenost.
Votlina vsakega hemisfera je stranski prekat možganov medtem ko je levi stranski prekat prepoznan kot prvi, desni - drugi. Lateralni prekat ima osrednji del, ki se nahaja globoko v parietalnem režnju (lobus parietalis) in trije rogovi, ki segajo iz njega: sprednji rog prodre v čelni reženj (lobus frontalis), nižje - do časovnega (lobus temporalis), posteriorno - v okcipitalnem (lobus occipitalis). Vsak od stranskih ventriklov komunicira s tretjim ventriklom možganov preko interventrikularnega luknja Monroe.
Možganske hemisfere in kognitivne sposobnosti
In na koncu bomo rekli, da ima pri ljudeh večjo prevlado leva hemisfera možganov. Pri ženskah obstaja ravnovesje v uporabi obeh hemisfer, leve in desne. Zato lahko pri kognitivnih veščinah opazimo, da so moški zelo nagnjeni k temu, da so dobri v matematiki in računanju, medtem ko imajo ženske sposobnost jezika, dejavnosti, ki zahteva uporabo več področij znotraj.
Pokažejo nam, kako v evolucijskem procesu deluje živčni sistem oba spola sta bila specializirana za zagotavljanje prihodnosti vrste. Nikoli ne pozabite, da so razlike povezane s fizičnimi in duševnimi strukturami in da nihče ne sme podcenjevati človeške vrednosti vsakega od žanrov. Smo si različni, a se dopolnjujemo.
Osrednji deli medialne površine obeh hemisfer so med seboj povezani s cerebralnimi komisurami, od katerih je najbolj masivna corpus callosum, in strukturami diencefalona.
Telencefalon je tako kot drugi deli možganov sestavljen iz sivih in belo snov. Siva snov se nahaja v globini vsake hemisfere, kjer tvori subkortikalne vozle, in na obrobju prostih površin hemisfere, kjer sestavlja možgansko skorjo.
V knjigi Pomanjkanje pozornosti pri otrocih in mladostnikih je ena od stvari, ki jih avtorica Amanda Cespedes omenja, pomembnost poznavanja možganov in njihovih funkcij kot strategije za razumevanje in odzivanje na potrebe otrok in mladostnikov. Pravzaprav učenje pridobiva novo znanje. Da bi to dosegli, obstaja nekaj osnovnih možganskih funkcij, ki pridejo v poštev od kognitivnih do čustvenih. Zlasti študija človeški možgani v otroštvu temelji na obstoju nevronske mreže pri katerem se informacije obdelujejo na enak način kot računalnik pri obdelavi podatkov.
Osnovna vprašanja o zgradbi, funkcijah bazalnih ganglijev in različicah klinična slika ob njihovem porazu so obravnavani v poglavjih 5, 6. Možganska skorja je približno
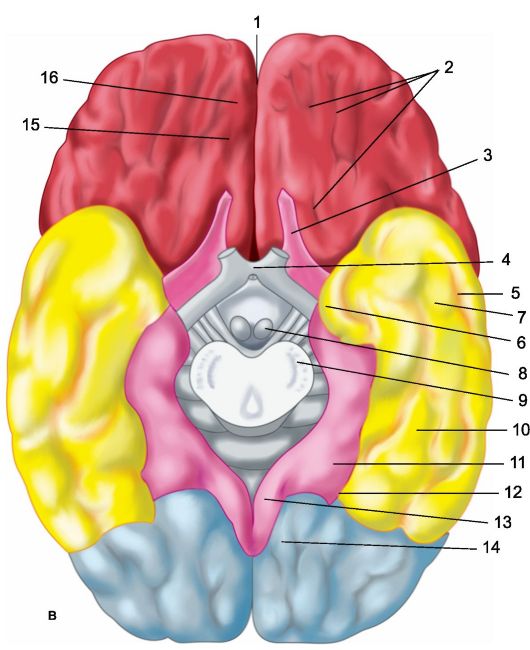
riž. 14.1.Hemisfere možganov.
To pomeni, da otrokovi možgani niso omejeni na sprejemanje informacij in v tem procesu postane na stotine tisoč nevronov na voljo za učenje in informacijski proces, ki se odvija v dveh hemisferah možganov: na desni in na levi. V knjigi The Disability of ADHD in Children and Adolescents avtorice Amande Cespedes se avtorica poglobi v to temo, izpostavi značilnosti vsake hemisfere in poda konkretni primeri situacije ali dejanja, v katerih se pri otrocih in učiteljih kaže delovanje obeh hemisfer, Zakaj?
Ker je le z globokim razumevanjem možganov mogoče oblikovati učne strategije, ki bodo ustrezale posebnim potrebam. Dozoreva pred rojstvom in krepi svoje funkcionalne mreže v prvih petih letih življenja. V tej polobli je šest talentov: vizualni, prostorska logika, melodični ritem, kineziko, medosebni in naturalistični. Desna polobla ni urejen z objektivno analizo zunanje realnosti in tako dopušča obstoj subjektivnih realnosti, ki spodbujajo domišljijo.
a - zgornja stranska površina leve poloble: 1 - osrednji sulkus; 2 - orbitalni del spodnjega čelnega gyrusa; I - čelni reženj; 3 - precentralni girus; 4 - precentralna brazda; 5 - zgornji čelni girus; 6 - srednji čelni girus; 7 - tegmentalni del spodnjega čelnega gyrusa; 8 - spodnji čelni girus; 9 - stranska brazda; II - parietalni reženj: 10 - postcentralni girus; 11 - postcentralna brazda; 12 - intraparietalni utor; 13 - supramarginalni girus; 14 - kotni girus; III - temporalni reženj: 15 - zgornji temporalni girus; 16 - zgornji temporalni sulkus; 17 - srednji temporalni girus; 18 - srednji temporalni sulkus; 19 - spodnji temporalni girus; IV - okcipitalni reženj: b - medialna površina desne poloble: 1 - paracentralni lobule, 2 - precuneus; 3 - parieto-okcipitalni sulkus; 4 - klin, 5 - jezikovni gyrus; 6 - stranski okcipitotemporalni girus; 7 - parahipokampalni girus; 8 - kavelj; 9 - trezor; 10 - corpus callosum; 11 - zgornji čelni girus; 12 - cingularni girus; c - spodnja površina možganskih hemisfer: 1 - vzdolžna interhemisferna razpoka; 2 - orbitalne brazde; 3 - vohalni živec; 4 - optična kiazma; 5 - srednji temporalni sulkus; 6 - kavelj; 7 - spodnji temporalni girus; 8 - mastoidno telo; 9 - osnova možganskega debla; 10 - stranski okcipitotemporalni girus; 11 - parahipokampalni girus; 12 - stranski utor; 13 - cingularni girus; 14 - jezikovni gyrus; 15 - vohalni utor; 16 - neposredni girus.
Kognitivni slog tega področja možganov je heterogenega tipa, kar pomeni, da poteka od začetne ideje do končne točke, ki je nejasno povezana s to začetno idejo. Intuicija in celostno procesiranje sta na tej polobli zelo pomembni. Razumevanje delovanja te strani možganov je koristno za ustvarjanje pedagoških praks, ki temeljijo na tem, zato avtor podrobno in preprosto opisuje dejavnosti, ki se pojavljajo na tej strani otrokovih možganov, in strategije, ki jih učitelj uporablja za uporabo te strani. hemisfera.
Dozori po šestih mesecih življenja, glavno zorenje pa se konča v puberteti. Talenti, ki temeljijo na tej strani možganov, so: jezikovna, matematična simbolna logika in medosebna ali refleksivna. Leva polobla otroka postavlja v stik s pojmovnim znanjem, vendar je tako učenje krhko in dovzetno za pozabljanje. Za konsolidacijo teh lahko rečemo, da daje prednost podpori desne poloble. Tako kot na desni hemisferi Amanda našteva dejanja, ki se pojavljajo na tej strani otrokovih in učiteljevih možganov.
3-kratna površina hemisfer, vidna med zunanjim pregledom. To je posledica dejstva, da je površina možganskih hemisfer prepognjena, ima številne vdolbine - brazde (sulci cerebri) in se nahaja med njimi zvitki (gyri cerebri). Možganska skorja pokriva celotno površino vijug in brazd (zato je njeno drugo ime palij - plašč), včasih pa prodre v veliko globino v možgansko snov.
Naučite se zamenjati številke, da oblikujete številke za izvajanje operacij in reševanje problemov. Razumevanje, pravi avtorica, zasnove možganov kot celote in njegovega izvršilnega mehanizma pomeni imeti pogoje za razumevanje otroka ali mladostnika in poseganje v pedagoške strategije, ki so prilagojene potrebam vsakega učenca. Vendar, koliko veste o možganih in kako to znanje uporabite za ustvarjanje orodij, ki zagotavljajo ne le učenje, ampak tudi gradnjo družbenega, šolskega in družinskega življenja?
Kaj so režnji v možganskih hemisferah?
Dejstvo je, da so pogovori staršev z otroki ključni v učnih procesih. Po mnenju mnogih strokovnjakov jim to pomaga razviti jezik, socialne veščine in druge stvari, ki so ključne za razumevanje sveta okoli njih. Vzporedno s tem so prišla nova odkritja, ki poudarjajo pomen komuniciranja s cilji.
Resnost in lokacija brazd in konvolucij možganskih polobel sta do neke mere spremenljiva, vendar se glavne oblikujejo v procesu ontogeneze in so stalne, značilne za vsak normalno razvit možgan.
14.2. VEČJI ZGRIŽKI IN PRIJEMI MOŽGANSKIH POLOBL
Zgornja stranska (konveksilna) površina hemisfer (slika 14.1a). Največji in najgloblji bočna brazda (sulcus lateralis),ali sylvian brazda, - ločuje čelni in sprednji del parietalnega režnja od spodnjega temporalni reženj. Čelni in temenski reženj sta ločena osrednja ali Rolandova brazda(sulcus centralis), ki prereže zgornji rob hemisfero in gre navzdol in naprej vzdolž njene konveksilne površine, nekoliko manj od stranskega sulkusa. Parietalni reženj je ločen od okcipitalnega režnja, ki se nahaja za njim, s parietalno-okcipitalnimi in prečnimi okcipitalnimi žlebovi, ki potekajo vzdolž medialne površine poloble.
Otoček, limbična skorja
Po mnenju strokovnjakov so nekatere raziskave, ki temeljijo na opazovanju družin, pokazale, da pogovori o številkah med vsakodnevnimi aktivnostmi in igrami napovedujejo otrokovo poznejše matematično znanje. Poleg tega dokazi še navajajo, da to zgodnje učenje dolgoročno prispeva k razumevanju matematike in jezika. Čeprav so otroci rojeni za učenje številk, potrebujejo podporo pogovarjamo se o povezovanju imen števil z točno število naučiti se predstaviti različno število predmetov.
V čelnem režnju pred osrednjim girusom in vzporedno z njim je precentral (gyrus precentralis), oz sprednji osrednji, gyrus, ki je spredaj omejen s precentralnim sulkusom (sulcus precentralis). Zgornji in spodnji čelni utori odhajajo spredaj od precentralnega sulkusa in delijo konveksilno površino sprednjih delov čelnega režnja na tri čelne giruse - zgornji, srednji in spodnji. (gyri frontales superior, media et inferior).
Podpora pri učenju številk
Ko imajo starši te pogovore s svojimi otroki, lahko ugotovijo. To razumevanje števila znakov je res pomembno, ko začnejo med delom delati z veliko večjimi in bolj zapletenimi količinami šolski proces. Vendar je pomembno, da otroci poznajo zaporedje številk, vendar pa bi morali starši z otroki vaditi veliko več, kot se zanje smatra. Če tega ne storite, bo otrok težko razumel, kaj pomenijo imena številk in kako jih uporabljati. Zato jim strokovnjaki ponujajo nekaj idej v podporo, preden se z matematiko srečajo v šoli.
Sprednji del konveksilne površine parietalnega režnja se nahaja za osrednjim postcentralnim sulkusom (gyrus postcentralis), ali zadnji osrednji, gyrus. Zadaj meji na postcentralni sulkus, od katerega se nazaj razteza intraparietalni sulkus. (sulcus intraparietalis), ki ločuje zgornje in spodnje parietalne lobule (lobuli parietales superior et inferior). V spodnjem parietalnem režnju se razlikuje supramarginalni gyrus (gyrus supramarginalis), ki obdaja zadnji del stranskega (Sylvijevega) žleba in kotni girus (girus angularis), meji na zadnji del zgornje temporalne vijuge.
Več kot samo branje številčno zaporedje, šteje in razpravlja o številnih vidnih predmetih. Prešteje in razvrsti število predmetov v nizu. To bo vašim otrokom omogočilo, da se povežejo s postopkom štetja glede na velikost kompleta.
Uporabite imena številk, ki so nekoliko večja od številk, ki jih vaš otrok že razume. Poglejte kretnje s številkami otrok. Velikokrat se pravilno odzovejo s kretnjami, ne pa z besedami. V pomoč so tudi vaše kretnje. Ko z otroki štejete števila, pokažite na vsak predmet, ki je del štetja. S prsti pokažite količino, o kateri govorite.
Na konveksalni površini okcipitalnega režnja možganov so brazde plitke in se lahko znatno razlikujejo, zaradi česar je tudi narava konvolucij, ki se nahajajo med njimi, spremenljiva.
Konveksitalno površino temporalnega režnja delita zgornja in spodnja temporalna brazda, ki sta skoraj vzporedna z lateralno (Silvijevo) brazdo in delita konveksitalno površino temporalnega režnja na zgornjo, srednjo in spodnjo temporalno vijugo. (gyri temporales superior, media et inferior). Zgornji temporalni girus tvori spodnjo ustnico stranskega (Sylvijevega) sulkusa. Na svoji površini obrnjena
Izkoristite količino Vsakdanje življenje. Med branjem knjig vadite štetje. Otroke prosite, naj med svojimi dnevnimi rutinami izberejo določeno število predmetov. Med igro primerjajte število. Pogovarjajte se s svojimi otroki o tem, kako uporabljate številke v vsakdanjem življenju. Na primer, analizirajte, kako jih uporabljate, da veste, kaj kupiti, načrtujete, koliko boste pojedli, ali pripravite mizo.
Če podpiramo proces branja, zakaj ne bi enako storili s številkami? Center za razvoj otrok na Univerzi Harvard je večkrat govoril o pomenu dojemljivih in obzirnih interakcij med odraslimi in otroki. To je, pravijo raziskovalci centra, temeljnega pomena za izgradnjo trdne možganske podlage za učenje in prihodnji razvoj. Ta komunikacija ali interakcija se imenuje "serviraj in vrni" in je bistvena za gradnjo temeljev učenja in zagotavljanje optimalnega dobrega počutja.
strani stranske brazde je več prečnih majhnih brazd, ki na njej poudarjajo majhen prečni girus (Geschlov gyrus), ki se vidi šele z razmikom robov stranske brazde.
Sprednji del stranskega (silvijskega) žleba je vdolbina s širokim dnom, ki tvori t.i. otok (otok) oz insula (lubus insularis). Zgornji rob stranske brazde, ki pokriva ta otok, se imenuje pnevmatika (operkulum).
Struktura temporalnega režnja
Zdaj, da bi vadili to "igro", ki vključuje obe strani, obstajajo dejanja ali koraki, ki jih je mogoče izvesti, kaj je to? Opazujte pri interakciji z otroki. Gledate nekaj posebnega ali povzročate hrup? Imate poseben izraz na obrazu? Ali premikate roke ali noge? Te stvari, pravijo v centru, so storitve in pomembno je, da jih spremljajo ves dan v različnih situacijah, saj tako spoznavajo veščine, interese in potrebe otrok. Tako jih boste motivirali za raziskovanje in okrepili medsebojno vez.
Notranja (medialna) površina poloble (Slika 14.1b). Osrednji del notranje površine hemisfere je tesno povezan s strukturami diencefalona, od katerih ga ločujejo tiste, ki so povezane z velikimi možgani. trezor (fornix) in corpus callosum (corpus callosum). Slednji je na zunanji strani omejen z brazdo corpus callosum (sulcus corporis callosi), začenši na sprednji strani - kljun (roster) in se konča na odebeljenem zadnjem koncu (splenij). Tukaj sulkus corpus callosum preide v globoko hipokampalno brazdo (sulcus hippocampi), ki prodre globoko v snov poloble in jo pritisne v votlino spodnjega roga. stranski ventrikel, kar povzroči nastanek tako imenovanega amonijevega roga.
Objemi, ljubeče besede, igra, obrazna mimika in druge dejavnosti so lahko način vračanja otrokovih uslug. Na ta način jim ponudijo oporo in jih motivirajo za nadaljnje služenje, kar je v centru, kot pravijo, neke vrste nagrada za otroško radovednost in zanimanje. To, da od odrasle osebe ne dobijo ničesar v zameno, je zanje lahko vir stresa. Po drugi strani pa, ko prejmejo odgovor, morda vedo, da so njihove misli in občutki poleg skrbi pomembni.
Pri vračanju storitev je pomembno, da poimenujete, kaj otroci vidijo, čutijo ali počnejo. Tako boste ustvarili pomembne jezikovne povezave na ravni možganov, še preden boste razumeli besede in govorili. Na primer, če otrok kaže na svoje noge, lahko izgovorite besedne zveze, kot je "Da, to so noge." Zakaj je pomembno? Ker mu boste na ta način pomagali razumeti svet okoli sebe in jim dali vedeti, kaj lahko pričakujejo od njega. Poimenovanje stvari je tudi način, kako jim dati besede, ki jih lahko uporabljajo, in jim pokazati, da vam je mar.
Nekoliko odmaknjeni od sulkusa corpus callosum in hipokampalne brazde se nahajajo corpus callosum, subparietalne in nosne brazde, ki so nadaljevanje drug drugega. Ti utori omejujejo od zunaj ločni del medialne površine možganske hemisfere, znan kot limbični reženj(lobus limbicus). V limbičnem režnju sta dva zavoja. Zgornji del limbičnega režnja je zgornji limbični (superiorni robni) ali pas, gyrus (girus cinguli), spodnji del tvori spodnji limbični girus ali girus morskega konjička (girus hippocampi), ali parahipokampalni girus (girus parahypocampalis), pred katerim je kljuka (uncus).
Okoli limbičnega režnja možganov so tvorbe notranje površine čelnega, temenskega, okcipitalnega in temporalni reženj th. Večino notranje površine čelnega režnja zavzema medialna stran zgornjega čelnega girusa. Na meji med čelnim in parietalnim režnjem možganske hemisfere se nahaja paracentralnega lobula (lobulis paracentralis), ki je tako rekoč nadaljevanje sprednjega in zadnjega osrednjega vijuga na medialni površini hemisfere. Na meji med parietalnim in okcipitalnim režnjem je jasno viden parietalno-okcipitalni sulkus. (sulcus parietooccipitalis). Od dna odhaja nazaj ostrožna brazda (sulcus calcarinus). Med temi globokimi brazdami je trikotna vijuga, znana kot klin. (cuneus). Pred klinom je štirikotni girus, povezan z parietalni reženj možgani - precuneus.
Spodnja površina hemisfere (slika 14.1c). Spodnjo površino možganske hemisfere sestavljajo čelne, temporalne in čelne tvorbe okcipitalni reženj th. sosednji srednja črtačelni reženj je rectus gyrus (girus rectus). Zunaj je omejen z vohalnim utorom (sulcus olfactorius), na katerega od spodaj mejijo tvorbe olfaktornega analizatorja: olfaktorni bulbus in olfaktorni trakt. Bočno od njega, do stranskega (Sylvijevega) žleba, ki se razteza na spodnjo površino čelnega režnja, so majhni orbitalni vijugi (gyri orbitalis). Stranske dele spodnje površine hemisfere za stranskim sulkusom zaseda spodnji temporalni girus. Medialno od njega je lateralni temporo-okcipitalni girus. (gyrus occipitotemporalis lateralis), ali vretenasti žleb. pred-
njeni oddelki z znotraj meja na gyrusu hipokampusa, zadnja pa na lingvalnem (gyrus lingualis) ali medialni temporoccipitalni girus (gyrus occipitotemporalis medialis). Slednji s svojim zadnjim koncem meji na ostrožni žleb. Sprednji deli fusiformne in lingvalne vijuge pripadajo temporalnemu režnju, zadnji deli pa okcipitalnemu režnju možganov.
14.3. BELA SNOVI VELIKE HEMISFERE
Bela snov možganskih hemisfer je sestavljena iz živčna vlakna, pretežno mielin, ki predstavlja poti, ki zagotavljajo povezave med nevroni korteksa in grozdi nevronov, ki tvorijo talamus, subkortikalne vozle, jedra. Glavni del bele snovi možganskih hemisfer se nahaja v njegovi globini polovalno središče ali sijoča krona (korona radiata), sestavljena predvsem iz aferentne in eferentne projekcija poti, ki povezujejo možgansko skorjo s podkortičnimi vozli, jedri in retikularno snovjo diencefalona in možganskega debla, s segmenti hrbtenjača. Posebej kompaktno se nahajajo med talamusom in subkortikalnimi vozlišči, kjer tvorijo notranjo kapsulo, opisano v 3. poglavju.
Imenujejo se živčna vlakna, ki povezujejo dele skorje ene poloble asociativno. Čim krajša so ta vlakna in povezave, ki jih tvorijo, tem bolj so površinska; daljše asociativne povezave, ki se nahajajo globlje, povezujejo relativno oddaljene dele možganske skorje (sl. 14.2 in 14.3).
Imenujejo se vlakna, ki povezujejo možganski polobli in imajo zato skupno prečno usmeritev komisura, ali spanje. Komisuralna vlakna povezujejo enake dele možganskih hemisfer, kar ustvarja možnost kombiniranja njihovih funkcij. Oblikujejo tri konice veliki možgani: najbolj masivni med njimi - corpus callosum (corpus callosum), poleg tega sestavljajo komisuralna vlakna sprednja komisura, ki se nahaja pod kljunom corpus callosum (rostrum corporis collosum) in povezuje tako vohalne regije kot tudi komisura oboka (commissura fornicis), ali hipokampalna komisura, ki jo tvorijo vlakna, ki povezujejo strukture amonovih rogov obeh hemisfer.
V sprednjem delu corpus callosum so vlakna, ki povezujejo čelne režnje, nato so vlakna, ki povezujejo parietalne in temporalne režnje, zadnji del corpus callosum povezuje okcipitalne režnje možganov. Sprednja komisura in komisura forniksa v glavnem združujeta odseke starodavne in stare skorje obeh polobel; sprednja komisura poleg tega zagotavlja povezavo med njihovim srednjim in spodnjim časovnim vijugom.
14.4. Vohalni sistem
V procesu filogeneze je razvoj velikih možganov povezan z nastankom vohalnega sistema, katerega funkcije prispevajo k ohranjanju sposobnosti preživetja živali in niso majhnega pomena za življenje ljudi.
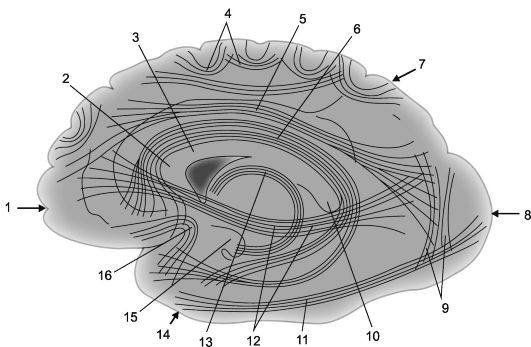
riž. 14.2.Asociativne kortikalno-kortikalne povezave v možganskih hemisferah [po V.P. Vorobjov].
1 - čelni reženj; 2 - koleno corpus callosum; 3 - corpus callosum; 4 - ločna vlakna; 5 - zgornji vzdolžni nosilec; 6 - cingularni girus; 7 - parietalni reženj, 8 - okcipitalni reženj; 9 - navpični snopi Wernickeja; 10 - valj corpus callosum;
11 - spodnji vzdolžni nosilec; 12 - subkalcificiran snop (čelno-okcipitalni spodnji snop); 13 - trezor; 14 - temporalni reženj; 15 - kavelj gyrusa hipokampusa; 16 - snopi kavljev (fasciculus uncinatus).
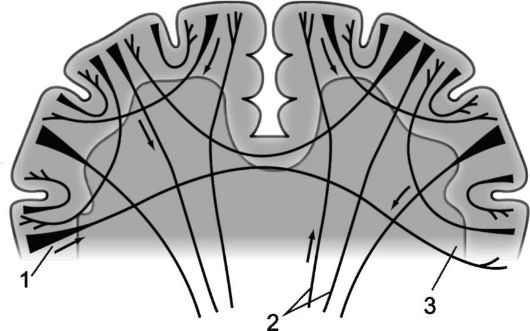
riž. 14.3.Mieloarhitektonika možganskih hemisfer.
1 - projekcijska vlakna; 2 - komisuralna vlakna; 3 - asociativna vlakna.
14.4.1. Zgradba vohalnega sistema
Telesa prvih nevronov vohalnega sistema se nahajajo v sluznici nos, predvsem zgornji del nosnega septuma in zgornji nosni prehod. Vohalne celice so bipolarne. Njihovi dendriti pridejo na površino sluznice in se tu končajo s specifičnimi receptorji in aksoni so združeni v ti vohalne nitke (filiolfactorii), katerih število na vsaki strani je približno dvajset. Takšna snop vohalnih filamentov in sestavlja I kranialni ali vohalni živec(slika 14.4). Te niti prehajajo v sprednjo (olfaktorno, vohalno) lobanjsko foso skozi etmoidno kost in se končajo pri celice, ki se nahajajo tukaj vohalne čebulice. Olfaktorni bulbusi in proksimalni vohalni trakti so pravzaprav posledica izrastkov snovi velikih možganov, ki so nastali v procesu ontogeneze in predstavljajo z njim povezane strukture.
Vohalne čebulice vsebujejo celice, ki so telesa drugih nevronov. vohalne poti, katere aksoni tvorijo vohalne poti (vohalni trakti), ki se nahajajo pod vohalnimi utori, bočno od neposrednih vijug, ki se nahajajo na bazalni površini čelni režnji. Vohalni trakti so usmerjeni nazaj v subkortikalne olfaktorne centre. Ko se približamo sprednji perforirani plošči, so vlakna vohalnega trakta razdeljena na medialne in lateralne snope, ki na vsaki strani tvorijo vohalni trikotnik. Kasneje so ta vlakna primerna telesom tretjih nevronov vohalnega analizatorja, ki se nahajajo
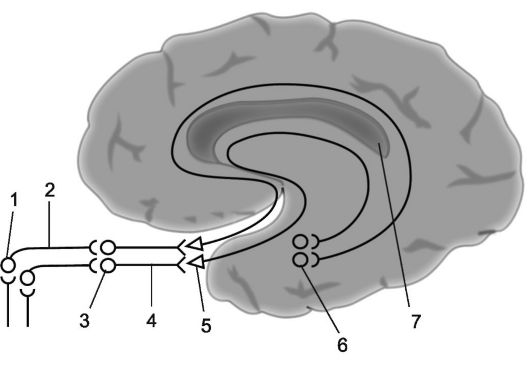
riž. 14.4.Vohalni analizator.
1 - vohalne celice; 2 - vohalne niti (skupaj sestavljajo vohalne živce); 3 - vohalne čebulice; 4 - vohalni trakti; 5 - vohalni trikotniki; 6 - parahipokampalni girus; 7 - projekcijsko območje olfaktornega analizatorja (poenostavljen diagram).
v perialmondastih in subkalozalnih območjih, v jedrih prozornega septuma, ki se nahajajo spredaj od sprednje komisure. Sprednja komisura povezuje obe vohalni regiji in zagotavlja tudi njuno povezavo z limbičnim sistemom možganov. Del aksonov tretjih nevronov vohalnega analizatorja, ki poteka skozi sprednjo komisuro možganov, se križa.
Aksoni tretjih nevronov vohalni analizator, ki se nahaja v subkortikalnih vohalnih centrih, usmerjen proti filogenetsko stara skorja mediobazalna površina temporalnega režnja (do piriformnega in parahipokampalnega girusa ter do kljuke), kjer se nahaja projekcijsko vohalno območje, ali kortikalni konec olfaktornega analizatorja (polje 28 po Brodmannu).
Vohalni sistem je torej edini senzorični sistem, v katerem specifični impulzi na poti od receptorjev do skorje obidejo talamus. vendar Vohalni sistem ima še posebej izrazite povezave z limbičnimi strukturami možganov, informacije, ki jih prejme, pomembno vplivajo na stanje čustvene sfere in funkcije avtonomnega živčnega sistema. Vonji so lahko prijetni in neprijetni, vplivajo na apetit, razpoloženje, lahko povzročijo različne vegetativne reakcije, zlasti slabost, bruhanje.
14.4.2. Preiskava voha in pomen njegovih motenj za lokalno diagnostiko
Pri preučevanju stanja vonja je treba ugotoviti, ali bolnik čuti vonjave, ali so ti občutki enaki na obeh straneh, ali bolnik razlikuje naravo zaznanih vonjav, ali ima vohalne halucinacije - paroksizmične občutke vonja. ki jih ni v okolju.
Za preučevanje občutka za vonj se uporabljajo dišavne snovi, katerih vonj ni oster (ostri vonji lahko povzročijo draženje receptorjev trigeminalnega živca, ki se nahajajo v nosni sluznici) in je pacientu znan (sicer ga je težko prepoznati). perverznost vonja). Voh preverjamo na vsaki strani posebej, medtem ko mora biti druga nosnica zaprta. Uporabite lahko posebej pripravljene komplete šibkih raztopin dišečih snovi (meta, katran, kafra itd.), V praktičnem delu pa lahko uporabite tudi improvizirana sredstva ( rženi kruh, milo, banana itd.).
Zmanjšan občutek za vonj - hiposmija, pomanjkanje vonja - anosmija, povečan občutek za vonj - hiperosmija, perverznost vonjav disosmija, občutek vonja v odsotnosti dražljaja - parozmija, subjektivni občutek slab vonj, ki dejansko obstaja in je posledica organske patologije v nazofarinksu - kakosmija, vonjave, ki v resnici ne obstajajo, ki jih bolnik občuti paroksizmalno - vohalne halucinacije - so pogosteje vohalna aura temporalne epilepsije, ki je lahko posledica različnih vzrokov, zlasti tumorja temporalnega režnja.
Hipozmija ali anosmija na obeh straneh je običajno posledica poškodbe nosne sluznice zaradi akutnega katarja, gripe, alergijskega rinitisa, atrofije sluznice.
nosu zaradi kroničnega rinitisa in dolgotrajna uporaba vazokonstrikcijske kapljice za nos. Kronični rinitis z atrofijo nosne sluznice (atrofični rinitis), Sjögrenova bolezen obsoja osebo na vztrajno anosmijo. Dvostransko hiposmijo lahko povzročijo hipotiroidizem, diabetes mellitus, hipogonadizem, odpoved ledvic, dolgotrajen stik s težkimi kovinami, formaldehidom itd.
vendar enostranska hiposmija ali anosmija je pogosto posledica intrakranialnega tumorja, pogosteje meningioma sprednje lobanjske (vohalne) jame, ki predstavlja do 10% intrakranialnih meningiomov, pa tudi nekatere glialne tumorje čelnega režnja. Vohalne motnje se pojavijo kot posledica stiskanja vohalne poti na strani patološkega žarišča in so lahko edini žariščni simptom bolezni za določen čas. Tumorje lahko vizualiziramo s CT ali MRI skeniranjem. Ko se meningiomi vohalne jame povečajo, se praviloma razvijejo duševne motnje značilnost frontalnega sindroma (glej poglavje 15).
Enostranska poškodba delov vohalnega analizatorja, ki se nahajajo nad njegovimi subkortikalnimi središči, zaradi nepopolne prekrivanja poti na ravni sprednje cerebralne komisure običajno ne vodi do bistvenega zmanjšanja občutka za vonj. Draženje patološkega procesa skorje mediobazalnih delov temporalnega režnja, predvsem parahipokampalnega girusa in njegovega kljuka, lahko povzroči paroksizmalni pojav vohalne halucinacije. Pacient nenadoma brez razloga začne vonjati, pogosto neprijetne narave (vonj po zažganem, gnilem, gnilem, zažganem itd.). Vohalne halucinacije v prisotnosti epileptogenega žarišča v mediobazalnih regijah temporalnega režnja možganov je lahko manifestacija avre epileptičnega napada. Poraz proksimalnega dela, zlasti skorje konca vohalnih analizatorja, lahko povzroči zmerno dvostransko (več na nasprotni strani) hiposmijo in oslabljeno sposobnost prepoznavanja in razlikovanja vonjav (vohalna agnozija). Zadnja oblika vohalne motnje, ki se kaže v starosti, je najverjetneje povezana s kršitvijo funkcije skorje zaradi atrofičnih procesov v njeni projekcijski vohalni coni.
14.5. LIMBIČNO-RETIKULARNI KOMPLEKS
Leta 1878 P. Broca(Broca P., 1824-1880) pod imenom "veliki robni ali limbični reženj" (iz lat. limbus - rob) združuje hipokampus in cingularni gyrus, medsebojno povezani s pomočjo isthmusa cingulate gyrus, ki se nahaja nad grebenom corpus callosum.
Leta 1937 D. Papets(Papez J.) je na podlagi eksperimentalnih podatkov podal utemeljen ugovor proti prej obstoječemu konceptu vpletenosti mediobazalnih struktur možganskih hemisfer predvsem v zagotavljanje vonja. On predlagal, da je morfološka osnova glavni del mediobazalnih delov možganske hemisfere, takrat imenovan vohalni možgani (rhinencephalon), ki mu pripada limbični reženj. živčni mehanizem čustveno vedenje, in jih združil pod imenom"čustveni krog" ki vključuje hipotalamus,
anteriorna jedra talamusa, cingularni girus, hipokampus in njihove povezave. Od takrat te strukture imenujejo tudi fiziologi okoli Papetza.
koncept "visceralni možgani" predlagal P.D. McLean (1949), s čimer označuje kompleksno anatomsko-fiziološko združbo, ki se od leta 1952 imenuje "limbični sistem". Kasneje se je izkazalo, da je limbični sistem vključen v izvajanje različnih funkcij, zdaj pa je večina, vključno s cingulatnim in hipokampalnim (parahipokampalnim) girusom, običajno združena v limbično regijo, ki ima številne povezave s strukturami retikularna tvorba, ki jo sestavlja limbično-retikularni kompleks, ki zagotavlja širok spekter fizioloških in psiholoških procesov.
Trenutno do limbični reženj običajno je pripisati elemente stare skorje (arhiokorteks), ki pokriva zobni girus in hipokampalni girus; starodavna skorja (paleokorteks) sprednjega hipokampusa; kot tudi srednja ali vmesna skorja (mezokorteks) cingularnega gyrusa. Izraz "limbični sistem" vključuje komponente limbičnega režnja in sorodne strukture - entorhinalne (zasedajo večina parahipokampalni girus) in septalni predel, kot tudi kompleks amigdale in mastoidno telo (Duus P., 1995).
Mastoidno telo povezuje strukture tega sistema s srednjimi možgani in z retikularna tvorba. Impulzi, ki izvirajo iz limbičnega sistema, se lahko prenašajo skozi anteriorno jedro talamusa do cingularnega girusa in do neokorteksa po poteh, ki jih tvorijo asociativna vlakna. Impulzi, ki izvirajo iz hipotalamusa, lahko dosežejo orbitofrontalni korteks in medialno dorzalno jedro talamusa.
Številne neposredne in povratne povezave zagotavljajo medsebojno povezanost in soodvisnost limbičnih struktur in številnih tvorb diencefalona in ustnih delov možganskega debla (nespecifična jedra talamusa, hipotalamusa, putamena, frenuluma, retikularne formacije možganskega debla), pa tudi s subkortikalno. jedrih (pallidus, putamen, caudatus nucleus ) in z neokorteksom možganskih hemisfer, predvsem s skorjo temporalnega in čelnega režnja.
Kljub filogenetskim, morfološkim in citoarhitektonskim razlikam številne omenjene strukture (limbični predel, centralne in medialne strukture talamusa, hipotalamus, retikularna formacija možganskega debla) običajno uvrščamo v t.i. limbično-retikularni kompleks, ki deluje kot območje integracije številnih funkcij, ki zagotavlja organizacijo polimodalnih, celostnih reakcij telesa na različne vplive, kar je še posebej izrazito v stresnih situacijah.
Strukture limbično-retikularnega kompleksa imajo veliko število vhodi izhodi, skozi katere začaranih krogih iz številnih aferentnih in eferentnih povezav, ki zagotavljajo skupno delovanje formacij, vključenih v ta kompleks in njihova interakcija z vsemi deli možganov, vključno z možgansko skorjo.
V strukturah limbično-retikularnega kompleksa obstaja konvergenca občutljivih impulzov, ki se pojavljajo v intero- in eksteroreceptorjih, vključno z receptorskimi polji čutnih organov. Na tej podlagi v limbično-retikularnem kompleksu primarna sinteza informacij o stanju notranjega okolja telesa, pa tudi o dejavnikih zunanjega okolja, ki vplivajo na telo, in se oblikujejo osnovne potrebe, biološke motivacije in spremljajoča čustva.
Limbično-retikularni kompleks določa stanje čustvene sfere, sodeluje pri uravnavanju vegetativno-visceralnih odnosov, katerih cilj je vzdrževanje relativne konstantnosti notranjega okolja (homeostaza), pa tudi oskrba z energijo in korelacija motoričnih dejanj. Od njegovega stanja je odvisna stopnja zavesti, možnost avtomatiziranih gibov, aktivnost motoričnih in duševnih funkcij, govor, pozornost, sposobnost orientacije, spomin, sprememba budnosti in spanja.
Poškodbe struktur limbično-retikularnega kompleksa lahko spremljajo različne klinični simptomi: izrazite spremembe v čustveni sferi trajne in paroksizmalne narave, anoreksija ali bulimija, spolne motnje, motnje spomina, še posebej znaki Korsakoffovega sindroma, pri kateri pacient izgubi sposobnost spominjanja trenutnih dogodkov (aktualne dogodke zadrži v spominu največ 2 minuti), avtonomno-endokrine motnje, motnje spanja, psihosenzorične motnje v obliki iluzij in halucinacij, spremembe zavesti, klinične manifestacije akinetičnega mutizma, epileptični napadi.
Do danes je bilo izvedenih veliko število študij o preučevanju morfologije, anatomskih razmerij, delovanja limbične regije in drugih struktur, vključenih v limbično-retikularni kompleks, vendar so fiziologija in značilnosti klinične slike njegove lezije danes je še vedno treba v veliki meri razjasniti. Večino informacij o njegovi funkciji, zlasti funkcije parahipokampalne regije, pridobljeno v poskusih na živalih metode draženja, ekstirpacije ali stereotaksije. Pridobljeno na ta način rezultati zahtevajo previdnost pri ekstrapolaciji na ljudi. Posebej pomembna so klinična opazovanja bolnikov z lezijami mediobazalnih delov možganske hemisfere.
V 50-60 letih XX stoletja. med razvojem psihokirurgije so poročali o zdravljenju bolnikov z neozdravljivimi duševne motnje in sindrom kronične bolečine z bilateralno cingulotomijo (disekcija cingularnega girusa), medtem ko je bila običajno opažena regresija anksioznosti, obsesivnih stanj, psihomotorične agitacije, bolečinski sindromi, ki je bil prepoznan kot dokaz o vpletenosti cingularnega gyrusa v nastanek čustev in bolečine. Istočasno je bicingulotomija povzročila globoke osebnostne motnje, dezorientacijo, zmanjšanje kritičnosti stanja in evforijo.
Analiza 80 potrjenih kliničnih primerov lezij hipokampusa na podlagi Nevrokirurškega inštituta Ruske akademije medicinskih znanosti je podana v monografiji N.N. Bragina (1974). Avtor pride do zaključka, da temporalni mediobazalni sindrom vključuje viscerovegetativne, motorične in duševne motnje, ki se običajno kažejo v kompleksu. Vsa raznolikost kliničnih manifestacij N.N. Bragin zmanjša na dve glavni večfaktorski različici patologije s prevlado "dražilnih" in "inhibicijskih" pojavov.
Prva od teh vključuje čustvene motnje ki ga spremlja motorična anksioznost (povečana razdražljivost, besedičnost, sitnost, občutek notranje tesnobe), paroksizmi strahu, vitalna tesnoba, različne viscerovegetativne motnje (spremembe pulza, dihanja, prebavne motnje, zvišana telesna temperatura, povečano potenje itd.). Pri teh bolnikih so se v ozadju stalnega motoričnega nemira pogosto pojavili napadi motoričnega vzbujanja.
niya. Za EEG te skupine bolnikov so bile značilne blage možganske spremembe v smeri integracije (pospešen in poudarjen alfa ritem, difuzna beta nihanja). Ponavljajoči se aferentni dražljaji so izzvali jasne odzive EEG, ki za razliko od običajnih niso zbledeli, ko so bili dražljaji večkrat predstavljeni.
Za drugo ("inhibitorno") različico mediobazalnega sindroma so značilne čustvene motnje v obliki depresije z motorično zaostalostjo (depresivno razpoloženje, osiromašenje in upočasnitev tempa). miselni procesi, spremembe v motiliteti, ki spominjajo na vrsto akinetično-rigidnega sindroma. Viscerovegetativni paroksizmi, opaženi v prvi skupini, so manj značilni. Za EEG bolnikov v tej skupini so značilne cerebralne spremembe, ki se kažejo v prevladi počasnih oblik aktivnosti (nepravilen, počasen alfa ritem, skupine theta nihanj, difuzni delta valovi). Močno zmanjšanje reaktivnosti EEG je pritegnilo pozornost.
Med tema dvema skrajnima možnostma so bile vmesne s prehodnimi in mešanimi kombinacijami. posamezne simptome. Tako so za nekatere od njih značilni razmeroma šibki znaki agitirane depresije s povečano motorična aktivnost in utrujenost, s prevlado senestopatskih občutkov, suma, ki pri nekaterih bolnikih doseže paranoidna stanja, hipohondrijski delirij. Drugo vmesno skupino je odlikovala izjemna intenzivnost depresivnih simptomov glede na bolnikovo okorelost.
Ti podatki nam omogočajo, da govorimo o dvojnem (aktivacijskem in zaviralnem) vplivu hipokampusa in drugih struktur limbične regije na vedenjske reakcije, čustva, duševno stanje in bioelektrično aktivnost korteksa. Trenutno zapleteno klinični sindromi te vrste ne bi smeli obravnavati kot primarno žariščno. Namesto tega jih je treba obravnavati v luči idej o večstopenjskem sistemu organizacije možganske dejavnosti.
S.B. Buklina (1997) je navedel podatke iz raziskave 41 bolnikov z arteriovenskimi malformacijami v območju cingularnega gyrusa. Pred operacijo je imelo 38 bolnikov v ospredju motnje spomina, pet jih je imelo znake Korsakoffovega sindroma, pri treh bolnikih se je Korsakoffov sindrom pojavil po operaciji, medtem ko je resnost povečanja spominskih okvar korelirala s stopnjo uničenja cingularnega gyrusa. sama, kot tudi z vpletenostjo v patološki proces sosednjih struktur corpus callosum, medtem ko amnezijski sindrom ni bil odvisen od strani lokacije malformacije in njene lokalizacije po dolžini cingulate gyrus.
Glavne značilnosti ugotovljenih amnestičnih sindromov so bile motnje v reprodukciji slušno-govornih dražljajev, kršitve selektivnosti sledi v obliki vključkov in kontaminacij ter nezmožnost ohranjanja pomena pri prenosu zgodbe. Pri večini bolnikov je bila kritičnost ocene njihovega stanja zmanjšana. Avtor je opozoril na podobnost teh motenj z amnestičnimi okvarami pri bolnikih s čelnimi lezijami, kar je mogoče razložiti s prisotnostjo povezav med cingulatnim girusom in čelnim režnjem.
več Razširjeni patološki procesi v limbični regiji povzročajo izrazite motnje vegetativno-visceralnih funkcij.
corpus callosum(corpus callosum)- največja komisura med možganskima hemisferama. Njegovi sprednji deli, zlasti koleno corpus callosum
telo (genu corporis callosi), povezujejo čelne režnjeve, srednje odseke - deblo corpus callosum (truncus corporis callosi)- zagotavljajo komunikacijo med temporalnimi in parietalnimi deli hemisfer, posteriornimi deli, zlasti grebenom corpus callosum (splenium corporis callosi), povežite okcipitalne režnje.
Lezije corpus callosum običajno spremljajo motnje duševnega stanja pacienta. Uničenje njegovega sprednjega dela vodi do razvoja "čelne psihe" (aspontanost, kršitve akcijskega načrta, vedenje, kritičnost, značilnost frontalni kalozni sindrom - akinezija, amimija, aspontanost, astazija-abazija, apraksija, prijemalni refleksi, demenca). Prekinitev povezav med parietalnimi režnji vodi v perverzijo razumevanje "načrti telesa" in pojav apraksije večinoma v levi roki. Disociacija temporalnih režnjev se lahko manifestira kršitev zaznavanja zunanjega okolja, izguba pravilne orientacije v njem (amnestične motnje, konfabulacije, sindrom že videnega). in tako naprej.). Za patološka žarišča v posteriornih delih corpus callosum so pogosto značilni znaki vizualne agnozije.
14.6. ARHITEKTONIKA MOŽGANSKE SKORJE
Struktura možganske skorje je heterogena. Manj zapletena struktura, zgodaj se pojavi v procesu filogeneze starodavno lubje (arhiokorteks) in staro lubje (paleokorteks), povezano večinoma do limbičnega režnja možgani. Večji del možganske skorje (95,6 %) se zaradi poznejšega filogenetskega nastanka imenuje novo lubje (neokorteks) in ima veliko bolj zapleteno večplastno strukturo, a tudi heterogena v svojih različnih conah.
Zaradi arhitektonika skorje je v določeni povezavi z njeno funkcijo, njegovemu preučevanju je bilo posvečeno veliko pozornosti. Eden od ustanoviteljev doktrine citoarhitektonike skorje je bil V.A. Betz (1834-1894) je leta 1874 prvi opisal velike piramidne celice. motorično območje korteks (Betzove celice) in določil principe delitve možganske skorje na glavna področja. V prihodnosti so veliko prispevali k razvoju teorije o strukturi korteksa številni raziskovalci - A. Campbell (A. Cambell), E. Smith (E. Smith), K. Brodmann (K. Brodmann). ), Oscar Vogt in Cecilia Vogt (O. Vogt , S. Vogt). Velike zasluge pri preučevanju arhitektonike skorje pripada osebju Inštituta za možgane Akademije medicinskih znanosti (S.A. Sarkisov, N.I. Filimonov, E.P. Kononova itd.).
Glavna vrsta strukture nove skorje (slika 14.5), s katerim se primerjajo vsi njegovi deli - skorja, sestavljena iz 6 plasti (homotipska skorja, po Brodmanu).
Plast I - molekularna ali conska, najbolj površinska, revna v celicah, njena vlakna imajo smer, večinoma vzporedno s površino skorje.
II sloj - zunanji zrnat. Sestavljen je iz velikega števila gosto razporejenih majhnih zrnc živčne celice.
III sloj - majhne in srednje piramide, najširše. Sestavljen je iz piramidnih celic, katerih velikosti niso enake, kar omogoča razdelitev te plasti na podsloje v večini kortikalnih polj.
IV sloj - notranji zrnat. Sestavljen je iz gosto razporejenih majhnih celic-zrn okrogle in oglate oblike. Ta plast je najbolj spremenljiva
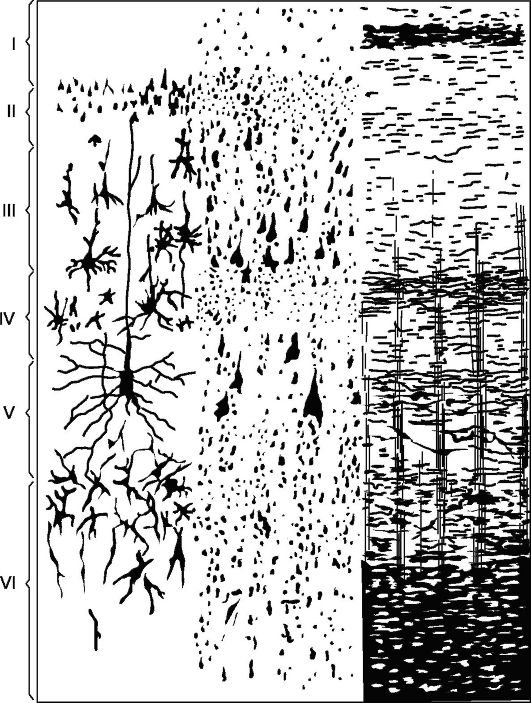
riž. 14.5.Citoarhitektonika in mieloarhitektonika motorične cone možganske skorje.
Levo: I - molekularna plast; II - zunanja zrnata plast; III - plast majhnih in srednjih piramid; IV - notranja zrnata plast; V - plast velikih piramid; VI - plast polimorfnih celic; na desni - elementi mieloarhitektonike.
v nekaterih poljih (npr. polje 17) je razdeljena na podsloje, ponekod se močno stanjša in celo popolnoma izgine.
V plast - velike piramide ali ganglijske. Vsebuje velike piramidne celice. V nekaterih predelih možganov je plast razdeljena na podplasti, v motorični coni pa je sestavljena iz treh podplasti, od katerih je sredina Betzova velikanska piramidna celica, ki v premeru doseže 120 mikronov.
VI sloj - polimorfne ali multiformne celice. Sestavljen je predvsem iz trikotnih vretenastih celic.
Struktura možganske skorje ima veliko variacij zaradi spreminjanja debeline posameznih plasti, tanjšanja ali izginjanja oz.
nasprotno, zgostitev in delitev na podsloje nekaterih od njih (heterotipne cone, po Brodmanu).
Skorja vsake možganske hemisfere je razdeljena na več regij: okcipitalno, zgornjo in spodnjo parietalno, postcentralno, osrednjo vijugo, precentralno, frontalno, temporalno, limbično, otoško. Vsak od njih v skladu z lastnostmi razdeljen na več polj, poleg tega ima vsako polje svojo konvencionalno vrstno oznako (slika 14.6).
Preučevanje arhitektonike možganske skorje, skupaj s fiziološkimi, vključno z elektrofiziološkimi študijami in kliničnimi opazovanji, je veliko prispevalo k rešitvi problema porazdelitve funkcij v skorji.
14.7. PROJEKCIJSKA IN ASOCIACIONA POLJA CORTUSA
V procesu razvoja doktrine o vlogi možganske skorje in njenih posameznih delov pri opravljanju določenih funkcij so obstajala različna, včasih nasprotna stališča. Tako je obstajalo mnenje o strogo lokalni zastopanosti v možganski skorji vseh človeških sposobnosti in funkcij, do najbolj zapletenih, duševnih. (lokalizacizem, psihomorfologija). Nasprotoval mu je drugo mnenje o absolutni funkcionalni enakovrednosti vseh delov možganske skorje. (ekvipotencializem).
Pomemben prispevek k teoriji o lokalizaciji funkcij v možganski skorji je prispeval I.P. Pavlov (1848-1936). Izločil je projekcijske cone korteksa (kortikalni konci analizatorjev določene vrste občutljivost) in asociativne cone, ki se nahajajo med njimi, preučevali procese inhibicije in vzbujanja v možganih, njihov vpliv na funkcionalno stanje možganska skorja. Delitev skorje na projekcijske in asociativne cone prispeva k razumevanju organizacije dela možganske skorje in se upravičuje pri reševanju praktične naloge, zlasti v lokalni diagnostiki.
projekcijske cone zagotavljajo predvsem preprosta specifična fiziološka dejanja, predvsem zaznavanje občutkov določene modalnosti. Projekcijske poti, ki se jim približujejo, povezujejo ta območja z receptorskimi ozemlji na obrobju, ki so v funkcionalni korespondenci z njimi. Primeri projekcijskih kortikalnih con so območje posteriornega osrednjega gyrusa, ki je že opisano v prejšnjih poglavjih (območje splošne vrste občutljivost) ali predel utora na ostrogu, ki se nahaja na medialni strani okcipitalnega režnja (projekcijsko vidno območje).
Asociacijske cone korteks nima neposrednih povezav s periferijo. Nahajajo se med projekcijskimi conami in imajo številne asociativne povezave s temi projekcijskimi conami in z drugimi asociativnimi conami. Funkcija asociacijskih con je izvajanje višja analiza in sintezo številnih osnovnih in kompleksnejših komponent. Tu gre v bistvu za razumevanje informacij, ki vstopajo v možgane, za oblikovanje idej in konceptov.
G.I. Poljakov je leta 1969 na podlagi primerjave arhitektonike možganske skorje človeka in nekaterih živali ugotovil, da asociativni
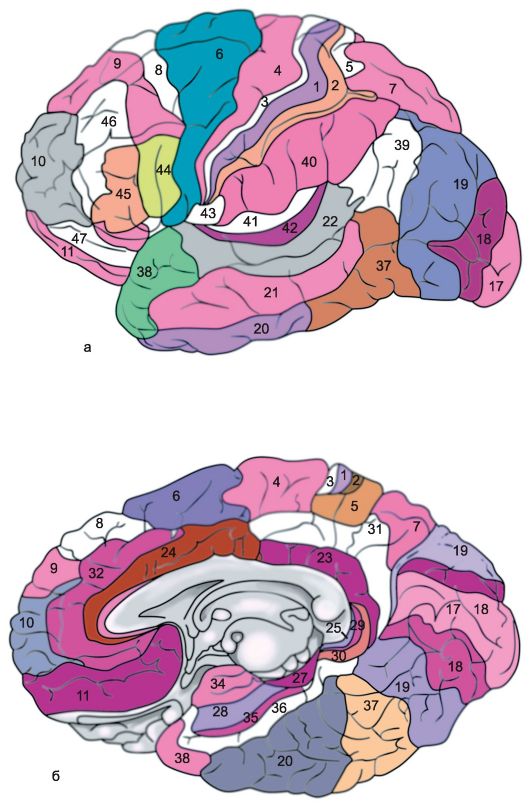
riž. 14.6.Arhitektonska polja možganske skorje [po Brodmanu]. a - zunanja površina; b - medialna površina.
cone v človeški možganski skorji so 50%, v skorji višjih (humanoidnih) opic - 20%, pri nižjih opicah je ta številka 10% (slika 14.7). Med asociacijskimi področji skorje človeških možganov je isti avtor predlagal izolacijo sekundarno in terciarno področje. Sekundarna asociativna polja so poleg projekcijskih. Izvajajo analizo in sintezo elementarnih občutkov, ki še vedno ohranjajo določeno usmerjenost.
Terciarna asociativna polja se nahajajo predvsem med sekundarnimi in prekrivajo območja sosednjih ozemelj. Povezani so predvsem z analitično aktivnostjo korteksa, ki zagotavlja najvišje duševne funkcije, ki so lastne človeku v njihovih najbolj zapletenih intelektualnih in govornih manifestacijah. Funkcionalna zrelost terciarne as-
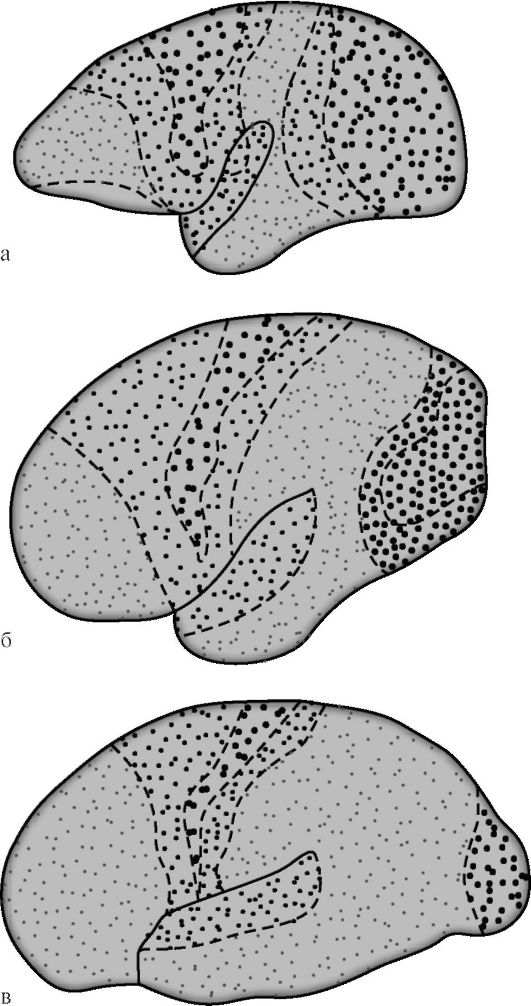
riž. 14.7. Diferenciacija projekcijskih in asociativnih področij možganske skorje med evolucijo primatov [po G.I. Polyakov]. a - možgani spodnje opice; b - možgani višje opice; c - človeški možgani. Velike pike označujejo projekcijska območja, majhne pike - asociativne. Pri nižjih opicah asociativne cone zavzemajo 10% površine korteksa, pri višjih - 20%, pri ljudeh - 50%.
socialnih polj možganske skorje se pojavi najpozneje in le v ugodnem družbenem okolju. Za razliko od drugih kortikalnih polj je za terciarna polja desne in leve hemisfere značilna izrazita funkcionalna asimetrija.
14.8. AKTUALNA DIAGNOSTIKA LEZIJ MOŽGANSKE SKORJE
14.8.1. Manifestacije poškodb projekcijskih območij možganske skorje
V skorji vsake možganske hemisfere, za osrednjim girusom, je 6 projekcijskih con.
1. V sprednjem delu parietalnega režnja, v predelu posteriornega centralnega girusa (citoarhitektonska polja 1, 2, 3) nahaja projekcijsko območje splošnih vrst občutljivosti(slika 14.4). Območja skorje, ki se nahajajo tukaj, sprejemajo občutljive impulze, ki prihajajo po projekcijskih poteh splošnih vrst občutljivosti iz receptorskega aparata nasprotne polovice telesa. Višje kot je območje te projekcijske cone skorje, nižje locirani deli nasprotne polovice telesa ima projekcijske povezave. Deli telesa z obsežnim sprejemom (jezik, dlančna površina roke) ustrezajo neustrezno velikim delom območja projekcijskih con, drugi deli telesa (proksimalni udi, trup) pa imajo majhno površino skorje. reprezentanca.
Draženje s patološkim procesom kortikalne cone splošnih tipov občutljivosti vodi do napada parestezije v delih telesa, ki ustrezajo razdraženim območjem možganske skorje (občutljivi Jacksonov napad), ki se lahko spremeni v sekundarno generalizirano paroksizem. Poraz skorje konca analizatorja splošne vrste občutljivosti lahko povzroči razvoj hipalgezije ali anestezije v ustreznem območju nasprotne polovice telesa, medtem ko je mesto hipestezije ali anestezije lahko vertikalno cirkulacijsko ali radikularno- segmentni tip. V prvem primeru se motnja občutljivosti manifestira na nasprotni strani patološkega žarišča v predelu ustnic, palca ali distalnega dela okončine s krožno obrobo, včasih kot nogavica ali rokavica. V drugem primeru je območje motenj občutljivosti v obliki traku in se nahaja vzdolž notranjega ali zunanjega roba roke ali noge; to je razloženo z dejstvom, da je notranja stran okončin predstavljena v sprednji in zunanja stran - v zadnjih delih projekcijskega območja analizatorja splošnih vrst občutljivosti.
2. Območje vizualne projekcije nahaja v skorji medialne površine okcipitalnega režnja v predelu žleba ostroga (polje 17). V tem polju je stratifikacija IV (notranje zrnate) plasti skorje s snopom mielinskih vlaken v dve podplasti. Ločeni odseki polja 17 sprejemajo impulze iz določenih odsekov istoimenskih polovic mrežnice obeh očes; medtem ko impulzi, ki prihajajo iz spodnjih delov istoimenskih polovic mrežnice, dosežejo
lubje spodnje ustnice žleba za ostroge in impulzi, ki prihajajo iz zgornji deli mrežnice, se pošljejo v skorjo njene zgornje ustnice.
Poraz patološkega procesa vidne projekcijske cone vodi do pojava na nasprotni strani kvadranta ali popolne homonimne hemianopije na strani, ki je nasprotna patološkemu žarišču. Dvostranska lezija kortikalnih polj 17 ali projekcijskih polj, ki gredo do njih vidne poti lahko povzroči popolno slepoto. Draženje skorje vidnega projekcijskega območja lahko povzroči pojav vidnih halucinacij v obliki fotopsij v ustreznih delih nasprotnih polovic vidnih polj.
3. Območje projekcije sluha nahaja v skorji Heschlovih vijug na spodnja ustnica stranska (silvijanska) brazda (polji 41 in 42), ki so pravzaprav del zgornje temporalne vijuge. Draženje tega področja skorje lahko povzroči pojav slušnih halucinacij (napadi občutka hrupa, zvonjenja, žvižganja, brenčanja itd.). Uničenje območja slušne projekcije na eni strani lahko povzroči nekaj izgube sluha v obeh ušesih, v večji meri na nasprotni strani glede na patološko žarišče.
4 in 5. Vohalne in okusne projekcijske cone so na medialni površini obokanega gyrusa (limbična regija) možganov. Prvi od njih se nahaja v parahipokampalnem girusu (polje 28). Projekcijska cona okusa je običajno lokalizirana v skorji operkularnega območja (polje 43). Draženje projekcijskih con vonja in okusa lahko povzroči njihovo perverzijo ali vodi do razvoja ustreznih vohalnih in okusnih halucinacij. Enostranska izguba funkcije projekcijskih con vonja in okusa lahko povzroči rahlo zmanjšanje vonja oziroma okusa na obeh straneh. Dvostransko uničenje kortikalnih koncev istih analizatorjev se kaže v odsotnosti vonja in okusa na obeh straneh.
6. Vestibularna projekcijska cona. Njegova lokalizacija ni določena. Hkrati je znano, da ima vestibularni aparat številne anatomske in funkcionalne povezave. Možno je, da lokalizacija zastopanosti vestibularnega sistema v skorji še ni pojasnjena, ker je polifokalna. N.S. Blagoveshchenskaya (1981) meni, da so v možganski skorji cone vestibularne projekcije predstavljene z več medsebojno povezanimi anatomskimi in funkcionalnimi kompleksi, ki se nahajajo v polju 8, na stičišču čelnega, časovnega in parietalni reženj in v območju osrednjih konvolucij se domneva, da vsak od teh odsekov korteksa opravlja svoje funkcije. Polje 8 je poljubno središče pogleda, njegovo draženje povzroči obračanje pogleda v nasprotni smeri od patološkega žarišča, spremembe v ritmu in naravi eksperimentalnega nistagmusa, zlasti kmalu po epileptičnem napadu. V skorji temporalnega režnja so strukture, katerih draženje povzroča vrtoglavico, kar se kaže zlasti pri epilepsiji temporalnega režnja; poraz področij zastopanja vestibularnih struktur v skorji osrednjega gyrusa vpliva na stanje tonusa progastih mišic. Klinična opazovanja kažejo, da jedrsko-kortikalne vestibularne poti naredijo delno križanje.
Treba je poudariti, da so lahko znaki draženja naštetih projekcijskih območij manifestacija avre epileptičnega napada, ki ustreza naravi.
I.P. Pavlov je menil, da je mogoče upoštevati skorjo precentralnega girusa, ki vpliva na motorične funkcije in mišični tonus pretežno nasprotne polovice telesa, s katero je povezan predvsem s kortikalno-jedrnimi in kortikalno-spinalnimi (piramidnimi) potmi, kot projekcijsko cono ti motorični analizator. To območje zaseda najprej polje 4, na katerem je v obrnjeni obliki projicirana nasprotna polovica telesa. To polje vsebuje večino velikanskih piramidnih celic (Betzovih celic), katerih aksoni sestavljajo 2-2,5% vseh vlaken piramidne poti, pa tudi srednje in majhne piramidne celice, ki skupaj z aksoni istega celice, ki se nahajajo v bližini polja 4, obsežnejšega polja 6, sodelujejo pri izvajanju monosinaptičnih in polisinaptičnih kortikalno-mišičnih povezav. Monosinaptične povezave zagotavljajo predvsem hitre in natančne ciljne akcije, odvisno od kontrakcij posameznika progasta mišice.
Poškodba spodnjih delov motorične cone običajno vodi do razvoja na nasprotni strani brahiofacialni (obrazna rama) sindrom ali lingvofaciobrahialni sindrom, ki ga pogosto opazimo pri bolnikih z moteno cerebralno cirkulacijo v bazenu srednje možganske arterije, medtem ko kombinirana pareza mišic obraza, jezika in roke, zlasti rame v osrednjem tipu.
Draženje skorje motorične cone (polja 4 in 6) vodi do pojava konvulzij v mišicah ali mišičnih skupinah, ki so projicirane na to cono. Pogosteje gre za lokalne konvulzije tipa Jacksonove epilepsije, ki se lahko spremenijo v sekundarni generalizirani epileptični napad.
14.8.2. Manifestacije poškodbe asociativnih polj možganske skorje
Med projekcijskimi conami korteksa so asociacijska polja. Prejemajo impulze predvsem iz celic projekcijskih con korteksa. V asociativnih poljih poteka analiza in sinteza informacij, ki so bile podvržene primarna obdelava v projekcijskih poljih. Asociativne cone skorje zgornjega parietalnega lobula zagotavljajo sintezo elementarnih občutkov, v zvezi s tem tako kompleksne vrste občutljivosti, kot so občutek lokalizacije, občutek teže, dvodimenzionalno-prostorski občutek, pa tudi kompleksen tu se oblikujejo kinestetični občutki.
V predelu interparietalnega sulkusa je asociativna cona, ki zagotavlja sintezo občutkov, ki izhajajo iz delov lastnega telesa. Poškodba tega predela korteksa vodi do avtopagnozija, tiste. do neprepoznavanja ali ignoriranja delov lastnega telesa oz psevdomelija občutek dodatne roke ali noge in anozognozija - nezavedanje telesne okvare, ki je nastala v povezavi z boleznijo (na primer paraliza ali pareza okončine). Običajno se vse vrste avtopagnozije in anozognozije pojavijo, ko se patološki proces nahaja na desni.
Poraz spodnjega parietalnega lobula se lahko kaže v motnji sinteze osnovnih občutkov ali nezmožnosti primerjave sintetiziranih kompleksnih občutkov z "nekoč v zaznavanju podobnih"
na enak način, na podlagi rezultatov katerega pride do priznanja «(V.M. Bekhterev). To se kaže s kršitvijo dvodimenzionalnega prostorskega občutka (grafestezija) in tridimenzionalnega prostorskega občutka (stereognozija) - astereognoza.
V primeru poškodbe premotoričnih con čelnega režnja (polja 6, 8, 44) se običajno pojavi frontalna ataksija, pri kateri je motena sinteza aferentnih impulzov (kinestetična aferentacija), ki signalizirajo položaj delov telesa v prostoru, ki spremembe med opravljenimi gibi.
V primeru kršitve funkcije skorje sprednjih delov čelnega režnja, ki je povezan z nasprotno hemisfero malih možganov (čelno-mostno-cerebelarne povezave), se pojavijo statokinetične motnje na nasprotni strani patološkega žarišča. (frontalna ataksija). Posebej izrazite so kršitve pozno nastalih oblik statokinetike - pokončno stanje in pokončna hoja. Posledično ima bolnik negotovost, nestabilnost hoje. Med hojo se njegovo telo nagne nazaj. (Hennerjev znak) stopala postavi v ravno črto (lisičja hoja) včasih pri hoji pride do "prepletanja" nog. Pri nekaterih bolnikih s poškodbo sprednjih delov čelnih režnjev se razvije poseben pojav: v odsotnosti paralize in pareze ter ohranjene sposobnosti gibanja nog v celoti bolniki ne morejo stati. (astazija) in hoditi (abazija).
Za poraz asociativnih con korteksa je pogosto značilen razvoj kliničnih manifestacij kršitve višjih duševnih funkcij (glej poglavje 15).
Splošni pregled zgradbe možganskih hemisfer
Možganske hemisfere so najmasivnejši del možganov. Pokrivajo male možgane in možgansko deblo. Možganske hemisfere predstavljajo približno 78% celotne mase možganov. V procesu ontogenetskega razvoja organizma se možganske hemisfere razvijejo iz medule nevralne cevi, zato ta del možganov imenujemo tudi terminalni možgani.
Možganski hemisferi sta vzdolž srednje črte razdeljeni z globoko navpično razpoko na desno in levo hemisfero.
V globini srednjega dela sta obe hemisferi povezani z velikim oprijemom - corpus callosum. V vsaki polobli se razlikujejo režnji; frontalni, parietalni, temporalni, okcipitalni in insula.
Režnji možganskih hemisfer so med seboj ločeni z globokimi brazdami. Najpomembnejše so tri globoke brazde: osrednja (Rolandova), ki ločuje čelni reženj od temenskega, stranska (Sylvian), ki ločuje temporalni reženj od temenskega, parietalno-okcipitalna, ki ločuje parietalni reženj od okcipitalnega na notranji površini polobla.
Vsaka polobla ima zgornjo stransko (konveksno), spodnjo in notranjo površino.
Vsak reženj hemisfere ima možganske vijuge, ki so med seboj ločene z brazdami. Od zgoraj je hemisfera prekrita z lubjem - tanko plastjo sive snovi, ki jo sestavljajo živčne celice.
Možganska skorja je evolucijsko najmlajša tvorba centralnega živčnega sistema. Pri človeku doseže najvišji razvoj. Možganska skorja je zelo pomembna pri uravnavanju vitalne aktivnosti telesa, pri izvajanju kompleksnih oblik vedenja in oblikovanju nevropsihičnih funkcij.
Pod skorjo je bela snov hemisfer, sestavljena je iz procesov živčnih celic - prevodnikov. Zaradi nastajanja možganskih vijug se skupna površina možganske skorje znatno poveča. celotna površina hemisferna skorja je 1200 cm 2, pri čemer se 2/3 njene površine nahaja v globinah brazd, 1/3 pa na vidni površini hemisfer. Vsak del možganov ima drugačen funkcionalni pomen.
a - zgornja stranska površina: 1 - spodnji čelni girus; 2 - srednji čelni girus; 3 - zgornji čelni girus; 4 - sprednji osrednji girus; 5 - osrednja (Rolandova) brazda; 6 - zadnji osrednji girus; 7 - zgornji parietalni lobule; 8 - spodnji parietalni lobule; 9 - supramarginalna (supramarginalna) brazda; 10 - kotna (kotna) brazda; 11 - parieto-okcipitalni sulkus; 12 - spodnji temporalni girus;
13 - srednji temporalni girus; 14 - zgornji temporalni girus; 15 - stranska (Sylvian) brazda;
b - notranja površina: 1 paracentralni lobule; 2 - osrednja brazda; 3 - cingularni gyrus: 4 - corpus callosum; 5 - parieto-okcipitalni sulkus; 6 - klin; 7 - brazda; 8 - lingularni girus; 9 - girus hipokampusa (parahipokampalni girus).
Struktura čelnega režnja
Čelni reženj zavzema sprednje dele hemisfer. Od parietalnega režnja je ločen s centralnim sulkusom, od temporalnega režnja pa z lateralnim sulkusom. V čelnem režnju so štirje girusi: en navpični - precentralni in trije vodoravni - zgornji, srednji in spodnji čelni girus. Zvitki so med seboj ločeni z brazdami.
Na spodnji površini čelnih režnjev se razlikujeta neposredni in orbitalni girus. Neposredni girus leži med notranjim robom poloble, vohalnim žlebom in zunanjim robom poloble.
V globini vohalne brazde ležita vohalni bulbus in vohalni trakt.
Človeški čelni reženj predstavlja 25–28 % korteksa; povprečna masa čelnega režnja je 450 g.
Delovanje čelnih režnjev povezana z organizacijo prostovoljnih gibov, motoričnih mehanizmov govora, regulacije kompleksnih oblik vedenja, miselnih procesov. Več funkcionalno pomembnih centrov je koncentriranih v vijugah čelnega režnja. Sprednji osrednji girus je "reprezentacija" primarne motorične cone s strogo določeno projekcijo delov telesa. Obraz se "nahaja" v spodnji tretjini gyrusa, roka je v srednji tretjini, noga je v zgornja tretjina. Trup je predstavljen v zadnjih delih zgornjega čelnega gyrusa. Tako je oseba projicirana v sprednjem osrednjem girusu z glavo navzdol in navzdol.
Sprednji osrednji girus, skupaj s sosednjimi posteriornimi in čelnimi vijugami, opravlja zelo funkcionalno pomembno vlogo. Je središče prostovoljnih gibanj. V globinah skorje osrednjega gyrusa, iz tako imenovanih piramidnih celic - osrednjega motoričnega nevrona - se začne glavna motorična pot - piramidni, kortikospinalni, pot. Periferni procesi motoričnih nevronov izhajajo iz skorje, se zberejo v en močan snop, prehajajo skozi osrednjo belo snov hemisfer in vstopijo v možgansko deblo skozi notranjo kapsulo; na koncu možganskega debla se delno križajo (prehajajo z ene strani na drugo) in se nato spustijo v hrbtenjačo. Te veje se končajo v sive snovi hrbtenjača. Tam pridejo v stik s perifernim motoričnim nevronom in nanj prenašajo impulze iz centralnega motoričnega nevrona. Impulzi prostovoljnega gibanja se prenašajo po piramidni poti.
V zadnjih delih zgornjega čelnega gyrusa je tudi ekstrapiramidne središče korteksa, anatomsko in funkcionalno tesno povezana s formacijami tako imenovanega ekstrapiramidnega sistema. Ekstrapiramidni sistem - pogonski sistem za pomoč pri prostovoljnem gibanju. To je sistem "zagotavljanja" poljubnih gibov. Ker je filogenetsko starejši, človeški ekstrapiramidni sistem zagotavlja samodejno regulacijo "naučenih" motoričnih dejanj, ohranjanje splošne mišični tonus, pripravljenost obrobja lokomotivnega sistema za izvajanje gibov, prerazporeditev mišičnega tonusa med gibi. Poleg tega sodeluje pri ohranjanju normalne drže.
motorična področja korteksa se nahajajo predvsem v precentralnem girusu (polja 4 in 6) in paracentralnem lobulu na medialni površini hemisfere. Obstajata primarna in sekundarna področja – polja 4 in 6. Ta polja so motorična, vendar se po svojih značilnostih, po raziskavah Inštituta za možgane, razlikujejo. V primarni motorični skorji(polje 4) so nevroni, ki inervirajo motorične nevrone mišic obraza, trupa in udov.
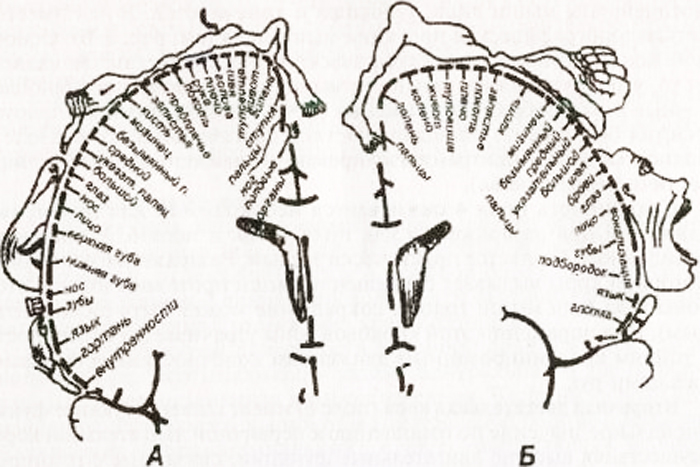
Shema samotematske projekcije splošne občutljivosti in motoričnih funkcij v možganski skorji (po W. Penfieldu):
A - kortikalna projekcija splošne občutljivosti; B - kortikalna projekcija motoričnega sistema. Relativne velikosti organov odražajo območje možganske skorje, iz katerega je mogoče izzvati ustrezne občutke in gibe.
Ima jasno topografsko projekcijo mišic telesa. Glavni vzorec topografske predstavitve je, da regulacija aktivnosti mišic, ki zagotavljajo najbolj natančne in raznolike gibe (govor, pisanje, izrazi obraza), zahteva sodelovanje velikih področij motorične skorje. Polje 4 je v celoti zasedeno s središči izoliranih gibov, polje 6 je zasedeno le delno (podpolje 6a).
Ohranjanje polja 4 se izkaže kot nujno za doseganje gibov pri stimulaciji tako polja 4 kot polja 6. Pri novorojenčku je polje 4 praktično zrelo. Stimulacija primarnega motoričnega korteksa povzroči krčenje mišic nasprotna stran telo (za mišice glave je kontrakcija lahko dvostranska). S porazom te kortikalne cone se izgubi sposobnost finih usklajenih gibov okončin in zlasti prstov.
sekundarni motorični korteks(polje 6) ima prevladujoč funkcionalni pomen v odnosu do primarnega motoričnega korteksa, ki izvaja višje motorične funkcije, povezane z načrtovanjem in usklajevanjem prostovoljnih gibov. Tukaj se počasi povečuje negativno potencial pripravljenosti, ki se pojavi približno 1 s pred začetkom gibanja. Korteks polja 6 prejme večino impulzov iz bazalnih ganglijev in malih možganov in sodeluje pri zapisovanju informacij o kompleksnih gibih.
Draženje skorje polja 6 povzroči kompleksne koordinirane gibe, kot so obračanje glave, oči in trupa v nasprotni smeri, prijazne kontrakcije fleksorjev ali ekstenzorjev na nasprotni strani. Premotorični korteks vsebuje motorične centre, povezane z socialne funkciječlovek: središče pisnega govora v zadnjem delu srednjega čelnega girusa (polje 6), središče Brocinega motoričnega govora v zadnjem delu spodnjega čelnega girusa (polje 44), ki zagotavlja govor, pa tudi glasbeni motor center (polje 45), ki zagotavlja tonaliteto govora, sposobnost petja . Spodnji del polja b (podpolje bora), ki se nahaja v predelu pnevmatike, reagira na električni tok z ritmičnimi žvečilnimi gibi. Nevroni motorične skorje prejemajo aferentne vhode skozi talamus iz mišičnih, sklepnih in kožnih receptorjev, iz bazalnih ganglijev in malih možganov. Glavni eferentni izhod motorične skorje v deblo in hrbtenične motorične centre so piramidne celice plasti V.
V zadnjem delu srednjega čelnega gyrusa je čelni okulomotorni center, ki nadzoruje prijazno, sočasno vrtenje glave in oči (središče vrtenja glave in oči v nasprotni smeri). Draženje tega centra povzroči obračanje glave in oči v nasprotno smer. Funkcija tega centra je zelo pomembna pri izvajanju tako imenovanih orientacijskih refleksov (ali refleksov "kaj je?"), ki so zelo pomembni za ohranitev živalskega sveta.
Prejema tudi čelna skorja možganskih hemisfer Aktivno sodelovanje pri oblikovanju mišljenja, organizaciji namenskih dejavnosti, dolgoročnem načrtovanju.
Struktura parietalnega režnja
Parietalni reženj zavzema zgornje stranske površine poloble. Od čelnega parietalnega režnja, spredaj in ob strani, ga omejuje osrednji sulkus, od temporalnega od spodaj - stranski sulkus, od okcipitalnega - namišljena črta, ki poteka od zgornjega roba parietalno-okcipitalnega sulkusa do spodnji rob poloble.
Na zgornji stranski površini parietalnega režnja so trije zavoji: ena navpična - posteriorna osrednja in dve vodoravni - zgornja parietalna in inferiorna parietalna. Del spodnjega parietalnega girusa, ki ovija posteriorni del lateralne brazde, imenujemo supramarginalno (supramarginalno), del, ki obdaja zgornji temporalni girus, se imenuje nodalno (angularno) območje.
Parietalni reženj, tako kot čelni reženj, predstavlja pomemben del možganskih hemisfer. V filogenetskem smislu se v njem razlikuje stari del - zadnji osrednji girus, nov - zgornji parietalni girus in novejši - spodnji parietalni girus.
Funkcija parietalnega režnja je povezana z zaznavanjem in analizo občutljivih dražljajev, orientacijo v prostoru. Več funkcionalnih centrov je koncentriranih v vijugah parietalnega režnja.
V posteriornem osrednjem girusu so centri občutljivosti projicirani s projekcijo telesa, podobno kot v sprednjem osrednjem girusu. Obraz je projiciran v spodnji tretjini vijuge, roka in trup v srednji tretjini, noga pa v zgornji tretjini. V zgornjem parietalnem girusu so centri, odgovorni za kompleksne vrste globoka občutljivost: mišično-sklepni, dvodimenzionalno-prostorski čut, občutek za težo in volumen gibanja, občutek za prepoznavanje predmetov na dotik.
Za zgornjimi deli posteriornega osrednjega gyrusa je lokaliziran center, ki zagotavlja sposobnost prepoznavanja lastnega telesa, njegovih delov, njihovih razmerij in medsebojnega položaja (polje 7).
Polja 1, 2, 3 postcentralnega območja sestavljajo glavno kortikalno jedro kožnega analizatorja. Skupaj s poljem 1 je polje 3 primarno, polje 2 pa sekundarna projekcijska cona kože. analizator. Postcentralna regija je povezana z eferentnimi vlakni s subkortikalnimi in stebelnimi formacijami, s precentralno in drugimi področji možganske skorje. Tako je kortikalni del občutljivega analizatorja lokaliziran v parietalnem režnju.
Primarna senzorična področja - to so področja senzorične skorje, katerih draženje ali uničenje povzroči jasne in trajne spremembe v občutljivosti telesa (jedro analizatorjev, po I.P. Pavlovu). Sestavljeni so predvsem iz monomodalnih nevronov in tvorijo občutke enake kakovosti. V osnovni senzorična področja običajno obstaja jasna prostorska (topografska) predstavitev delov telesa, njihovih receptorskih polj.
Okoli primarnih senzoričnih območij so manj lokalizirani sekundarna senzorična področja katerih nevroni se odzivajo na delovanje več dražljajev, tj. so polimodalni.
Najpomembnejše čutno področje je parietalni korteks postcentralni girus in ustrezen del paracentralnega lobula na medialni površini hemisfer, ki je označen kot somatosenzorično področje I. Tukaj je projekcija kožne občutljivosti nasprotne strani telesa iz taktilnih, bolečinskih, temperaturnih receptorjev, interoceptivne občutljivosti in občutljivosti mišično-skeletnega sistema - iz mišičnih, sklepnih, kitnih receptorjev.
Poleg somatosenzoričnega področja I obstajajo manjše velikosti somatosenzorično regija II, nahaja se na meji presečišča osrednjega sulkusa z zgornjim robom temporalnega režnja, v globini stranskega sulkusa. Stopnja lokalizacije delov telesa je tukaj manj izrazita.
Nahaja se v spodnjem parietalnem režnju praxis centri. Praxis razumemo kot namenske gibe, ki so postali avtomatizirani v procesu ponavljanj in vaj, ki se razvijajo v procesu učenja in nenehne vaje v življenju posameznika. Hoja, prehranjevanje, oblačenje, pisanje mehanski element, različne vrste delovna dejavnost(npr. voznikova vožnja, košnja itd.) so prakse. Praxis je najvišja manifestacija motorične funkcije človeka. Izvaja se kot posledica skupnega delovanja različnih področij možganske skorje.
V spodnjih delih sprednjega in zadnjega osrednjega gyrusa se nahaja center za analizo interoceptivnega impulza notranji organi in plovila. Središče je tesno povezano s subkortikalnimi vegetativnimi formacijami.
Struktura temporalnega režnja
Temporalni reženj zavzema inferolateralno površino hemisfer. Od čelnega in parietalnega režnja je temporalni reženj omejen s stranskim žlebom. Na zgornji stranski površini temporalnega režnja so trije zavoji: zgornji, srednji in spodnji.
Zgornja temporalna vijuga leži med silvijsko in zgornjo temporalno brazdo, srednja vijuga med zgornjo in spodnjo temporalno brazdo, spodnja vijuga pa med spodnjim temporalnim sulkusom in prečno medularno fisuro. Na spodnji površini temporalnega režnja se razlikujejo spodnji temporalni girus, lateralni okcipitotemporalni girus in girus hipokampusa (noge morskega konja).
Funkcija temporalnega režnja je povezana z zaznavanjem slušnih, okusnih, vohalnih občutkov, analizo in sintezo govornih zvokov ter spominskimi mehanizmi. Glavno funkcionalno središče zgornje stranske površine temporalnega režnja se nahaja v zgornjem temporalnem girusu. Tu je slušno ali gnostično središče govora (Wernickejevo središče).
Dobro proučeno primarno projekcijsko območje je slušna skorja(polja 41, 42, 52), ki se nahaja v globini lateralnega sulkusa (skorja transverzalnih temporalnih vijug Heschla). Projekcijska skorja temporalnega režnja vključuje tudi Center za vestibularni analizator● v zgornjem in srednjem temporalnem girusu (polja 20 in 21).
Vohalno projekcijsko območje se nahaja v hipokampalnem girusu, zlasti v njegovem sprednjem delu (tako imenovani kavelj). Poleg vohalnih projekcijskih con so okušalne.
Temporalni režnji igrajo pomembno vlogo pri organizaciji kompleksnih duševnih procesov, zlasti spomina.
Struktura okcipitalnega režnja
Okcipitalni reženj zavzema zadnje dele hemisfer. Na konveksni površini hemisfere okcipitalni reženj nima ostrih meja, ki bi ga ločevale od parietalnega in temporalnega režnja, z izjemo zgornjega dela parietalno-okcipitalnega sulkusa, ki se nahaja na notranji površini hemisfere. ločuje parietalni reženj od okcipitalnega režnja. Brazde in zavoji zgornje stranske površine okcipitalnega režnja so nestabilni in imajo spremenljivo strukturo. Na notranji površini okcipitalnega režnja je žleb, ki ločuje klin (trikotna norma lobule okcipitalnega režnja) od lingvalnega gyrusa in occipitotemporal gyrusa.
Funkcija okcipitalnega režnja je povezana z zaznavanjem in obdelavo vizualnih informacij, organizacijo kompleksnih procesov vizualne percepcije - Hkrati se zgornja polovica mrežnice projicira v območju klina, ki zaznava svetloba iz spodnjih vidnih polj; v predelu lingularnega gyrusa je spodnja polovica mrežnice, ki zaznava svetlobo iz zgornjih vidnih polj.
Nahaja se v okcipitalnem korteksu primarno vidno področje(skorja dela sfenoidnega girusa in lingularnega lobula, polje 17). Tukaj je aktualna predstavitev mrežničnih receptorjev. Vsaka točka mrežnice ustreza svojemu območju vidne skorje, območje makule pa ima razmeroma veliko območje predstavitve. V povezavi z nepopolnim križanjem vidnih poti se iste polovice mrežnice projicirajo v vidno območje vsake hemisfere. Prisotnost v vsaki hemisferi projekcije mrežnice obeh očes je osnova binokularnega vida. Lubje se nahaja blizu polja 17 sekundarno vizualno območje(polja 18 in 19). Nevroni teh območij so polimodalni in se ne odzivajo samo na svetlobo, ampak tudi na taktilne in slušne dražljaje. V tem vizualnem območju pride do sinteze različne vrste občutljivost, nastanejo bolj zapletene vidne podobe in izvede se njihovo prepoznavanje.
Otoček, limbična skorja
otok, ali tako imenovana zaprta lobula, ki se nahaja globoko v stranskem žlebu. Otoček je od sosednjih sosednjih delov ločen s krožnim žlebom. Površina otočka je z vzdolžnim osrednjim žlebom razdeljena na sprednji in zadnji del. V otočku je projiciran analizator okusa.
limbični korteks. Na notranji površini hemisfer nad corpus callosum je cingularni gyrus. Ta girus z ožino za corpus callosum prehaja v girus blizu morskega konjička - parahipokampalni girus. Cingulatni girus skupaj s parahipokampalnim girusom tvori obokani girus.
Notranje in spodnje površine hemisfer so združene v tako imenovano limbično (marginalno) skorjo, skupaj z jedrom amigdale iz skupine subkortikalnih jeder, vohalnim traktom in bulbusom, območji čelnega, temporalnega in parietalnega režnja. možgansko skorjo, pa tudi s hipotuberozno regijo in retikularno tvorbo debla. Limbična skorja se združi v eno samo funkcionalni sistem- limbično-retikularni kompleks. Glavna funkcija teh delov možganov ni toliko zagotavljanje komunikacije z zunanjim svetom, temveč uravnavanje tonusa možganske skorje, nagonov in čustvenega življenja. Uravnavajo kompleksne, večplastne funkcije notranjih organov in vedenjske odzive. Limbično-retikularni kompleks je najpomembnejši integrativni sistem telesa. limbični sistem je pomembna tudi pri oblikovanju motivacije. Motivacija (ali notranja motivacija) vključuje najkompleksnejše instinktivne in čustvene reakcije (prehrambene, obrambne, spolne). Limbični sistem sodeluje tudi pri uravnavanju spanja in budnosti.
Limbična skorja opravlja tudi pomembno funkcijo voha. Voh - zaznavanje bivanja v zraku kemične snovi. Človeški vohalni možgani zagotavljajo občutek vonja, pa tudi organizacijo kompleksnih oblik čustvenih in vedenjskih reakcij. Vohalni možgani so del limbičnega sistema.
Vohalni možgani so sestavljeni iz dveh delov - perifernega in osrednjega. Periferni del predstavljajo vohalni živec, vohalne čebulice, primarni vohalni centri. Osrednji del vključuje girus morskega konjička - hipokampus, zobat in obokan girus.
Vohalni receptorski aparat se nahaja v nosni sluznici. Preko sistema živčnih prevodnikov se informacije iz receptorjev prenašajo v kortikalni del vohalnega analizatorja.
Kortikalni predel vohalnega analizatorja se nahaja v cingularnem girusu, girusu morskega konjička in v kavlju morskega konjička, ki skupaj tvorita zaprto obročasto območje. Periferni del olfaktornega analizatorja je povezan s kortikalnimi regijami obeh hemisfer.
Fiziološki mehanizem zaznavanja vonjav z vohalnim analizatorjem ni povsem jasen. Obstajata dve glavni hipotezi, ki pojasnjujeta naravo tega procesa z različnih stališč. Po eni od hipotez se interakcija med molekulami odorantov in kemoreceptorji dogaja kot ključ in ključavnica, tj. vrsta molekule ustreza posebnemu receptorju. Druga hipoteza temelji na predpostavki, da imajo molekule dišeče snovi določen nihajni val, na katerega so "nastavljeni" vohalni receptorji. Molekule, ki imajo podobne vibracije, bi morale imeti skupno valovanje in posledično oddajati podobne vonjave.
Izraz "vohalni možgani" v zvezi s človeško fiziologijo je nekoliko poljuben in ne razkriva v celoti njegove večplastne in univerzalne funkcije. "Umestitev" osrednje vezi vohalnih možganov v možganske hemisfere ni naključna in je posledica velike "informacijske" vloge, ki jo je imel občutek za vonj v procesu evolucije pri prilagajanju zunanjemu okolju in regulaciji kompleksa. vedenjske reakcije. Pridobivanje hrane, izbira osebka nasprotnega spola, skrb za potomce, celovitost ozemlja, organiziranje skupinskih skupnosti znotraj vrste - vse te vsakodnevne funkcije pri mnogih živalih potekajo z neposrednim sodelovanjem fino zgrajenega sistema vohalne recepcije in , ki temelji na tem, sposobnost številnih živali, da pošiljajo subtilne diferencirane specifične vonjave - signale-informatorje.
V življenju ljudi je voh izgubil biološko informacijsko vrednost, ki jo je imel pri živalih. Človeški vohalni sistem je zasnovan tako za opravljanje ozke, "lastne" funkcije kot za nekakšno "polnjenje" čustev. Moč vpliva vonjav na čustveno sfero, da so najpomembnejši »prehranski substrat čustev«, je dobro znana že od pradavnine v zgodovini človeštva.
Človekov čut za vonj je lahko različen. Praviloma so te razlike nepomembne, v nekaterih primerih pa je lahko ostrina vonja zelo visoka (degustatorji v parfumski industriji).
Ker ima vohalni analizator pomembno vlogo pri uravnavanju čustev, je njegov centralni oddelek se nanaša na limbični sistem, figurativno imenovan "skupni imenovalec" za številne čustvene in viscerosomatske reakcije telesa.
Center za analizo okusa se nahaja v neposredni bližini središča olfaktornega analizatorja, tj. v kljuki in Amonovem rogu, poleg tega pa v najnižjem delu posteriornega osrednjega girusa (polje 43), pa tudi v insuli. Tako kot vohalni analizator center zagotavlja funkcijo projekcije, shranjevanje in prepoznavanje vzorcev okusa.
Na meji temporalnega, okcipitalnega in parietalnega režnja se nahaja center za analizo pisnega govora(polje 39), ki je tesno povezano z Wernickejevim središčem temporalnega režnja, s središčem vizualnega analizatorja okcipitalnega režnja, pa tudi s središči temenskega režnja. Bralno središče omogoča prepoznavanje in shranjevanje slik pisnega govora.
Glavni del informacij približno okolje in notranje okolje telesa, ki je vstopilo v senzorično skorjo, se prenese v nadaljnjo obdelavo v asociativno skorjo, po kateri se (če je potrebno) sproži vedenjska reakcija z obveznim sodelovanjem motorične skorje.
corpus callosum- ločna tanka plošča, filogenetsko mlada, povezuje srednje površine obeh hemisfer. Podolgovati srednji del corpus callosum zadaj prehaja v odebelitev, spredaj pa se ločno ukrivlja in ukrivlja navzdol. Corpus callosum povezuje filogenetsko najmlajše dele hemisfer in ima pomembno vlogo pri izmenjavi informacij med njimi.
Arhitektonika možganske skorje
Nauk o strukturnih značilnostih strukture korteksa se imenuje arhitektonika. Celice možganske skorje so manj specializirane kot nevroni drugih delov možganov; kljub temu pa so nekatere njihove skupine anatomsko in fiziološko tesno povezane z določenimi specializiranimi deli možganov.
Mikroskopska zgradba možganske skorje v različnih delih ni enaka. Te morfološke razlike v skorji so omogočile razlikovanje posameznih kortikalnih citoarhitektonskih polj. Obstaja več možnosti za razvrščanje kortikalnih polj. Večina raziskovalcev identificira 50 citoarhitektonskih polj, katerih mikroskopska struktura je precej zapletena.
Korteks je sestavljen iz 6 plasti celic in njihovih vlaken. Glavni tip zgradbe šestplastnega lubja pa ni povsod enoten. Obstajajo področja skorje, kjer je ena od plasti močno izražena, druga pa šibka. Na drugih območjih skorje so nekatere plasti razdeljene na podplasti itd.
Ugotovljeno je bilo, da imajo področja skorje, povezana z določeno funkcijo, podobno strukturo. Področja skorje, ki so po funkcionalnem pomenu blizu živali in ljudi, imajo določeno podobnost v strukturi. Tista področja možganov, ki opravljajo čisto človeške funkcije (govor), so prisotna samo v človeški skorji, medtem ko živali, tudi opice, niso prisotne.
Morfološka in funkcionalna heterogenost možganske skorje je omogočila izločanje centrov za vid, sluh, vonj itd., Ki imajo svojo specifično lokalizacijo. Vendar pa je nepravilno govoriti o kortikalnem centru kot o strogo omejeni skupini nevronov. Specializacija področij korteksa se oblikuje v procesu življenja. V zgodnjih otroštvo funkcionalne cone korteksa se med seboj prekrivajo, zato so njihove meje nejasne in nejasne. Samo v procesu učenja, kopičenja lastne izkušnje praktična dejavnost je postopna koncentracija funkcionalnih območij v centrih, ločenih drug od drugega.
Belo snov možganskih hemisfer sestavljajo živčni prevodniki. V skladu z anatomskimi in funkcionalnimi značilnostmi so vlakna bele snovi razdeljena na asociativna, komisuralna in projekcijska.
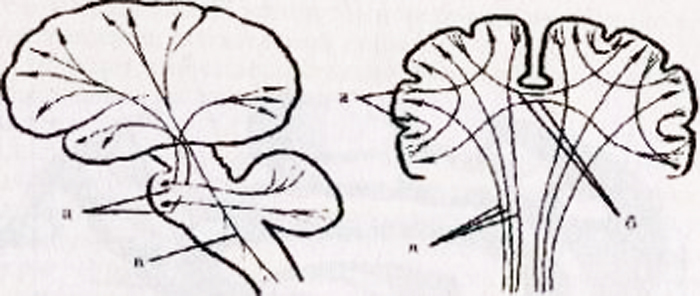
a – asociacijska vlakna; b - komisuralna vlakna; vlakna v projekciji.
Asociativno vlakna se združijo različne razdelke korteks znotraj ene hemisfere. Ta vlakna so kratka in dolga. Kratka vlakna so običajno lokasta in povezujejo sosednje vijuge. Dolga vlakna povezujejo oddaljene dele korteksa. Običajno imenujemo komisuralna vlakna tista vlakna, ki povezujejo topografsko enake dele desne in leve poloble. Komisuralna vlakna tvorijo tri komisure: sprednjo belo komisuro, komisuro forniksa in corpus callosum. Sprednja bela komisura povezuje vohalne regije desne in leve poloble. Forniksna komisura povezuje vijuge hipokampusa desne in leve poloble. Glavna masa komisuralnih vlaken prehaja skozi corpus callosum, ki povezuje simetrične dele obeh hemisfer možganov.
projekcija imenovana tista vlakna, ki povezujejo možganske hemisfere s spodaj ležečimi deli možganov - deblom in hrbtenjačo. V sklopu projekcijskih vlaken so prevodne poti, ki prenašajo aferentne (občutljive) in eferentne (motorične) informacije.
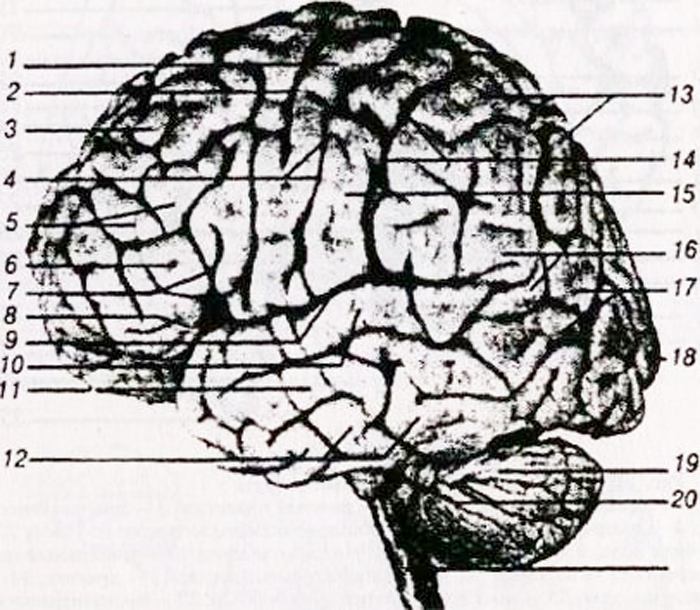
1 - precentralni girus; 2 - precentralna brazda; 3 - zgornji čelni girus; 4 - osrednja brazda; 5 - srednji čelni girus;
b - spodnji čelni girus; 7 - naraščajoča veja stranske brazde;
8 - vodoravna veja stranske brazde; 9 - posteriorna veja stranska brazda; 10 - zgornji temporalni girus; 11 - srednji temporalni girus;
12 - spodnji temporalni girus; 13 - parietalni lobule; 14 - postcentralna brazda; 15 - postcentralni girus; 16 - supramarginalni girus;
17 - kotni girus; 18 - okcipitalni reženj; 19 - mali možgani; 20 - vodoravna razpoka malih možganov; 21 - medulla oblongata

