Mišično-skeletni sistem služi za držanje telesa v določenem položaju in premikanje v prostoru. Mišično-skeletni sistem sestavljajo skeletni sistem (skelet), vezi, sklepi in skeletna mišica. Kosti, vezi in sklepi so pasivni elementi gibalnih organov. Mišice so aktivni del motoričnega sistema.
Evolucija somatomotornega živčnega sistema
V nadaljevanju se bomo ukvarjali le s somatomotoričnim živčevjem, ki oživčuje skeletne mišice. Animacija: molekularni mehanizem mišične kontrakcije. Somatomotorični živčni sistem se spreminja, postopoma se razvija med evolucijo. Razvoj poteka vzporedno s tvorbo skeletnih mišic in mehanizma gibanja, saj so strukture odvisne ena od druge. Med evolucijo lahko o živalih govorimo le, če delujejo mišica pojavi v človeškem telesu. Primitivne, preprosto zgrajene živali z mrežnim živčnim sistemom reagirajo na udarec okolju z močnim gibanjem njihove celote mišična masa, katerega namen je bil izogniti se škodljivi vplivi ali se približati njihovi hrani.
Okostje služi kot opora in zaščita celemu telesu in posameznih teles, številne kosti pa so tudi močni vzvodi, s pomočjo katerih se izvajajo različna gibanja telesa in njegovih delov v prostoru. Mišice poganjajo celoten sistem kostnih vzvodov. Okostje predstavlja osnovo telesa in določa njegovo velikost in obliko. deli okostja, kot je lobanja, rebra in medenico hrbtenica, služijo kot prostor za shranjevanje in zaščito vitalnih organov - možganov, pljuč, srca, črevesja itd. Okostje je vključeno v presnovo, zlasti pri vzdrževanju na določeni ravni mineralna sestava krvi. Poleg tega številne snovi, ki sestavljajo kosti (kalcij, fosfor, citronska kislina itd.), Po potrebi zlahka vstopijo v presnovne procese. Organsko osnovo kostne snovi sestavljajo predvsem beljakovine, mineralno pa kalcijeve in fosforjeve soli.
Njihov preprost živčni sistem in mišični sistem ni bilo dovoljeno več diferenciranega gibanja. Med evolucijo so se razvijala vse bolj popolna gibanja. Postale so možne zaradi razvoja, diferenciacije živčni sistem in mehanizem gibanja ter njihovo usklajevanje.
Mehkužci, ki živijo brez fiksnega okostja, gredo skozi valovite peristaltične kontrakcije svojih mišičnih obročev. Segmente mišičnih obročev nadzirajo ganglijske celice, ki so vzdolžno povezane z živčna vlakna za sinhronizacijo gibanja.
Pri povečavi imajo trde in gladke površine kosti porozno strukturo. obstajati različni tipi kostno tkivo, ki sta največkrat različne dele ena kost: kompaktna plast in gobasta snov. Kosti, kot so vretenca, vrat stegnenice, epifiza polmer, sestojijo predvsem iz gobaste snovi. V gobasti snovi so kostni tramovi razporejeni v obliki ukrivljenih plošč, povezanih s prečnimi ali poševnimi prečkami. Dolge cevaste kosti okončin so sestavljene predvsem iz snovi, v kateri so kostne plošče nameščene zelo tesno.
Sprva difuzno lokaliziran mišična vlakna pozneje tvorijo medsebojno delujoče mišične snope; mišice. Evolucija fiksnega okostja je omogočila hiter razvoj, ki je bil različno rešen pri členonožcih kot zunanje okostje, pri vretenčarjih - kot notranje okostje. Medtem ko sta obe učinkoviti pri gibanju telesa, se je slednja rešitev izkazala za koristnejšo, saj je omogočila nemoteno rast, kar je omogočilo ustvarjanje večjih, kompleksnejših živih bitij.
Živčni sistem členonožcev je segmentiran. Aktivnost okončin v medialnem segmentu nadzirajo segmentni gangliji. Gangliji členonožcev lahko delujejo neodvisno. Čeprav je ganglij v glavi večji od drugega, ne nadzoruje aktivnosti drugih ganglijev. Njegova velikost zadostuje za oskrbo z očmi, antenami in orodji za usta, ki so shranjeni na glavi. Ko členonožcu odrežejo glavo, se lahko žival tudi več dni premika naokoli, dokler ne strada brez ust. Medganglijske povezave zagotavljajo koordinacijo gibanja okončin.
Kosti kot druge notranji organi so sestavljeni iz celic. Obstajajo posebne celice, ki nenehno uničujejo kostno snov (osteoklasti); celice, ki obnavljajo, obnavljajo kost (osteoblasti) in celice, odgovorne za tvorbo kostnega skeleta in mineralizacijo kostnega tkiva (osteociti).
Med življenjem človeka se v kosteh nenehno odvijajo procesi prestrukturiranja kostnega tkiva: na nekem ločenem majhnem območju kostnega tkiva se kostno tkivo uniči, nato pa se odstranjena stara kost nadomesti s popolnoma enako količino nove kosti. . Pri zdravih ljudeh so procesi uničenja kostnega tkiva in tvorbe nove kosti kvantitativno enaki. Delovanje teh celic nadzorujejo številne biološko aktivne snovi, kot so hormoni ščitnice in obščitničnih žlez, hormoni nadledvične skorje, vitamin D3 in končno spolni hormoni (estrogeni in progesteron). Rast in razvoj kostnega tkiva se pojavi do 16-25 let. Po doseganju najvišje ravni kostne mase, do starosti 30-40 let, se začne njena nepomembna izguba, ki je 0,2-0,5% na leto.
V primerjavi z njegovim kompleksna funkcija, je v gangliju presenetljivo malo živčnih celic. Celotna mišica okončine je običajno opremljena s štirimi nevroni. Povezave med nevroni zagotavljajo medsebojno inervacijo med fleksorji in ekstenzorji ter za koordinacijo gibanja z drugimi udi.
Prvi vretenčarji so živali, ki živijo v vodi; ribe. Sprva so bile njihove skeletne mišice sestavljene le iz aksialnih mišic, ki jih je absorbirala vretenčna kislina in so izhajale iz miotoma. Mišice zagotavljajo samo upogibanje in obračanje telesa. Prva homogena aksialna mišična masa se kasneje segmentira; tako so se razvile posamezne mišice. Nato so bile mišice raztegnjene tudi v steni telesa. Kot naslednji rezultat evolucije so plavuti in nato, ko so se nekateri drznili na kopno, razporejeni udi.
V starosti 30-40 let je izguba kostne mase 0,5 % na leto. In po nastopu menopavze se pri ženskah izgubi 3-5% kostne mase na leto.
V človeškem telesu je izoliran tudi mehak skelet (skelet), ki sodeluje pri držanju organov ob kosteh. Mehko okostje vključuje fascije, ligamente, vezivnotkivne kapsule organov in druge strukture. Večina mišic je pritrjenih na kosti. Mišice vključujejo kosti okostja v gibanju in opravljajo delo. Številne mišice, ki obdajajo telesne votline, ščitijo notranje organe.
Mišice, ki so zagotavljale te strukture, izstopijo iz telesne stene. Razvoj udov je bil skokovit korak v evoluciji gibalnega sistema vretenčarjev. Okončine omogočajo živim bitjem učinkovitejše gibanje, zato so se njihove življenjske razmere, prilagoditev na okolje bistveno izboljšale. Med evolucijo sta se njihova struktura in velikost nenehno izboljševali. Okončine, ki so bistveno večje od velikosti telesa, so omogočale dvonožno življenje. Posledično bi lahko bolje krmarili z višjimi čutili v glavi, roke, osvobojene bremena gibanja, pa bi lahko uporabili za druge namene.
Stanje kosti je odvisno od bremena, ki ga prenašajo. Dobro razvito mišično tkivo krepi sklepe in prispeva k normalnemu razvoju in delovanju kosti. Tako mišice kot kosti izgubijo svojo maso, če so obremenitve premajhne. Da bi ohranili mišično-skeletni sistem dolgo časa v zdravem stanju, je potrebno nenehno trenirati, izvajati različne psihične vaje. Vsak človek po 30. letu starosti je obsojen na telesno vzgojo.
Te in temu primerna preureditev malih sklepov rok je spodbudila uporabo orodja, kar je pospešilo tudi razvoj možganov, ki so bili najpomembnejši organ antropoidne prilagodljivosti. Bilateralnost je posebej poudarila razvoj gibalnega sistema in hkrati prispevala, saj vzravnan položaj telesa zahteva veliko več gibalne koordinacije kot stoje na štirih nogah.
Razširitev udov je spremenila tudi mišice. V veliko večjem razpoložljivem prostoru je dovoljeno tako množenje kot podaljšanje okončin. Pomagajo povečati mišično moč in učinkovitost manjših gibov. Razvoj mehanizma gibanja je vretenčarjem dal nove priložnosti. Njihovo gibanje postane popolnejše, hitrejše, natančnejše in ustreznejše. Nujno razvite živčne strukture, ki nadzorujejo delovanje skeletnih mišic. Gibanje gibanja je bilo olajšano s sočasnim razvojem čutil.
To je še posebej pomembno za ženske, katerih kosti so manj goste kot pri moških. Poleg tega so ženske v starosti bolj nagnjene k izgubi mišične mase.
Kostno maso, stopnjo njene tvorbe in njeno izgubo je mogoče nadzorovati dobra prehrana. Kalcij in vitamin D krepita kosti in preprečujeta njihovo pokanje, zlom in druge poškodbe. Potreben dnevni odmerek kalcija je približno 1200-1500 mg za odrasle. Ženske, tako kot moški, do 20. leta pridobijo 50 % svoje kostne mase. V kombinaciji z redno vadbo kalcij spodbuja zdravo in močno kostno maso. Med 20. in 30. letom se kostna masa rahlo poveča, po 30. letu pa se začne proces njene izgube.
Z njihovo pomočjo lahko človek iz okolja pridobiva vse bolj popolne in točne informacije. Da bi izkoristili te priložnosti, so se nevroni, ki podpirajo motorično koordinacijo, razmnožili v možganih in da bi ostali aktivni, so oblikovali vedno bolj kompleksne in intenzivne povezave s čutili. Aksialne mišice so sprva nadzirali izključno hrbtenični motorični nevroni. Po ločitvi posameznih mišic so nadzor nad posameznimi mišicami prevzeli možgani, ki se razvijejo pri vretenčarjih in imajo osrednjo nadzorno vlogo.
V povezavi z biološko funkcijo vašega telesa posebna pozornost zdrava prehrana je treba dati ženskam. Nosečnost in dojenje potrebujejo dodaten kalcij. Če ženska kri ne vsebuje zahtevani znesek tega minerala kosti postanejo dodaten vir kalcija, kar na koncu vodi v njihovo oslabitev. kajenje in prekomerna uporaba alkoholne pijače prispevajo tudi k izgubi kosti. Hormon estrogen uravnava izpiranje kalcija iz kosti in tako ustvarja normalne pogoje za dolgotrajno rast kosti. V menopavzi in po menopavzi pa žensko telo preneha proizvajati estrogen, ki je glavni vzrok za pospešeno izgubo kostne mase. Redna vadba in povečan dnevni vnos kalcija bosta pomagala preprečiti ta proces v postmenstrualnem obdobju. Poleg uživanja kalcija kot prehranskega dopolnila je priporočljivo v vsakodnevno prehrano vključiti živila, ki vsebujejo ta element.
Prvi avtocenter na vretenčarju je bil lokomotorni predel, ki se nahaja v mezencefalonu. Čeprav se je evolucija razvila v nova področja možganov, specializirana za nadzor motorike, retikulospastični trakt obstaja tudi pri ljudeh. Poleg nadzora mišičnega tonusa se ukvarja predvsem z izvajanjem ritmičnih gibov.
Med evolucijo so možgani vretenčarjev sprva prevladovali nad delom sodobnega možganskega debla. Ko so možganske funkcije postale bolj zapletene, je bilo potrebnih več živčnih celic za njihov nadzor. Ker takrat v možganih ni bilo dovolj prostora, so novi nevroni oblikovali novo kolonijo pred do tedaj lociranimi možgani. V naslednjem koraku se oblikujejo tri jedra, odgovorna za vid: dve za in eno nad srednjimi možgani. Prejšnji vretenčarji so imeli tri oči. Podatke dveh stranskih očes sta obdelali dve bazalokativni lateralni genikulatni telesci, ki ju prečkata zgornja parietalna očesa skozi zgornje jedro.
Na program telesne dejavnosti mora biti vključen trening z utežmi. Pomemben dejavnik, ki določa zdravje mišično-skeletnega sistema, je vzdrževanje optimalne teže.
Posebno pozornost je treba nameniti nogam. Noge imajo zelo pomembno vlogo v mišično-skeletnem sistemu, pa tudi pri zdravju celotnega organizma. Pravilna nega za nogami vključuje poleg drugih higienskih postopkov tudi skrbno izbiro čevljev. Udobni in pravilno izbrani čevlji so odlična preventiva pred številnimi boleznimi stopal.
Stranske oči in njihova jedra so se tudi pri človeku ohranila kot vidni organi, hkrati pa se je parietalno oko in njegovo jedro postopoma umaknilo, izgubilo vid in potonilo v globino kot Epithalamus. Naslednja funkcija dienecephalona je bila somatosenzorična. Pri plazilcih se je zdravljenje somatosenzorične funkcije še naprej razvijalo; razvita dorzalna in anteriorna talamusna jedra. Hkrati je bilo anteroventralno jedro prvi koordinacijski center za mezencefalon.
Zadnje področje talamusa, ki se je razvilo, je bil Pulvinar Talami, ki je skrbel za koordinacijo med čutili. Pri nesesalcih je prevzel celotno vlogo treh čutnih režnjev možganov. Bazalne ganglije, ki usklajujejo gibalni sistem, so našli že pri dvoživkah. Hitro so se razvili, a le pri sesalcih.
"Mišično-skeletni sistem" in drugi članki iz razdelka Bolezni mišično-skeletnega sistema
Funkcionalna struktura prostovoljnega gibanja. Iz zgoraj navedenega izhaja, da so različne komponente vključene v zagotavljanje kakršnega koli gibanja, zato je eno glavnih vprašanj, kako zagotoviti enkraten ukaz, ki ga prejme izvršilni aparat. Ne glede na strategijo in taktiko posameznega gibanja je glavna naloga sistema, ki zagotavlja program, koordinacija vseh komponent ekipe.
Pri dvoživkah že nastajajo usedline uranovega telencefalona, ki so se razširile iz stranskih sten dienske celice. Prve telencefalne strukture ustrezajo rastlinam bazalnih ganglijev in limbičnemu sistemu sesalcev. Njihovi glavni nalogi sta bili voh in usklajevanje nekaterih, predvsem spolnih vedenj. Možganska skorja je "izum" sesalcev. Glede na zgornje razloge najdemo takšne nove funkcije, katerih nevroni niso našli dovolj prostora v diencecephalonu.
Ker dienecephalon ni mogel več rasti, so iz stene nevroepitelijske cevi, ki je zrasla iz njegove stene, nastali novi nevroni. Postopoma so se razvili stranski ventrikli, iz njihovih sten pa režnji možganov. Prvi sesalci so bili majhni paličasti glodalci, ki so sprva švigali naokoli, nato pa so postali vrečasti. V dobi dinozavrov so se zatekli v jame v podzemni železnici. V skladu s svojim načinom življenja so v temnih jamah krmarili predvsem po vonju.
CNS ima številne genetsko določene programe (na primer program lokomotorne hoje, ki temelji na dejavnosti hrbtenični - ki se nanaša na hrbtenico.");" onmouseout="nd();" href="javascript:void(0);">hrbtenice generator). Tako preproste programe združujemo v kompleksnejše sisteme, kot je vzdrževanje pokončne drže. Ta povezava se pojavi kot posledica učenja, ki je zagotovljeno s sodelovanjem sprednje skorje. hemisfere.
Najbolj kompleksna in filogenetsko najmlajša je sposobnost oblikovanja zaporedja gibov in predvidevanja njegovega izvajanja. Rešitev tega problema je povezana s frontalnim asociativnim sistemom, ki si zapomni in shrani v spomin takšna zaporedja gibov. Najvišji odraz tega kodiranja v človeku je verbalizacija oziroma verbalna spremljava osnovnih konceptov gibanja.
Splošna pravilnost sistema za nadzor gibanja je uporaba povratnih informacij. To ne vključuje samo proprioceptivne povratne informacije od gibanja, ki se je začelo, ampak tudi Aktivacija - vzbujanje ali povečana aktivnost, prehod iz stanja mirovanja v aktivno stanje. onmouseout="nd();" href="javascript:void(0);">aktivacijo sistemi nagrajevanja ali kaznovanja. Poleg tega so vključene tudi notranje povratne informacije, tj. informacije o aktivnosti spodnjih ravni motoričnega sistema ali eferentna kopija samega motoričnega ukaza. Ta vrsta povratne informacije je potrebna za razvoj novih motoričnih koordinacij. Za gibe različne kompleksnosti in hitrosti se lahko povratna informacija zapre na različnih ravneh. Zato lahko obe vrsti nadzora - programiranje in sledenje - sobivata v istem sistemu za nadzor gibanja.
Na koncu je priporočljivo navesti izjavo izjemnega fiziologa N.A. Bernstein, da gibi "...niso prostorski, ampak pomenski in motorične komponente verig akcijske ravni narekujejo in izbirajo glede na pomensko bistvo predmeta in kaj je treba z njim narediti."
Za preživetje sta dve najpomembnejši funkciji vonj in razmnoževanje. Tako sta bili ti dve funkciji obravnavani v dveh novih regijah v razvoju iz dienecefalona. njihovi ostanki, limbični sistem in dišeči korteks, ki opravljata enake funkcije, najdemo tudi pri ljudeh v medialni osrednji skorji. Ti dve funkciji sta morfološko in funkcionalno tesno povezani pri vsakem vretenčarju, tudi pri ljudeh z močno zmanjšanim vonjem. Na tem stališču temelji industrija parfumov.
Hemisfere corpus callosum v možganih najstarejših preživelih sesalcev, kloakov in ptic niso prisotne. Naslednji korak v evoluciji je bil pomemben razvoj somatosenzoričnega sistema. Ti nočni glodavci, podobni današnjim podganam in mišem, so še vedno živeli v temi pod zemljo. S svojimi so lahko hitro in zanesljivo krmarili po temnih podzemnih kanalih dolgi lasje na obrazu, saj bi jih lahko uporabljali kot radar, preko zelo občutljivih tlačnih receptorjev povezanih. Za to natančno mehansko "analizo slike" so potrebovali novo populacijo nevronov.
10.4. Elektrofiziološki korelati organizacije gibanja
Elektrofiziološke metode se uporabljajo za preučevanje različnih vidikov motorična aktivnost, predvsem pa tiste, ki so nedostopne neposrednemu opazovanju. Dragocene informacije o fizioloških mehanizmih organizacije gibanja dajejo metode za ocenjevanje medsebojnega delovanja con možganske skorje, analiza lokalnega EEG in z gibom povezanih potencialov ter beleženje nevronske aktivnosti.
Preučevanje medzonskih povezav možganskih biopotencialov omogoča sledenje dinamiki interakcije posameznih kortikalnih con na različnih stopnjah izvajanja gibanja, pri učenju novih motoričnih veščin, ugotavljanje posebnosti medzonskega medsebojnega delovanja med različni tipi gibanja.
Ti nevroni so migrirali v novonastali teleencefalon in oblikovali prvi pravi reženj - parietalni reženj. Ta loputa je ostala tudi somatosenzorični center pri človeku. Pri oposumu in podobnih vrstah se možganska skorja uporablja predvsem za celjenje kože. Pri starih sesalcih so signali tekli od mišičnih vreten do na novo razvitih malih možganov in od tam skozi jedra malih možganov do talamusa.
Kot naslednja stopnja evolucije so se možgani zajcem podobnih glodalcev, ki so se že prebijali na površje zemlje, soočili z novim izzivom: poslušati so morali od daleč in poiskati morebitnega napadalnega sovražnika, da bi lahko pobegnili. čas. Novo akustično središče se je preselilo iz spodnjega kolikulusa v temporalni reženj, ki se razvije iz stene nevroepitelijske cevi, ki izrašča iz dienozifalona in se zaradi pomanjkanja prostora zaniha navzdol. Deli stranski ventrikel, ki vsebuje dve žilni steni pleksusa, celični medij in spodnjo koruzo, pa tudi parietalno in temporalni reženj, ki se razvijejo iz njegove stene, so najstarejši deli možganske skorje.
Prostorska sinhronizacija (PS), tj. Sinhronizirana dinamika električnih nihanj, posnetih iz različnih točk možganske skorje, odraža stanje možganskih struktur, kar olajša širjenje vzbujanja in ustvarja pogoje za medzonsko interakcijo. Metodo registracije PS je razvil izjemen ruski fiziolog M.N. Livanov.
Študije ritmičnih komponent EEG posameznih con in njihovih prostorsko-časovnih odnosov pri ljudeh med izvajanjem prostovoljnih gibov so dale prava priložnost pristopiti k analizi centralnih mehanizmov funkcionalnih interakcij, ki se razvijejo na sistemski ravni med gibalno aktivnostjo. Korelacijska analiza EEG, posnetega med izvajanjem ritmičnih gibov, je pokazala, da pri človeku pri kortikalni organizaciji gibov sodelujejo ne le centri motorične skorje, temveč tudi frontalna in spodnja parietalna cona.
Učenje prostovoljnih gibov in njihovo urjenje povzročita prerazporeditev intercentralnih korelacije kortikalnih biopotencialov. Na začetku treninga skupno število centri, vključeni v skupno aktivnost, se močno povečajo, sinhrono razmerje ritmičnih komponent EEG motoričnih con s sprednjo in zadnjo Asociativne cone korteksa - cone, ki prejemajo informacije od receptorjev, ki zaznavajo draženje različnih modalitet, in iz vseh projekcijskih con. ");" onmouseout="nd();" href="javascript:void(0);">asociativno področja. Z obvladovanjem gibanja se splošna raven PS znatno zmanjša, nasprotno pa se povečajo povezave med motoričnimi in spodnjimi parietalnimi conami. 
Pomembno je omeniti, da se v procesu učenja ritmična sestava biopotencialov različnih kortikalnih con preuredi: v EEG se začnejo beležiti počasni ritmi, ki po frekvenci sovpadajo z ritmom izvajanja gibov. Ti ritmi v človeškem EEG se imenujejo "označeni". Enaka označena nihanja so bila ugotovljena pri otrocih predšolska starost ko izvajajo ritmične gibe na ergografu.
Sistematične študije človeškega EEG med izvajanjem ciklične (občasno ponavljajoče se) in aciklične motorične aktivnosti so pokazale pomembne spremembe v dinamiki električne aktivnosti možganske skorje. V EEG se poveča tako lokalna kot oddaljena sinhronizacija biopotencialov, kar se izraža v povečanju moči periodičnih komponent, v spremembah frekvenčnega spektra avto- in navzkrižnih korelogramov, v določeni poravnavi maksimumi frekvenčnih spektrov in funkcij Koherenca - stopnja sinhronizacije indikatorjev frekvence EEG med različnimi deli možganske skorje.");" onmouseout="nd();" href="javascript:void(0);">skladnost na isti frekvenci.
PS in reakcijski čas. Reakcijski čas je eden najpreprostejših motoričnih indikatorjev. Zato je še posebej zanimivo dejstvo, da ima lahko tudi preprosta motorična reakcija različne fiziološke korelate glede na povečanje ali zmanjšanje njenega trajanja. Tako se je pri primerjavi slike medcentralnih korelacijskih razmerij spektralnih komponent EEG možganov s časom preproste motorične reakcije izkazalo, da je preureditev prostorsko-časovnih razmerij EEG asociativnih con povezana z odzivom časa do danega dražljaja. Pri hitrih reakcijah pri zdravem človeku so se najpogosteje pojavile visoke korelacije biopotencialov v obeh spodnjih parietalnih predelih (nekoliko bolj z levo hemisfero možganov). Če se je reakcijski čas podaljšal, je to spremljala sinhronizacija biopotencialov v čelne regije korteks in spodnji temenski predel leve poloble je bil izključen iz interakcije. Poleg tega je bilo ugotovljeno razmerje med velikostmi faznih premikov Alfa ritem - glavni ritem elektroencefalograma v stanju relativnega mirovanja s frekvenco v območju 8 - 14 Hz in povprečno amplitudo 30 - 70 μV. ");" onmouseout="nd();" href="javascript:void(0);">ritem alfa registrirano v frontalni, precentralni in okcipitalna področja možganov in hitrost preproste motorične reakcije.
Pomembno je omeniti, da se povečanje sinhronizacije biopotencialov pojavi pri osebi že v preddelovnem stanju v procesu koncentracije pred motorično akcijo, pa tudi med mentalnim izvajanjem gibov.
PS in specifike gibanja. Poleg nespecifičnega povečanja PS biopotencialov je bilo ugotovljeno njegovo izrazito selektivno povečanje med kortikalnimi conami, ki so neposredno vključene v organizacijo določenega motoričnega dejanja. Na primer, največja podobnost v električni aktivnosti je ugotovljena: pri premikanju rok - med čelno regijo in motorično reprezentacijo mišic zgornjih udov; pri premikanju nog - med čelno regijo in motorično reprezentacijo mišic spodnjih okončin. Z natančnimi dejanji, ki zahtevajo fino orientacijo v prostoru in vizualni nadzor (streljanje, sabljanje, košarka), so interakcije med vidnimi in motoričnimi področji okrepljene.
Kompleksna dinamika PS biopotencialov različnih delov možganov pri športnikih je bila razkrita med izvajanjem različne vaje in prikazana je odvisnost povečanja interakcije ritmičnih komponent EEG od načina motorične aktivnosti, od kvalifikacij športnikov, od sposobnosti osebe za reševanje taktičnih problemov, od kompleksnosti situacije. Tako so pri visokokvalificiranih športnikih medcentralne interakcije veliko bolj intenzivne in jasneje lokalizirane. Izkazalo se je tudi, da zahtevnejše motorične naloge zahtevajo več visoka stopnja prostorska sinhronizacija ritmov EEG, čas za reševanje taktičnih nalog pa je v korelaciji s hitrostjo povečanja intercentralnih interakcij. V tem primeru motorični odziv sledi po doseženi maksimalni sinhronosti biopotencialov v možganski skorji.
Skupne študije PS možganskih biopotencialov pri ljudeh so omogočile ugotovitev, da pri izvajanju enostavnih in zapletenih motoričnih dejanj različni možganski centri vstopijo v interakcije in tvorijo kompleksne sisteme med seboj povezanih območij z žarišči aktivnosti ne le v projekciji, ampak temveč tudi v asociativnih območjih, zlasti čelnem in spodnjem temenskem. Te medcentralne interakcije so dinamične in se spreminjajo v času in prostoru, ko se izvaja motorično dejanje.
10.5. Kompleks možganskih potencialov, povezanih z gibi
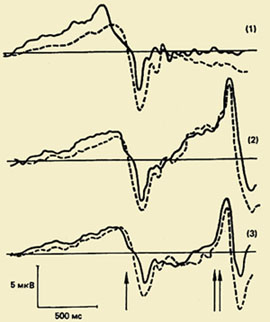 Ena od pomembnih smeri preučevanja psihofiziologije motoričnega akta je preučevanje kompleksa nihanj možganskih potencialov, povezanih z gibi (PMSD). Pomen tega pojava za razumevanje fiziološki mehanizmi organizacija gibanja je zelo velika, saj študija PMSD omogoča razkritje skritega zaporedja procesov, ki se dogajajo v možganski skorji med pripravo in izvedbo gibanja, kronometrija procesov obdelave informacij pa je skupek metod za merjenje trajanje posameznih stopenj v procesu obdelave informacij na podlagi merjenja fiziološki indikatorji, zlasti latentna obdobja komponent evociranih in z dogodki povezanih potencialov.");" onmouseout="nd();" href="javascript:void(0);"> ura ti procesi, tj. določijo svoje časovne omejitve.
Ena od pomembnih smeri preučevanja psihofiziologije motoričnega akta je preučevanje kompleksa nihanj možganskih potencialov, povezanih z gibi (PMSD). Pomen tega pojava za razumevanje fiziološki mehanizmi organizacija gibanja je zelo velika, saj študija PMSD omogoča razkritje skritega zaporedja procesov, ki se dogajajo v možganski skorji med pripravo in izvedbo gibanja, kronometrija procesov obdelave informacij pa je skupek metod za merjenje trajanje posameznih stopenj v procesu obdelave informacij na podlagi merjenja fiziološki indikatorji, zlasti latentna obdobja komponent evociranih in z dogodki povezanih potencialov.");" onmouseout="nd();" href="javascript:void(0);"> ura ti procesi, tj. določijo svoje časovne omejitve.
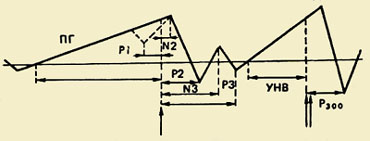
Komponentna sestava PMSD. Prvič je bil ta kompleks, ki odraža procese priprave, izvedbe in vrednotenja gibanja, registriran v 60. letih prejšnjega stoletja. Izkazalo se je, da pred gibanjem sledi počasno negativno nihanje - potencial pripravljenosti (RP). Začne se razvijati 1,5 - 0,5 s pred začetkom gibanja. Ta komponenta je registrirana predvsem v osrednjih in fronto-centralnih odvodih obeh hemisfer. 500-300 ms pred začetkom gibanja postane PG asimetričen - njegova največja amplituda se opazi v precentralnem območju, Kontralateralno - nanaša se na nasprotno stran telesa.");" onmouseout="nd();" href="javascript:void(0);"> kontralateralno premikanje. Pri približno polovici odraslih preiskovancev se na ozadju tega počasnega negativnega nihanja tik pred začetkom gibanja zabeleži majhna amplitudna pozitivna komponenta. Imenuje se "premotorna pozitivnost" (PMP). Naslednje hitro naraščajoče negativno nihanje po amplitudi, tako imenovani motorični potencial (MP), se začne razvijati 150 ms pred začetkom gibanja in doseže največjo amplitudo v območju motorične reprezentacije gibljivega uda v možganski skorji. Ta kompleks potencialov se zaključi s pozitivno komponento približno 200 ms po začetku gibanja.
Funkcionalni pomen komponent. Splošno sprejeto je, da potencial pripravljenosti (RP) nastane v motoričnem korteksu in je povezan s procesi načrtovanja in priprave gibanja. Spada v razred počasnih negativnih nihanj možganskega potenciala, katerih pojav je razložen z aktivacijo nevronskih elementov ustreznih območij korteksa.
Hipoteze glede funkcionalna vrednost PMP so drugačni.
To nihanje se obravnava kot odraz prenosa centralnega ukaza iz skorje v mišice in kot rezultat sprostitve skorje po zaključku določene stopnje organizacije gibanja ter kot odraz procesov zatiranja povezanih gibov drugega uda in kot povratna informacija iz aferentnih mišic. Trenutno nekateri avtorji verjamejo, da so PMP le odraz nastopa motoričnega potenciala.
Pri registraciji MP pri opicah sta bili v MP identificirani dve podkomponenti. Prva podkomponenta je povezana z aktivacijo motoričnega korteksa, povezanega z začetkom gibanja (sinaptična aktivnost piramidni nevroni), drugi pa - z aktivacijo polj 2,3 in 4 Brodmannovega polja - ločena področja možganske skorje, ki se razlikujejo po svoji celični strukturi (citoarhitektoniki) in funkcijah. Na primer, polja 17,18,19 so vidna področja možganske skorje, ki imajo različne strukture in funkcije pri zagotavljanju vizualne percepcije.");" onmouseout="nd();" href="javascript:void(0);">avtor Brodman. Registracija MP pri osebi z epilepsijo je omogočila razlikovanje treh komponent v njem. Prva komponenta se je imenovala iniciacijski potencial. Ima visoko amplitudo in se pojavi po začetku gibanja v precentralnem kontralateralnem korteksu. Drugi, ki se pojavi po začetku miograma in je bolj lokaliziran v kontralateralnem somatosenzoričnem polju, je lahko povezan z iniciacijo gibanja in senzorično povratno informacijo. Tretja komponenta odraža impulz, ki prihaja iz mišičnih aferentov v skorjo.
Pozitivni potencial, ki sledi MP, se obravnava kot odraz povratne aferentacije, ki prihaja iz perifernih receptorjev, naraščajoče aktivnosti iz motoričnih centrov, operacije primerjave med motoričnim programom in nevronsko sliko njegovega izvajanja ali kortikalnih procesov. sprostitev po gibanju.
Val pričakovanja. Poleg PMSD je opisan še en elektrofiziološki fenomen, ki je v bistvu blizu potencialu pripravljenosti. To je približno o negativnem nihanju potenciala, zabeleženem v sprednjih delih možganske skorje v obdobju med delovanjem opozorilnih in sprožilnih (ki zahtevajo reakcijo) signalov. To nihanje ima več imen: val pričakovanja, E-val, pogojno negativno odstopanje (CNV). E-val se pojavi 500 ms po opozorilnem signalu, njegovo trajanje se povečuje s povečevanjem intervala med prvim in drugim dražljajem. Amplituda E-vala narašča premosorazmerno s hitrostjo motoričnega odziva na začetni dražljaj. Povečuje se z napetostjo pozornosti in povečanjem voljnega napora, kar kaže na povezavo tega elektrofiziološkega pojava z mehanizmi prostovoljne regulacije motorične aktivnosti in vedenja na splošno.
10.6. nevronska aktivnost
Funkcionalni kortikalni stebri. V motorični coni človeške skorje so tako imenovane velikanske piramide Beza celice - piramidne celice možganske skorje. onmouseout="nd();" href="javascript:void(0);">Betz celice, ki so organizirani v različnih stolpcih. Piramidne celice, ki opravljajo podobne funkcije, se nahajajo ena poleg druge, sicer bi težko razložili natančno somatotopsko organizacijo skorje. Takšni motorični stebri lahko vzbudijo ali zavirajo skupino funkcionalno homogenih Motorni nevron (motorni nevron) - živčna celica, katere akson inervira mišična vlakna. onmouseout="nd();" href="javascript:void(0);">motorični nevroni.
Registracija aktivnosti posameznih piramidnih celic z implantiranimi mikroelektrodami pri živalih, ki izvajajo različne gibe, je omogočila ugotovitev bistveno pomembnega dejstva. Kortikalni nevroni, ki uravnavajo aktivnost katere koli mišice, niso koncentrirani v samo enem stolpcu. Motorični steber je v veliki meri funkcionalna zveza nevronov, ki uravnavajo aktivnost več mišic, ki delujejo na določen sklep. Tako so v stolpcih piramidnih nevronov motorične skorje gibi predstavljeni ne toliko kot mišice.
Nevronske kode motoričnih programov. Kodiranje informacij v nevronu se izvaja s frekvenco njegovih izpustov. Analiza impulzne aktivnosti nevronov med razvojem različnih motoričnih programov pri živalih je pokazala, da nevroni sodelujejo pri njihovi izgradnji. različne oddelke motorični sistem, medtem ko opravlja določene funkcije. Po nekaterih idejah se vključitev motoričnih programov pojavi zaradi aktivacije tako imenovanih ukaznih nevronov. Ukazni nevroni pa so pod nadzorom višjih kortikalnih centrov. Inhibicija je živčni proces, ki je nasproten vzbujanju; se kaže v oslabitvi ali prenehanju aktivnosti, značilne za določen sistem telesa. onmouseout="nd();" href="javascript:void(0);">Upočasni ukazni nevron vodi do ustavitve programa, ki ga nadzira, vzbujanje, nasprotno, do aktivacije živčnega kroga in aktualizacije motoričnega programa.
Vključenost ukaznih nevronov v celovito aktivnost možganov je določena s trenutno motivacijo in specifičnim motoričnim programom, namenjenim zadovoljevanju te motivacije. Motorni program, da bi bil po naravi prilagodljiv, mora upoštevati vse signalno pomembne komponente zunanjega okolja, glede na katere se izvaja namensko gibanje, tj. ki temelji na principu multisenzorike Konvergenca je združevanje aksonov skupine nevronov, ki nastane zaradi tvorbe sinaps na istem postsinaptičnem nevronu. onmouseout="nd();" href="javascript:void(0);">konvergenco.
Slovar izrazov
- lokomotiva
- motorični nevron
- artikulacija
- aktiviranje
- skladnost
- Brodmannova polja
- kronometrija procesov obdelave informacij
Vprašanja za samopregledovanje
- Kakšna je razlika med statično in dinamično podobo telesa?
- Katere strukture možganov igrajo odločilno vlogo pri organizaciji prostovoljnega gibanja?
- Kako se razlikujeta funkciji piramidnega in ekstrapiramidnega sistema?
- Kaj potenciali možganov, povezani z gibanjem, dajejo za razumevanje možganskih mehanizmov organizacije gibanja?
Bibliografija
- Batuev A.S. Funkcije motoričnega analizatorja. L.: Nauka, 1970.
- Batuev A.S. višje živčna dejavnost. Moskva: Višja šola, 1991.
- Bernstein N.A. Eseji o fiziologiji gibov in fiziologiji dejavnosti. Moskva: Nauka, 1966.
- Bloom F., Lizerson A., Hofstadter L. Možgani, um in vedenje. M.: Mir, 1988.
- Dudel J., Ruegg I., Schmidt R., Janig V. Človeška fiziologija. T. 1 / Ed. R. Schmidt in G. Thevs. M.: Mir, 1985.
- Splošni potek fiziologije človeka in živali / Ed. A.A. Nozdračev. Moskva: Višja šola, 1991.
- Sologub E.B. Električna aktivnost človeških možganov v procesu motorične aktivnosti. L.: Nauka, 1973.
- Krizman T.P. Gibanje otroka in električna aktivnost možganov. Moskva: Pedagogika, 1973.
- Evarts E. Možganski mehanizmi, ki nadzorujejo gibanje // Brain. M.: Mir, 1982.
Teme seminarskih nalog in esejev
- Nauki N.A. Bernstein o strukturi gibanja.
- Psihofiziološke tipologije gibov.
- Gibanje človeških rok in njihovi nevrofiziološki mehanizmi.
- Prostovoljni gibi in njihovi nevrofiziološki mehanizmi.
- Zgodovinske faze raziskovanja možganskih potencialov, povezanih z gibi.
- Vloga nevronske aktivnosti pri konstrukciji gibov.

