5-10-2012, 19:14
Aivokuoren toiminnot
etulohko. Etulohkojen päätehtävä on vapaaehtoisten liikkeiden hallinta, ekspressiivisen puheen motoristen mekanismien koordinointi sekä "luovan" tai abstraktin ajattelun tarjoaminen.
Etulohko voidaan jakaa viiteen alueeseen:
- moottori,
- esimoottori,
- etumotorinen visuaalinen,
- Brocan motorinen puhealue
- ja etuosan assosiaatioalue
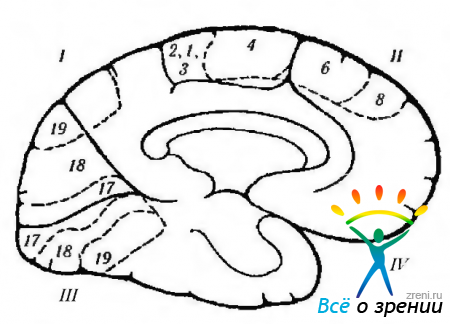
Riisi. 4.1.12. Kaavamainen esitys aivokuoren toiminnoista isot aivot, lokalisoituvat mediaaliselle pinnalle, ja niiden rikkominen aikana erilaisia sairauksia. johon liittyy aivokuoren kiihtyminen tai tuhoutuminen (Crosbyn et al., 1962 mukaan): 1 - parietaalinen lohko. Keskigyrus (4). Vasen takaraivoalue: -Aleksia ilman agrafiaa. Takakuori: -vaikeudet kääntää katsetta; - Balintin oireyhtymä (kaksipuolinen), 11 - otsalohko. Lisämoottorikenttä (6): + vastakkaisen käden nostaminen, silmien ja pään kääntäminen käteen; + pupillien supistuminen; + puheen viive. Etumoottorin silmäkenttä (8). Vyön gyrus: + pupillin laajeneminen; + kyynelvuoto. kiertoradan pinta. III - takaraivolohko. primaarinen näkökuori. Vastapuolinen näkö: - oikeanpuoleinen hemianopsia. IV-oimolohko: silmän liikkeiden halvaantuminen; ipsilateral hemiplegia; vesipää; deserebraalinen jäykkyys; pedunkulaarinen hallusinoosi; kontralateraalinen hemianopsia; parinaudin oireyhtymä. Pinnan alla: - kasvojentunnistus. Huomautus. Sytoarkkitehtoniset kentät vastaavat Brodmannin luokittelua. Plus osoittaa aivokuoren sähköisen stimulaation tuloksen ja miinus - kentän tuhoutumisen tuloksen
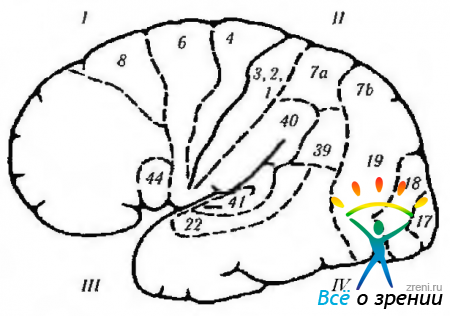
Riisi. 4.1.13. Aivojen sivupinta (nimitykset ovat samanlaiset kuin edellisessä kuvassa): 1 - etulohko. Moottorikenttä (4). Esimoottorikenttä (6). Motorinen silmäkenttä (8): + kontralateral saccade; + vastakkaisen silmän sulkeminen; + silmän gomolateral ystävällinen poikkeama. Motorinen puhekenttä (44): - motorinen (motorinen) afasia. 11 - parietaalilohko: - (- yksinkertainen hallusinoosi; - illuusiot; - metamorfopsia: - huomiokyvyn menetys; - agnosia; - apraksia; - optokineettisen nystagman heikkeneminen; - alempi kvadrantin hemianopia. Supramarginaalinen gyrus (40): - muistinmenetys; - ammattikieltä afasia. Angular gyrus (39): - agraphia - acalculia - menetys kyky erottaa vasen ja oikea oikea puoli; sormen agnosia. somaattinen herkkyys. III-oimolohko: + visuaaliset hallusinaatiot; + deja vu; + heikentynyt hajuaisti; + kohtaukset; + automatismit; - ylempi kvadrianopsia; -Muistin menetys; - Klüver-Bucyn oireyhtymä. primaarinen kuulokuori. Toissijainen kuulokuori: -kuuloafasia. Planum temporale: - lukihäiriö. IV-niskakyhmy (17, 18, 19): + valon tunne; -(-värien tunne; -(-liikkeen tunne; + lineaaristen kuvien tunne. Visuaaliset assosiatiiviset kentät (18, 19): -(-kontralateraaliset silmän liikkeet, kuten tasainen tavoittelu); + vergenssiliikkeet; -Antonin oireyhtymä
Ensisijainen moottorialue(moottorialue; 4. kenttä Brodmanin mukaan).
moottorialue b on keskellä olevan uurteen edessä, joka erottaa otsalohkon parietaalilohkosta. Se koostuu suuresta määrästä Betzin jättimäisiä pyramidisoluja.
Se edustaa liikkeitä, ei yksittäisiä lihaksia. Liikkeet ohjaavat solut sijaitsevat siten, että vartalon alaosan liikkeet näkyvät gyrusen yläosassa ja ylemmän puoliskon liikkeet alaosassa. Neuronaaliset aksonit laskeutuvat aivojen sisäisen kapselin ja varsien läpi kortikobulbaari- ja kortikospinaalisten teiden muodossa.
Motorisen aivokuoren poisto johtaa halvaantumiseen.
Premotorinen alue(kenttä b) sijaitsee suoraan ensisijaisen etupuolella motorinen aivokuori, joka sijaitsee kolmen vaakasuunnassa suunnatun etupyörän takana.
Se muodostaa frontopontocerebellaaristen teiden kuidut ja antaa myös lukuisia kuituja tyvihermoille.
On ehdotettu, että esimotorinen aivokuori osallistuu motoristen toimintojen suunnitteluun. Tästä on osoituksena se tosiasia, että kuudennen kentän hermosolut innostuvat vasta aiottua toimintaa ajatellen. Näiden hermosolujen stimulaatio johtaa käsivarren kohotukseen vastakkainen puoli, ja pää ja silmät kääntyvät kohotetun raajan suuntaan. Pupillit laajentuvat ja puhe heikkenee (katso kuva 4.1.12, 4.1.13).
Tämän alueen tuhoaminen liittyy vastapuolen pseudocerebellaarinen ataksia. Myös kyky kopioida opittuja liikkeitä (apraksia) on heikentynyt. Tällöin yksittäiset liikkeet eivät vaurioidu, mutta potilas ei pysty suorittamaan niitä halutussa järjestyksessä.
Etumoottorin näköalue(kenttä 8 Brodmanin mukaan).
Tämä alue (kenttä 8 ja osittain 6 ja 9) peittää mediaalisen etumyrskyn (gyrus frontalis medius) takaosan.
Etumoottorin näköalue tärkeä tahdissa silmien liikkeissä. Jotkut alueet määrittelevät lähentymisen, eron ja mukautumisen.
On huomattava, että silmien, pupillien ja silmäluomien liike voidaan havaita sähköstimulaation aikana lähes kaikilla aivopuoliskon alueilla. Etuosan motorisen silmäalueen sähköinen stimulaatio johtaa sakkadisiin liikkeisiin, toisin kuin vaste kenttien 18, 19 ja 22 stimulaatioon (tasainen seuranta ja vergenssiliikkeet). Apinoiden 8. kentän stimulaatioon liittyy samanaikainen silmien vastakkainen poikkeama. Tässä tapauksessa oppilas laajenee ja pää ja vartalo kääntyvät vastakkaiseen suuntaan. Etulohkon vaurioituminen aiheuttaa palautuvan kyvyn vastakkaisiin sakkadisiin liikkeisiin.
Tämä aivokuoren alue erottuu lukuisista yhteyksistä.
Afferentit kuidut sisään visuaaliseen etukuoreen näkötuberkkelistä sekä ylemmästä temporaalisesta, peristriataalisesta ja parietaalisesta (alue 7) aivokuoresta. Täältä on löydetty neuroneja, jotka ohjaavat silmien liikkeitä.
Efferentit projisoituna tyvihermosolmuihin, talamukseen, preektaaliseen alueeseen (mukaan lukien näkökanavan tuma), quadrigeminan yläkollikuuksiin ja osaan retikulaarinen muodostuminen silta.
Toisaalta on huomattava, että etuosan näköalueet projisoidaan kontralateraalisille frontaalisille alueille ja kortikaalialueille samalta puolelta. Ne tarjoavat visuaalisen havainnon (7. kentällä parietaalinen aivokuori ja peristriataalinen aivokuori).
Kun heijastetaan päälle keskiaivot prefrontaalisen aivokuoren kuidut ylittävät sisäisen kapselin anteriorisen osan ja jakautuvat dorsaalisiin ja ventraalisiin reitteihin rostraalisessa välilihassa.
Dorsaalinen transtalaminen reitti ylittää talamus opticuksen dorsomediaalisen ja intralamylaarisen ytimen sekä tyynyn keskiosan muodostaen samalla ei suuri määrä synaptisia yhteyksiä. Synapseja löytyy myös preektaalisista ytimistä ja quadrigeminan ylemmästä tuberkkelista.
Ventral pedunculo-tegmentaalinen reitti kulkee aivorungon ventraalisimmassa osassa, toimittamalla kuituja hypotalamuksen ytimiin ja syvemmälle - quadrigeminan ylempään tuberkuloosiin. Samanaikaisesti etuosan visuaalisilla alueilla on topografinen projektio quadrigeminan ylemmille tuberkeille. Polku päättyy sillan verkkomaiseen muodostukseen.
Kolmas eturintaliide esiintyy diencephalic-mesencephalic -liitoksen alueella, joka sijaitsee lähellä punaista ydintä. Tämä polku projisoidaan mediaalisen pitkittäisen fasciculuksen ytimeen ja Cajalin väliytimeen samalla puolella sekä Darkshevichin ytimeen ja silmän motorisen ytimen rastraaliseen osaan molemmilta puolilta.
Erilaiset etuosan näkökuoren alueet ohjaavat sakkadisia silmien liikkeitä. eri amplitudi, sekä sakkadien suunta. Ne tarjoavat niin sanotun "visuaalisen huomion" ohjaten katseet "tavoitteeseen".
Asiaa tutkitaan parhaillaan otsalohkon rooli silmäluomien toiminnassa. Silmäluomien vapaaehtoinen sulkeminen edellyttää nostoluomen rentoutumista ja orbicularis oculi -lihaksen supistumista. Silmäluomen tahdonvoimaisia liikkeitä ohjaavien aivojen alueiden sijainnista on kiistaa. Nämä paikat voivat sijaita moottorin esimoottorin tai etumoottorin näköalueilla. Tämän alueen stimulointi johtaa vastakkaiseen silmäluomien sulkeutumiseen. Ei-dominanttien etulohkojen yksipuolinen vaurio voi liittyä molemminpuoliseen silmäluomien sulkeutumiseen.
Brocan motorinen puhealue(kentät 44 ja 45 Brodmanin mukaan).
Tämä alue sijaitsee etulohkon taka-alaosassa. Hän osallistuu puheen aloittamiseen. Brocan alueen vaurioituminen johtaa afasiaan, joka koostuu vaikeudesta yhdistää tiettyyn sanasarjaan liittyvää puhetta (ekspressiivinen afasia). Potilaat ymmärtävät sekä kirjoitetun että puhutun kielen, mutta eivät yleensä osaa ääntää mitään. Myös kirjoituskyky on heikentynyt, vaikka kättä voidaan käyttää muuhun toimintaan.
frontaalinen assosiaatioalue. Tämä etulohkon etupuolen tärkeä alue on vastuussa kaikenlaisten käyttäytymis-älyllisten, sisäelinten ja tunnetoimintojen käynnistämisestä. Muutoksia muistissa, älykkyydessä ja persoonallisuudessa ilmenee otsalohkojen ärsytyksen tai massiivisen tuhoutumisen seurauksena. Tämä on erityisen voimakasta näissä tapauksissa. kun hallitseva aivopuolisko vaikuttaa.
Hajukanava sijaitsee otsalohkon orbitaalipinnan alapuolella ja on ainoa aistinreitti, joka saavuttaa aivokuoren ilman synapsien muodostumista talamuksessa. Otsalohkon orbitaalipinnan yksipuolinen kasvain (olfactory sulcus meningioma tai alasiiven meningioma) sphenoidinen luu) voi johtaa hajukanavan puristumiseen ja optinen hermo. Seurauksena kehittyy anosmia (hajupuute), ipsilateraalinen näköhermon surkastuminen ja näköhermon pään turvotus vastakkaisella puolella (Kennedy Fosterin oireyhtymä).
Etulohkon limbisen järjestelmän rakenteiden vaurioituminen, erityisesti gyrus, voi aiheuttaa häiriöitä elämän tunnepuolella sekä sisäelinten merkkejä, mukaan lukien syytön kyynelvuoto ja muutoksia pupillien vasteessa valoärsykkeisiin.
parietaalinen lohko(katso kuva 4.1.12, 4.1.13). Yleensä parietaalilohkojen toiminta liittyy herkkyyteen, puheeseen ja oppimiseen liittyvään muistiin sekä kehon suuntautumiseen avaruudessa visuaalisen tiedon käsittelyn kautta.
Etummaisessa parietaalilohkossa on gyrus, joka sijaitsee yhdensuuntainen precentraalisen gyrusin tai motorisen aivokuoren kanssa. Tämä on postcentral gyrus eli primaarinen somatosensorinen aivokuori (kentät 3, 1 ja 2) (katso kuva 4.1.13). Loput parietaalilohkosta voidaan jakaa kahteen lohkoon - ylä- ja alalohkoon. Kentät 5 ja 7, jotka sijaitsevat ylemmän parietaalilohkon pinnalla, liittyvät somatosensorisen tiedon korrelaatioon, mikä mahdollistaa esineen painon, pinnan ominaisuuksien, koon ja muodon tietoisen arvioinnin.
Alempi parietaalinen lobule(supramarginaalinen ja kulmikas gyrus) liittyy läheisesti puhemekanismeihin, koska tämän aivokuoren alueen tuhoutuminen vasemmalla pallonpuoliskolla johtaa puheen havaitsemisen heikkenemiseen.
Parietaalilohkon roolia avaruudellisessa orientaatiossa ei ole vielä tutkittu. Kuitenkin potilailla, joilla on tappio, havaitaan tämän toiminnon rikkominen.
Afferentit kuidut kulkevat parietaalilohkon läpi yhdistäen alueet 18 ja 19 otsalohkoihin ja aivorunkoon. Nämä reitit sisältävät ylemmät ja alemmat pitkittäiset niput.
Oletetaan, että parietaalinen aivokuori (kenttä 7) tarjoaa "huomio"-toiminnon visuaalisiin kohteisiin.
Seitsemäs kenttä vastaanottaa afferenteja cingulate gyrus(g. cinguli), esiperäiseltä alueelta ja quadrigeminan ylätuberkuloista talamuksen ytimien kautta. Löytyy myös afferentteja, jotka kulkevat etuosan näköalueelta ja esikuoresta parietaalilohkon aivokuoreen.
Efferentit projisoidaan preektaaliselle alueelle, quadrigeminan ylätuberkuloihin, sylvian vesijohdon ympärillä sijaitsevaan harmaaseen aineeseen ja aivojen kahden pallonpuoliskon parietaalilohkoihin.
Kun silmät liikkuvat, alemman parietaalilohkon hermosolut aktivoituvat, mikä osoittaa niiden roolin tahdonalaisten silmän liikkeiden ohjauksessa. Toiminta jatkuu kohteen havainnoinnin ajan.
Seitsemännen kentän solut eivät määritä sakkadisia liikkeitä, koska niiden aktiivisuus ei kasva sakkadien aikana.
Angular gyrusin stimulaatio(g. angularis) johtaa koe-eläimillä silmien ystävälliseen poikkeamiseen. Nämä ovat kentät 38 ja 39, jotka kulkevat keskimmäisen temporaalisen gyrusen (g. temporalis medius) takareunan ympäri. Operatiivinen poisto tämä aivokuoren alue ei johda silmien liikkeen heikkenemiseen. Parietaalikuoren molemminpuolinen vaurio voi johtaa silmän motorisen apraksian kehittymiseen.
Parietaalilohkon tappioon liittyy myös optokineettisen nystagman aikaansaavien visuaalisten motoristen mekanismien rikkominen, jonka aikana silmät tuottavat sakkadeja kohteen etsimiseksi ja kiinnittämiseksi.
Muita eristetyn parietaalilohkon vaurioiden motorisia merkkejä ovat silmien ystävällinen poikkeama vastakkaiseen suuntaan vammapaikasta silmäluomien pakkosulkeminen, potilaan vähäinen liikkuvuus ja vastakkaisen puolen raajojen huono hallinta (väärinkäsitys). Vaikeudet ymmärtää raajojen toiminnan luonnetta johtavat perustuslailliseen ja okulomotoriseen apraksiaan. Potilailla, joilla on vaurioita parietaalilohkossa, voi olla vaikeuksia siirtää katseensa yhdestä näkökohteesta toiseen näkökentän kontralateraalisella puoliskolla.
Toinen merkki parietaalilohkon toimintahäiriöstä on kyvyttömyys korjata visuaalista objektia.
Sensoristen toimintojen lisäksi, jotka sijaitsevat Rolandon keskussuluksen takahuulessa, parietaalilohko on monimutkaisen tunnistus- ja ymmärtämisprosessin paikka. Parietaalilohkojen diffuusi vaurio päättyy agnosiaan, näkökyvyttömyyteen, metamorfopsiaan, fotopsiaan, hallusinaatioihin, illuusioihin ja aleksiaan (ks. kuva 4.1.12, 4.1.13).
Lisää yksityiskohdat parietaalilohkon toimintahäiriön seurauksista on annettu neurologisessa kirjallisuudessa. Tässä esittelemme vain osan oireyhtymistä.
visuaalinen agnosia, joka kehittyy usein keskushermoston verenkierron vastaisesti, on tunnusomaista tunnistusprosessin rikkomisesta. Agnosia voi olla täydellinen tai osittainen. Visuaalinen agnosia diagnosoidaan tapauksissa, joissa potilas voi tunnistaa kohteen vain kosketusti. Visuaalisten kuvien ja kuvan värimerkityksen tunnistaminen ja ymmärtäminen heikkenevät usein.
Parietaalilohkon molemminpuolisiin vaurioihin (yleisempi seniilissä dementiassa, aivokuoren temporo-okcipitaalisten ja parietaalisten alueiden pehmeneminen, molempien aivopuoliskon takaosien vammat, enkefaliitti) liittyy visuaalinen agnosia ja visuaalinen ataksia (Balintin oireyhtymä).
Visuaalinen spatiaalinen agnosia jolle on ominaista vaikeuksia arvioida kohteen kokoa ja etäisyyttä siihen. Myös "synteettisten" spatiaalisten ongelmien, kuten kartan lukemisen, väärän ratkaisun ilmaantuminen on havaittu. Samanaikainen agnosia tiivistyy siihen tosiasiaan, että potilaat säilyttävät kyvyn tunnistaa yksittäisiä esineitä, mutta heillä ei ole kykyä havaita esineryhmää (kuvia) kokonaisuutena.
Asterognostia jolle on ominaista se, että potilas osaa analysoida esineitä niiden ulkonäön perusteella, mutta ei pysty karakterisoimaan niitä kosketusti. Tämä kehittyy aivokuoren parietaalilohkoissa koskettavan ja visuaalisen tiedon välisen monimutkaisen vuorovaikutuksen katkeamisen seurauksena.
Parietaalilohkojen diffuusi vauriot voivat johtaa illuusioita ja metamorfopsioita. Näihin vääristymiin kuuluvat mikropsia, macropsia. Potilaat huomaavat paikallaan olevien kohteiden liikkeen, arvioivat väärin etäisyyden esineisiin. TO erikoistyyppejä illuusioita ovat papinopsia (visuaalisen havainnon rikkominen visuaalisen kuvan säilymisen tai uudelleen ilmestymisen muodossa esineen katoamisen jälkeen näkökentästä), hallusinatorinen polyopia jne.
Illuusiot kehittyvät aivokuoren diffuusien häiriöiden seurauksena, ja hallusinaatiot ja fotopsiat kehittyvät kasvaimen tai toksisen aivokuoren vaurion kehittyessä.
Parietaalilohkon patologian ominaisuudet voivat myös riippua siitä, onko patologinen fokus paikallinen hallitsevassa (yleensä vasemmassa) vai ei-dominoivassa pallonpuoliskossa. Dominoiva parietaalilohko on vastuussa puhehäiriöiden, kuten kuuloaistin afasia, kehittymisestä, visuaalinen agnosia ja agrafioita.
Vaurioitunut hallitseva kulmikas gyrus(useammin verenkiertohäiriöiden seurauksena kulmavaltimon altaassa) kehittyy nystagmus, näkö-agnostiset häiriöt, agrafia, acalculia (kyvyttömyys lisätä tai vähentää lukuja), digitaalinen agnosia, kyvyttömyys erottaa oikea ja vasen puoli ( Gerstmannin oireyhtymä; angular gyrus -oireyhtymä). Ehkä homonyymisen hemianopsian kehittyminen.
Jos ei-dominoiva kulmakivi vaurioituu, potilaan vuorovaikutus ympäristön kanssa häiriintyy. Tämän seurauksena visuaalinen disorientaatio ja topografinen agnosia kehittyy.
Anterior angular gyrus ja poikki supratemporaalinen sulcus on supramarginaalinen gyrus. Dominoivalla pallonpuoliskolla tämä alue on kielen visuaalisten ja kuulokomponenttien kohtauspaikka.
Takaosan parietaalisen assosiaatioalueen vauriot päättyvät usein apraksian kehittyminen(vaikeus suorittaa "hienoja" erikoisliikkeitä). Tämä johtuu ilmeisesti väärinymmärryksestä tietyn toiminnon suorittamiseen tarvittavasta liikesarjasta (eli potilaalle ei ole selvää, mitä ja missä järjestyksessä pitäisi tehdä). Potilaalta puuttuu kyky piirtää yksinkertaista kaaviota (konstruktiivinen apraksia). Tämän patologian vähemmän vakava muoto on kosketusagnosia (kyvyttömyys tunnistaa kohdetta tunnustelussa). Joskus tätä sairautta kutsutaan asterognostiaksi. Samaan aikaan ei ole merkkejä tuntoherkkyyden tai proprioseptiivisen herkkyyden menetyksestä, ja visuaalisen ja somatosensorisen tiedon integraatio on heikentynyt. Apraxia ja asterognostia kehittyvät usein aivojen vasemman pallonpuoliskon vaurioitumisen jälkeen. Asterognostia rajoittuu yleensä kehon vastakkaiselle puolelle, kun taas apraksia on yleensä kahdenvälinen.
Visuaalinen ja auditiivinen informaatio liittyy kiinteästi kielen toimintaan. Tästä syystä alueiden 39 ja 40 sekä kuuloassosiaatioalueen osien vauriot johtavat yleensä afasiaan, ns. afasia Wernicke(Wernicke). Jos vaurio rajoittuu kulmakiveen, kyky ymmärtää kirjoitettua kieltä (alexia) ja kyky kirjoittaa (agraphia) menetetään. Samalla potilaat ymmärtävät puhetta. Tämä tila kehittyy yleensä vamman jälkeen.
Visuaalisen ja somatosensorisen tiedon yhdistäminen on tärkeää "kehokuvan" muodostumiselle, eli kehon asennon piirteiden ymmärtämiselle avaruudessa. Yhden pallonpuoliskon kenttien 7, 39 ja 40 laajat vauriot voivat johtaa kehon vastakkaisen puoliskon "väärinkäsitysten" tai "laiminlyöntien" kehittymiseen. Potilaat eivät pysty sitomaan tai irrottamaan omia raajojaan. He eivät myöskään tunnista omia raajojaan.
Okcipital lohko(kentät 17, 18 ja 19). Kannusuraa pitkin (fissura calcarina) on ensisijainen näköalue (kenttä 17). Sen vieressä on "arvioiva" visuaalinen alue (kenttä 18), jonka uskotaan muuntavan kentän 17 vastaanottamat signaalit visuaalisiksi kuviksi.
Okcipital cortexin alue 17 on visuaalisen tiedon vastaanotto- ja käsittelyalue ja on tärkeitä yhteyksiä aivokuoren motorisilla visuaalisilla alueilla. Näkyvän aivokuoren tällä tasolla arvioidaan sen merkitystä, mitä on nähty. Tämän alueen ärsytys aiheuttaa hallusinaatioita ja unenomaisia kuvia.
Myös takaraivolohkon motoriset yhteydet ansaitsevat huomiota, koska ne ovat mukana optokineettisessä nystagmissa, akkomodaatiorefleksissä ja kiikarin näön säätelyssä.
Kentät 18 ja 19 ovat toisistaan riippuvaisia ja niillä on enemmän yhteyksiä muihin aivoihin. Uskotaan, että alue 18 on enemmän kiinnostunut visuaalisen tiedon integroinnista. kentässä 17, kun taas kenttä 19 muuttaa tämän tiedon monimutkaisemmiksi henkiseksi ja motoriseksi toiminnaksi.
Viimeaikaiset anatomiset ja fysiologinen tutkimus paljasti, että niitä on vähintään, 10 ala-aluetta kentissä 18 ja 19, joiden hermosoluilla on kyky vastata visuaalisiin ärsykkeisiin. Jokainen näistä osa-alueista liittyy 17. kenttään. Ne eivät kuitenkaan liity toisiinsa. Näiden osa-alueiden vaurioituminen ei johda sokeuteen. Joitakin rikotaan visuaaliset toiminnot, kuten värianalyysi, kohteen eheyden havaitseminen jne.
visuaalinen assosiaatio aivokuori, joka sijaitsee joillakin ohimolohkon alueilla, osallistuu myös visuaalisen tiedon käsittelyyn. Tämä pätee suurimmassa määrin kenttää 21 (keskimäinen temporaalinen aivokuori). Juuri tämä kenttä tunnistaa kuvan, ja sen vauriot päättyvät agnosiaan.
Yllä olevat näkökuoren ominaisuudet saadaan myös lukuisista vaaka- ja pystysuorista yhteyksistä neuronien välillä näkökuoren eri kerroksissa. Niiden läsnäolon ansiosta visuaalisen aivokuoren poikkeuksellinen plastisuus havaitaan erityyppisen visuaalisen tiedon käsittelyssä.
Näkökuoren rakenteellinen organisaatio ja sen tehtävät selostetaan tarkemmin seuraavassa osiossa.
ajallinen lohko(kentät 41, 22).
Tämä aivojen osa liittyy kuuloaistimusten havaitsemiseen, osallistuu puhetoimintoon puheen auditiivisen ohjauksen kautta, ja sillä on myös rooli tilan ja muistin toiminnan arvioinnissa. On pitkään todettu, että poikittaissuuntainen temporaalinen gyrus (kenttä 41) on ensisijainen kuulokeskus. Pieni kenttä (22). tätä gyrusta ympäröivää, pidetään "henkisenä" tai "arvioivana", kuulokeskus. Useimpien ohimolohkon ärsytys, erityisesti keskimmäisessä ohimolohkossa, aiheuttaa aistimuksia, joita potilas kuvailee kuuloksi. Potilaat, joilla on vaurioita ohimolohkon "henkisessä kentässä", menettävät joskus kyvyn arvioida ääniä. Samaan aikaan potilas ei tunnista motiiveja, ja hän näkee musiikilliset äänet satunnaisena meluna.
Vasemman pallonpuoliskon kenttä 22 liittyy myös puhetoimintoon, koska tämän alueen vaurioituminen johtaa sanojen merkityksen ymmärtämisen menettämiseen. On myös näyttöä siitä, että ohimolohko liittyy "vestibulaariseen" herkkyyteen (tasapainon tunteeseen), koska tajuissaan olevan potilaan ylemmän ohimokehän stimulaatio aiheuttaa huimausta ja pyörimistunnetta.
Ohimolohkon syviin vaurioihin liittyy usein alhaisimmat optisen säteilyn kuidut lateraalisesta geniculate-kehosta. Jossa heikkonäköinen. Yleisin on ylemmän neljänneksen hemianopia.
Kun tappio temporaalinen tai psykomotorinen lohko kehittyy epilepsia, johon liittyy visuaalisia hallusinaatioita, deja vu ja deja pense ("jo nähty" - fr.), jossa visuaalisella kokemuksella ja ajatuksella on outo ystävällinen suhde, ilmaantuu automatismeja, jotka seuraavat muistinmenetystä, pakkokäyttäytymistä, vihaa ja ahdistusta.
Potilailla, joilla on yksipuolinen vaurio ensisijassa kuulokorteksti lievä kuulonalenema. Kuulotunnistuksen vastaisesti (auditiivinen agnosia) vaikuttaa assosiatiivisiin kuulokentoihin. Potilaat ilmoittavat yleensä kuulevansa ääniä, mutta eivät voi tunnistaa niitä.
Kasvojen tunnistamiseen osallistuva alue löytyy ohimolohkon vatsapinnalta. Tämän alueen vaurio (useammin häiritsee verenkiertoa aivojen tyvivaltimon alueella), joka yleensä ulottuu viereiseen takaraivolohkoon (kentät 20, 21), molemmille puolipalloille, mukana prosopagnosian kehittyminen(kyvyttömyys tunnistaa kasvoja). Yleensä tähän tilaan ei liity muita neurologisia oireita. Potilas osaa lukea ja nimetä esineitä. Tapauksissa, joissa potilas kuulee tutun henkilön äänen, hän voi tunnistaa hänet välittömästi.
Jäljelle jäävä osa ylemmän temporaalisen gyrusen miehittää alue 22 (auditory assosiatiivinen aivokuori), joka vastaanottaa suuren määrän afferentteja kentistä 41 ja 42 antaakseen kuituja parietaali- ja takaraivolohkoon. Alue 22 osallistuu myös kielitoimintoihin. 22. kentän yläosan vaurioituminen johtaa vaikeuksiin kielen ymmärtämisessä.
Ohimolohkot ovat erityisen tärkeitä muistitoiminnassa.. Esimerkiksi kuuloassosiaatiokuoren stimulaatio potilailla neurokirurgisten leikkausten aikana herättää monimutkaisia muistoja, sekä kuulo- että visuaalisia. Ohimolohkon laajoissa vaurioissa ilmenee muistin heikkenemistä.
limbinen lohko(Kuva 4.1.14).
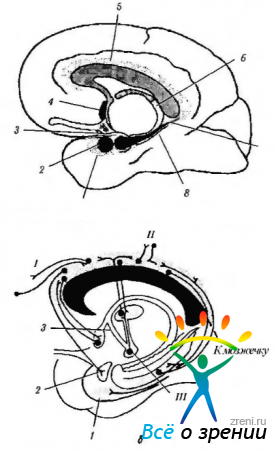
Riisi. 4.1.14. Limbinen järjestelmä (Brodslin, 1992 mukaan): a - limbisen järjestelmän rakenteiden sijainti (1 - entoriaalinen kenttä; 2 - amygdala; 3 - koukku; 4 - väliseinän ydin; 5 - singulaate gyrus; b - fornix; 7 - hippokampus; 8 - parahippokampus gyrus); b - cingulaattisen gyrusen liitännät (I - etuosan assosiatiivinen aivokuori; II - parietaalinen ja ajallinen assosiatiivinen aivokuori; III - mastoidirunko). Cingulate gyrus on yhteydessä aivokuoren ja limbisen järjestelmän assosiatiivisiin kenttiin, mikä tarjoaa niiden välistä vuorovaikutusta.
Limbinen järjestelmä on terminaalin, väliaivojen ja keskiaivojen muodostumien kompleksi. Itse asiassa " limbinen järjestelmä” sopii yhteen aiemmin käytetyn käsitteen kanssa - "hajuaivot".
Jos uusi aivokuori (neocortex) hallitsee organismin tila-ajallisia suhteita ympäristöön ja on myös vastuussa muodollis-loogisesta ajattelusta ja stereognostisista kyvyistä, niin limbinen järjestelmä määrittää tunnetoiminnot ja toimintamotivaation sekä oppimis- ja muistiprosessit. Se säätelee myös homeostaasia. Sen tärkeä tehtävä on yksilön käyttäytymisreaktioiden järjestäminen vastauksena ulkoisen ympäristön vaikutuksiin ja kehon sisäisen ympäristön muutoksiin. Nämä reaktiot tähtäävät yksilön säilyttämiseen ja ne välittyvät aivokalvon alempien keskusten kautta.
Limbisellä lohkolla on myös jotain tekemistä hajun toiminnan kanssa, koska aivotursoalueen ärsytykseen tai vaurioitumiseen liittyy hajuaisti.
Limbinen lohko muodostuu:
- subcausaalinen alue (alue subcallosa),
- cingulate gyrus (gyrus cinguli),
- cingulate gyrus -kanta
- parahippocampal gyrus (gyrus parahippocampal),
- hippokampus (ammonin sarvi, hampaita ja hippokampuksen tyvi eli subiculum)
- ja Brocan diagonaalinen nivelside.
Limbisen järjestelmän subkortikaalisia rakenteita ovat risat (corpus amygdaloideum), väliseinän ytimet ja anteriorinen talamuksen tuma.
Limbisen järjestelmän rakenteiden afferentit ja efferentit yhteydet sekä keskenään että muiden aivojen osien kanssa ovat erittäin monipuolisia. Sen vastavuoroiset yhteydet hypotalamuksen kanssa ovat selkeimpiä.
Hypotalamus ja maitorauhaset yhdistetty hippokampukseen ja väliseinän alueelle fornixin kautta, amygdalaan terminaalisen strian ja amygdalofugal-kimpun kautta ja hajuaivoihin mediaalisen nipun kautta etuaivot. Limbinen järjestelmä on yhteydessä keskiaivoon hypotalamuksen ja rintamaidon kautta.
Limbinen järjestelmä kommunikoi neokorteksin kanssa frontaalisessa ja ajallinen lohko. Ajalliset alueet ovat pääasiassa vastuussa tiedon välittämisestä näkö-, kuulo- ja somaattisesta aivokuoresta amygdalaan ja hippokampukseen. Frontaaliset alueet toimivat neokorteksin pääjakoina, jotka säätelevät limbisiä vyöhykkeitä.
Tyvitumake edustavat subkortikaalista hermosolujen kertymää tumaryhmän muodossa (kuva 4.1.15; 4.1.16).
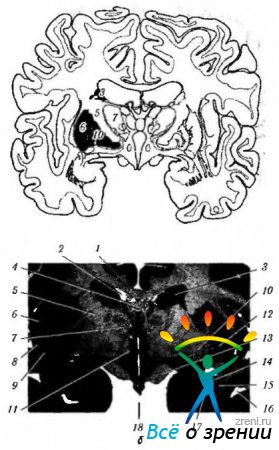
Riisi. 4.1.15. Tyvitumake: a - kaavamainen esitys tyviganglioihin liittyvien rakenteiden sijainnista; b - aivojen etuosa, joka kulkee tyviganglioihin liittyvien rakenteiden läpi (4 - corpus callosum; 2 - sarvi sivukammio; 3- häntäytimen runko; 4-holvi; 5-sisäkapseli; 6 - saareke; 7 - visuaalinen tuberkuloosi; 8- kuori; 9 - aita; 10 - vaalea pallo; 11 - hypotuberous alue; 12 - nimetön aine: 13 - näkökanava; 14 - hännän ytimen takaosa; 15 - risat: 16 - sivukammion temporaalinen sarvi; 17 - sivukammion etusarvi; 18 - kolmas kammio)
Riisi. 4.1.16. Tyviganglioiden tärkeimmät afferentit, efferentit ja sisäiset yhteydet ja niiden yhteys pikkuaivoon (selitys tekstissä): a - (1-motorinen aivokuori; 2 - efferentit kuidut aivokuoresta striatumiin; 3 - aivojuovio; 4 - vaalea pallo; 5 - afferentit kuidut; 6 - visuaalinen tuberkuloosi; 7 - subtalaminen ydin; 8 - substantia nigra); b - (1 - aivokuori (motorinen); 2 - tyvihermot; 3 - pikkuaivot; 4 - motoriset ja interkalaariset neuronit)
Ne ohjaavat kehon liikettä. Tämä tyviganglioiden toiminta määritettiin kliinisten havaintojen perusteella. Kun ne ovat vaurioituneet, kyseessä on rikkomus motorista toimintaa sekä kyvyttömyys käynnistää tarvittavia liikkeitä, että kyvyttömyys tukahduttaa liikkeitä.
Parkinsonin taudissa, joka ilmenee, kun tyvigangliot ovat vaurioituneet, potilailla on "ilmaisuttomat" kasvot. Tämä liittyy kasvolihasten heikentyneeseen motoriseen toimintaan ja silmien liikkeiden hallintaan, erityisesti sakkadisten liikkeiden häiriöihin. Tästä syystä tarkastelemme lähemmin tämän alueen neuroanatomiaa.
Niitä on kolme pääasiallista aivokuoren ytimien kertymät, joita kutsutaan striatumiksi ( corpus striatum), aita (claustrum) ja amygdala (corpus amigdaloideum).
- Corpus striatum koostuu kahdesta osasta - caudate-ytimestä (nucleus caudatus) ja linssimuotoisesta ytimestä (nucleus lentiformis).
Caudate ydin sijaitsee linssimäisen ytimen yläpuolella ja mediaalisesti, ja sen erottaa jälkimmäisestä valkoisen aineen kerros, jota kutsutaan sisäiseksi kapseliksi (capsula interna). Häntäytimen paksuuntunut etuosa, sen pää (caput nuclei caudati) muodostaa sivuseinän etusarvi sivukammio, kun taas takakammion ohut osa (corpus et cauda nuclei caudati) ulottuu taaksepäin sivukammion keskiosan pohjaa pitkin. Mediaalisella puolella nucleus caudatus on visuaalisen kasan vieressä, erotettuna siitä valkoisen aineen kaistaleella (stria terminalis). Edessä ja alempana häntäytimen pää saavuttaa etummaisen rei'itetyn aineen (substantia perforata anterior). Tässä vaiheessa pää on yhdistetty linssimäiseen ytimeen (osalla, jota kutsutaan putameniksi). Tämän molempien ytimien leveän yhteyden lisäksi vatsan puolella on myös ohuita harmaan aineen kaistaleita, joiden välissä on sisäkapselin valkoisia kimppuja. Niistä syntyi nimi "juovainen ruumis" (corpus striatum).
Linssimainen ydin(nucleus lentiformis) sijaitsee sivusuunnassa hännän ytimestä ja visuaalisesta kumpusta, erotettuna niistä sisäisellä kapselilla. Puolipallon vaakaleikkauksella linssimuotoisen ytimen keskipinta, joka on sisäkapselia päin, on haurauden muotoinen, jonka kärki on suunnattu keskelle. Kulman etupuoli on yhdensuuntainen häntäytimen kanssa ja takapuoli on yhdensuuntainen talamuksen kanssa. Sivupinta on hieman kupera ja on kohti puolipallon lateraalista puolta insulan alueella. Anteriorisesti ja ventraalisesti, kuten jo osoitettiin, linssimuotoinen ydin sulautuu häntäytimen pään kanssa.
Frontaaliosassa linssimuotoinen ydin on kiilan muotoinen, jonka kärki on käännetty mediaaliselle puolelle ja pohja on lateraalisesti. Linssimainen ydin on jaettu kolmeen segmenttiin kahdella yhdensuuntaisella valkoisella kerroksella (laminae medullares). Sivusuunnassa olevaa tummanharmaata aluetta kutsutaan kuoreksi (putamen), ja kahta keskimmäistä, vaaleampaa aluetta yhdessä kutsutaan vaaleaksi palloksi (globus pallidus).
Se eroaa jo makroskooppisen ulkonäöltään, vaalea pallo(globus pallidus) on myös histologinen rakenne, joka eroaa merkittävästi aivojuovion muiden osien rakenteesta.
Kaikkien näiden ominaisuuksien perusteella vaalea pallo erottuu erityiseksi morfologiseksi yksiköksi nimeltä pallidum, kun taas nimitys striatum jätetään vain putamenille ja nucleus caudatusille. Tämän seurauksena termi "linssimainen ydin" menettää entisen merkityksensä ja sitä voidaan käyttää vain puhtaasti topografisessa merkityksessä. Samanaikaisesti entisen nimen corpus striatum sijasta häntä- ja linssimäisiä ytimiä kutsutaan striopallidar-järjestelmäksi.
Striopallidary järjestelmä on pääosa ekstrapyramidaalista järjestelmää ja lisäksi se on korkein säätelykeskus autonomiset toiminnot suhteessa lämpösäätelyyn ja hiilihydraattien aineenvaihduntaan, hallitseva samankaltaisia vegetatiivisia keskuksia hypotalamuksessa.
- Aita(claustrum) on ohut harmaata ainetta, joka on asetettu saaren alueelle, sen ja putamenin väliin. Sen erottaa jälkimmäisestä valkoisen aineen kerros, jota kutsutaan ulkokapseliksi (capsula externa).
- amygdala(corpus amygdaloideum) sijaitsee putamenin alla ohimolohkon anteriorisessa päässä. Amygdala näyttää kuuluvan subkortikaalisiin hajukeskuksiin ja limbiseen järjestelmään. Se päättyy hajulohkosta tulevaan kuitukimppuun ja etummainen rei'itetty substanssi (substantia perforata anterior), joka on mainittu visuaalisen kasan kuvauksessa nimeltä stria, päättyy.
Amygdalan roolia ihmisissä ei ymmärretä hyvin. Pohjimmiltaan amygdalan rooli paljastettiin neurologisten potilaiden tutkimuksen perusteella. Näiden tutkimusten ansiosta on todettu, että amygdalan mekaaninen tai sähköinen stimulaatio ihmisillä johtaa usein pelkoon tai muihin tunnereaktioihin. Näin ollen ihmisen amygdala osallistuu tunnehavaintoprosesseihin. On myös osoitettu, että risojen kahdenvälisillä vaurioilla esiintyy "emotionaalisen" muistin menetystä, "pelästyneen" ilmeen ilmaantumista ja haju- ja visuaalisen tiedon integrointia.
On hyvin tunnettua, että henkilöllä on kyky määrittää toisen henkilön mieliala ja tunnetila. SISÄÄN Viime aikoina amygdalan osallistuminen tähän kompleksiin fysiologinen prosessi. Määritettäessä ihmisen tunnetilaa aivomme taltioivat ja analysoivat hienovaraisia muutoksia kasvojen ilmeissä ja ennen kaikkea katseen suunnassa. Näiden prosessien taustalla olevat hermomekanismit eivät ole täysin selviä. Siitä huolimatta on todettu, että "tarkka" katseen asettaminen, psykofysiologisesti arvioituna "huomioon" päättänyt alkuun ajallinen aivokuori ja amygdala. Autismia sairastavien potilaiden tutkimuksessa paljastettiin, että heillä on usein amygdala tai sen vaurio. Toisten ihmisten katseen suuntautumiseen perustuvan sosiaalisen arvioinnin taustalla olevat välittömät mekanismit ovat suurelta osin tuntemattomia.
Viime aikoina mustasubstantia ja subtalamuksen ytimiä on viitattu myös tyvihermoille. Lisäksi substantia nigrassa erotetaan 2 osaa - retikulaarinen (pars reticulata) ja kompakti (pars compacta).
Morfologiset ja elektrofysiologiset menetelmät ovat osoittaneet sen suurin osa basaaliganglioihin tulevat afferentit signaalit tulevat striatumiin (ks. kuva 4.1.16). Nämä signaalit tulevat monista lähteistä, joista tärkeimmät ovat:
- kaikki aivokuoren alueet pallonpuoliskot;
- talamuksen intralamellaariset tumat
- substantia nigra (dopaminergisen reitin varrella).
Vaalean maapallon sisältä on peräisin kaikista tärkein efferenttejä polkuja tyvitumake. Tämä polku päättyy pääasiassa visuaaliseen tuberkuloosiin ja väliaivojen kattoon.
Siten tyviganglionilla on välilenkin rooli ketjussa, joka yhdistää aivokuoren motoriset alueet kaikkiin muihin alueisiin. Niiden päätehtävä on fyysisen toiminnan "suunnittelua"..
SISÄÄN viime vuodet havaitsi, että tyvigangliot yhdessä joidenkin aivokuoren alueiden kanssa määrittävät myös joitakin kognitiiviset toiminnot, mukaan lukien huomio, muisti. Käsite "huomio" sanan laajimmassa merkityksessä on "valintaprosessi". Ottaen huomioon, että avaruudessa suuntautumisprosessi ja kiinnostuksen kohteena olevan tavoitteen valinta, johon sakkadiset silmäliikkeet osallistuvat, liittyy tyviganglioniin, tulee selväksi basaalikompleksin merkittävä rooli sakkadien läpi suuntautumisprosesseissa. Tässä tapauksessa erityinen paikka silmän sakkadien hallinnassa on substantia nigralla, erityisesti sen retikulaarisella osalla. Sekä anatomisesti että elektrofysiologisesti sen yhteys quadrigeminan ylempien tuberkuloosien kanssa on todistettu. Substantia nigran hermosolujen päätehtävä on estää sakkadien muodostumiseen osallistuvien quadrigeminan ylemmän colliculuksen hermosolujen aktiivisuutta.
Silmien motorisessa toiminnassa mukana on myös häntäydin, jolla on yhteydet substantia nigraan ja quadrigeminan ulompiin tuberculeisiin. Häntäytimen viritys johtaa silmän sakkadisiin liikkeisiin vastakkaiseen suuntaan kuin virittynyt tuma.
Subtalaminen ydin on myös mukana silmien liikkeissä. Vaalea pallo, etuosan aivokuori, projisoidaan tähän ytimeen. Kuidut lähtevät subtalamisesta ytimestä substantia nigraan, globus pallidus -kalvon ulko- ja sisäosaan.
Globus palliduksen tehtävä silmän motorisessa aktiivisuudessa on vähemmän selvä. Tämä johtuu siitä, että globus palliduksella on suuri määrä yhteyksiä tyviganglioiden eri ytimien välillä ja vähän yhteyksiä hermokeskukset sijaitsee basaaliganglioiden ulkopuolella. Silmänmotorisia hermosoluja löydettiin vain globus palliduksen dorsaalisesta osasta, eli siitä osasta, joka vastaanottaa syötteitä häntäytimestä.
Tyviganglioiden rooli silmien liikkeessä lukuisat kliiniset havainnot vahvistavat. Parkinsonin taudissa kehittyy progressiivinen supranukleaarinen halvaus, johon liittyy väliaivojen tyviganglioiden ja harmaan aineen rappeutuminen ja frontomesencefaalisten reittien katkaisu. Tällaisilla potilailla silmän sakkadisten liikkeiden nopeus vähenee. Sakkadit ovat yleensä hypometrisiä, mikä liittyy "sakkadimuisti" -toiminnon rikkomiseen. Usein silmän liikettä ei voida suorittaa kokonaan. Pystysuorat silmien liikkeet vaikuttavat enemmän kuin vaakasuuntaiset. Potilailla on myös aksiaalista jäykkyyttä ja dementiaa. Samanlaisia muutoksia havaittiin myös Huntingtonin taudissa.
corpus callosum(corpus callosum). Corpus callosum on suuri kuitukimppu (joka koostuu noin 250 miljoonasta kuidusta), joka yhdistää molemmat pallonpuoliskot (kuva 4.1.17).
Riisi. 4.1.17. Talamuksen ytimien lokalisointi: a - ydinparamagneettinen resonanssi; b - aivojen etuosa (1 - corpus callosumin runko; 2 - hännän ydin; 3 - sivukammion keskiosa; 4 - läpinäkyvä väliseinä; 5 - suonikalvon plexus sivukammio; 6 - holvi; 7 - visuaalisen kasan ytimien eturyhmä; 8- ulompi kapseli; 9 - talamuksen ytimien lateraalinen ryhmä; 10- talamuksen mediaaliset ytimet: - kuori; 12- lateraalinen ydinlevy; 13 - sisäkapseli; 14 - talamuksen retikulaarinen ydin; 15 - intertalaminen piikki; 16 - vaalean pallon sivuosa; 17 - mediaalinen medullarilevy; 18 - mammillotalaminen nippu; 19 - HI Taimenkenttä; 20 - gopa incerta; 21 - H2 Taimenkenttä; 22 - aita; vaalean pallon 23-mediaaalinen osa; 24 - kolmas kammio; 25 - hypotalamuksen ytimet; 26-näöntie; 27 - amygdala; 28-nännin runko; aivojen jalkojen 29 tyvestä)
Tämän säteen päätehtävä on tiedon välittäminen pallonpuoliskolta toiselle.
Corpus callosumin päätehtävät ovat:
- Kuvien korrelaatio näkökentän oikealla ja vasemmalla puoliskolla.
- Parillisista raajoista tulevien tunteiden integrointi, välttämätön motorisen koordinaation oppimiseen.
- Huomio- ja aktivointiprosessien integrointi pallonpuoliskolla.
Potilailla, joille corpus callosum leikattiin läpi terapeuttisia tarkoituksia varten, aivojen toimintahäiriö Jokapäiväinen elämä ei havaittu. Ne paljastettiin vasta läpi erityisiä testejä. Toisille potilaille kehittyy joskus varsin merkittäviä käyttäytymishäiriöitä. Tämä ilmenee useimmiten "verbaalisella sokeudella", "sanallisella kuuroudella" ja oikean ja vasemman raajan liikkeiden koordinoinnin menetyksellä.
Jatkuu seuraavassa artikkelissa: Aivojen anatomia? Osa 3
Ja koko pääkaupungin silmissä
Kukko lepahti neulepuikasta,
lensi vaunuihin
Ja istui kuninkaan kruunuun,
Hätkähdytti, noki kruunua
Ja nousi... ja samaan aikaan
Dadon putosi vaunuista -
Hän haukkoi henkeään kerran ja kuoli.
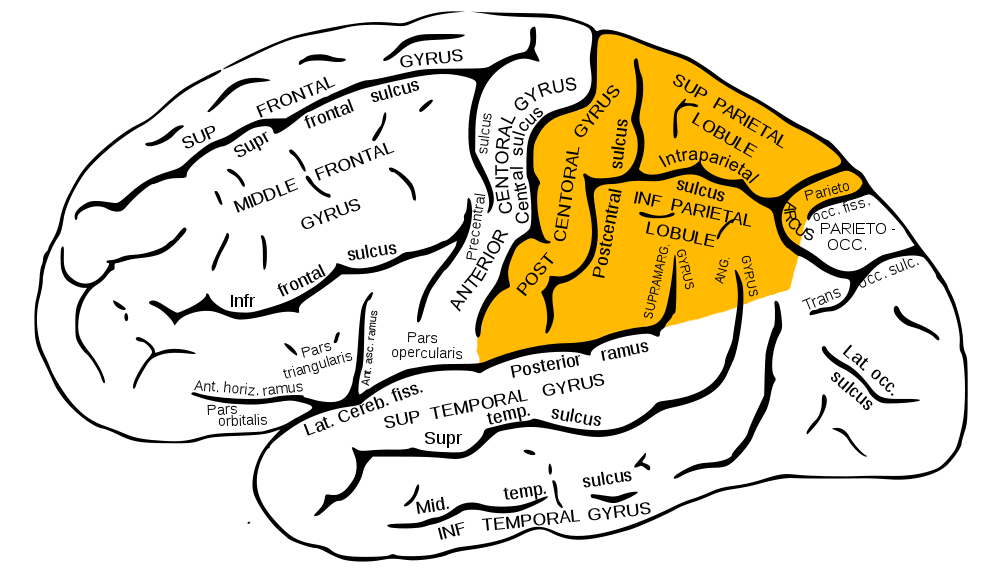
Näyttää siltä, että olet jo ymmärtänyt, mistä keskustellaan syklin "Kuinka aivot toimivat" seuraavassa materiaalissa. Olemme jo puhuneet otsalohkoista, temporaalisista ja parietaalisista, nyt siirrymme sivulohkoihin. Ne ovat myös latinaksi lääkäreille tuttuja Lobus parietalis.
Parietaalilohkot näkyvät keltaisella
Parietaalilohko sijaitsee juuri niskakyhmyn yläpuolella ja "koostuu" kolmesta kierteestä: yksi pystysuora- posterior keskus (vanhin osa) ja kaksi vaakasuuntaista - ylempi parietaalinen (uusi) ja alempi parietaalinen (uudempi).
Kuten otsalohkojen rakenne, ihmiskehon osat "projisoituvat" parietaalin keskeiseen etummaiseen gyrusin: alempi kolmannes on kasvot, keskikolmas on käsivarsi ja vartalo, ylempi on jalka. Älä unohda, että osuus on "kaksinkertaistunut", joten puolet siitä vastaa yhdestä (vastakkaisesta) puolesta kehosta.
Parietaalilohkojen rakenne
Lisäksi ylemmässä parietaalisessa gyrusssa on keskuksia, jotka vastaavat monimutkaiset tyypit syvä herkkyys: lihas-nivel-, kaksiulotteinen-tilaaisti (numeroiden, kirjainten, lyijykynällä piirrettyjen lukujen määrittely tai muu tylsä esine ihmisen iholla), painon ja liikkeen määrän tunne, esineiden tunnistamisen tunne koskettamalla.
Alemmassa parietaalilohkossa on harjoituskeskuksia, eli toiston ja harjoituksen aikana "automaattisiksi" tulleita liikkeitä, jotka kehittyvät oppimisprosessissa ja jatkuvassa harjoittelussa, esimerkiksi kävely, syöminen, pukeutuminen ja pian.
Parietaalilohko on osallisena somatosensoristen (lihasten, ihon, nivelten ja nivelten reseptoreista) sisäelimet) vapaaehtoisiin liikkeisiin vaikuttavat tiedot.
Ylemmän parietaalilohkon vaurioihin liittyy kyky tunnistaa esineitä koskettamalla niitä suljetuin silmin. Potilaat kuvaavat esineen yksilöllisiä ominaisuuksia, mutta eivät voi syntetisoida sen kuvaa.
Alemman parietaalilohkon tappiolla kehon järjestelmän tunne häiriintyy. Ihminen ei pysty ymmärtämään missä oikea ja missä vasen puoli on, hän ei tunnista omia sormiaan. Toinen häiriötyyppi on tietämättömyys puutteestaan (potilas väittää liikuttavansa halvaantuneita raajoja). Näille potilaille voi kehittyä pseudopolymelia.— ylimääräisten raajan tai kehon osien tunne. Tällaiset potilaat voivat itsenäisesti poistaa "häiritsevän" raajan tai myötävaikuttaa sen amputaatioon.
Kulmaisen gyrusen aivokuoren vaurioituessa potilas menettää ymmärrystä ympäröivän maailman avaruudellisen havainnoinnin, oman kehonsa asennon ja sen osien keskinäiset yhteydet. Tähän liittyy erilaisia psykopatologisia oireita: depersonalisaatio, derealisaatio. Niitä voidaan havaita tietoisuuden ja kriittisen ajattelun täydellisen säilymisen edellytyksenä.
Dominoivan pallonpuoliskon parietaalilohkon vaurioilla ihmiselle kehittyy dysleksia - kyvyttömyys lukea, erottaa oikea ja vasen sekä dyskalkulia - kyvyttömyys laskea. On huomionarvoista, että usein dyskalkulia on itsenäinen sairaus, eikä seuraus neurologisista tai psyykkisiä ongelmia. Näiden ongelmien lisäksi on mahdollista myös apraksia - rikkomus tai kyvyttömyys suorittaa jotakin tarkoituksenmukaista toimintaa (no esimerkiksi ottaa lasi ja juoda) säilyttäen samalla toiminnan peruskomponentit.
Anastasia Sheshukova
Takarautalohko sijaitsee puolipallojen takaosassa. Aivopuoliskon kuperalla pinnalla takaraivolohkossa ei ole teräviä rajoja, jotka erottaisivat sen parietaali- ja ohimolohkosta, lukuun ottamatta parietaali-okcipitaalisen uurteen yläosaa, joka sijaitsee puolipallon sisäpinnalla, erottaa parietaalilohkon takaraivolohkosta. Takarautalohkon ylemmän lateraalipinnan uurteet ja kierteet ovat epävakaita ja niillä on vaihteleva rakenne. Takalaukun lohkon sisäpinnalla on kannusura, joka erottaa kiilan (niskakyrskyn kolmiomaisen lohkon) linguaalista ja niskakyrskystä.
Oksipitaalilohkon toiminta liittyy visuaalisen tiedon havaitsemiseen ja käsittelyyn, monimutkaisten visuaalisen havainnon prosessien järjestämiseen. Tässä tapauksessa verkkokalvon ylempi puolisko heijastuu kiilan alueelle, joka havaitsee valon alemmista näkökentistä; lingulaarisen gyrusen alueella on verkkokalvon alapuoli, joka havaitsee valon ylemmistä näkökentistä.
Saari
Saaristo tai ns. suljettu lobule sijaitsee syvällä lateraalisessa urassa. Saaristo on erotettu vierekkäisistä vierekkäisistä osista pyöreällä uralla. Saaren pinta on jaettu pitkittäiskeskiurallaan etu- ja takaosaan. Makuanalysaattori projisoidaan saarekkeelle.
limbinen aivokuori
Aivopuoliskojen sisäpinnalla corpus callosumin yläpuolella on cingulaattinen gyrus. Tämä gyrus, jonka kannaksessa on corpus callosumin takana, siirtyy merihevosen lähellä olevaan gyrukseen - parahippokampaaliseen gyrukseen. Cingulate gyrus yhdessä paragitpocampal gyrus kanssa muodostavat holvikivyruksen.
Puolipallojen sisä- ja alapinnat yhdistyvät ns. limbiseen (marginaaliseen) aivokuoreen yhdessä amygdala kanssa aivokuoren ytimien ryhmästä, hajukanava ja sipuli sekä aivoaivojen etu-, temporaali- ja parietaalilohkot. aivokuoren sekä hypotuberous-alueen ja rungon retikulaarisen muodostumisen kanssa. Limbinen aivokuori yhdistetään yhdeksi toiminnalliseksi järjestelmäksi - limbinen-retikulaariseksi kompleksiksi. Näiden aivojen osien päätehtävä ei ole niinkään viestinnän tarjoaminen ulkomaailman kanssa, vaan aivokuoren sävyn, ajamien ja affektiivisen elämän säätely. Ne säätelevät monimutkaisia, monitahoisia sisäelinten toimintoja ja käyttäytymisvasteita. Limbi-retikulaarinen kompleksi on kehon tärkein integroiva järjestelmä. Limbinen järjestelmä on myös tärkeä motivaatioiden muodostuksessa. Motivaatio (tai sisäinen motivaatio) sisältää monimutkaisimmat vaistonvaraiset ja emotionaaliset reaktiot (ruoka, puolustus, seksuaalinen). Limbinen järjestelmä osallistuu myös unen ja valveillaolojen säätelyyn.
Limbinen aivokuori suorittaa myös tärkeän hajutoiminnon. Haju on ilmassa olevien kemikaalien havaitseminen. Hajuaivot henkilö tarjoaa hajuaistin sekä emotionaalisten ja käyttäytymisreaktioiden monimutkaisten muotojen järjestämisen. Hajuaivot ovat osa limbistä järjestelmää.
Hajuaivot koostuvat kahdesta osasta - perifeerisestä ja keskusosasta. Ääreisosaa edustavat hajuhermo, hajusipulit, primaariset hajukeskukset. Keskiosa sisältää merihevosen gyrus - hippokampuksen, hampaiden ja holvikierteen.
Hajureseptorilaitteisto sijaitsee nenän limakalvolla. Hermojohdejärjestelmän kautta tiedot reseptoreista välittyvät hajuanalysaattorin kortikaaliosaan (kuva 8).
Riisi. 8. Hajuanalysaattori (kaavio):
1 - hajuepiteeli, kaksisuuntaiset hajusolut; 2 - hajukäämissä; 3 - hajutie; 4 - ensisijaiset hajukeskukset; 5 - visuaalinen tuberkuloosi; 6 - kortikaalinen hajukeskus; 7 - corpus callosum
Hajuanalysaattorin aivokuoren osa sijaitsee singulate gyrus, merihevonen gyrus ja merihevosen koukku, jotka yhdessä muodostavat suljetun rengasmaisen alueen. Hajuanalysaattorin perifeerinen osa on yhdistetty molempien pallonpuoliskojen aivokuoren alueisiin.
Hajuanalysaattorin hajun havaitsemisen fysiologinen mekanismi ei ole täysin selvä. On olemassa kaksi päähypoteesia, jotka selittävät tämän prosessin luonnetta eri kohdista. Erään hypoteesin mukaan hajumolekyylien ja kemoreseptoreiden välinen vuorovaikutus tapahtuu avaimen ja lukon tavoin, ts. molekyylityyppi vastaa erityistä reseptoria. Toinen hypoteesi perustuu oletukseen, että hajuisen aineen molekyyleillä on tietty värähtelyaalto, johon hajureseptorit "viritetään". Molekyyleillä, joilla on samanlainen värähtely, tulisi olla yhteinen aalto ja vastaavasti antaa samanlaisia hajuja.
Termi "hajuaivot" suhteessa ihmisen fysiologiaan on jokseenkin mielivaltainen eikä paljasta täysin sen monitahoista ja yleismaailmallista toimintaa. Hajuaivojen keskeisen linkin "sijoittuminen" aivopuoliskoille ei ole sattumaa, ja se on seurausta siitä valtavasta "informaatiollisesta" roolista, joka hajuaistilla oli evoluutioprosessissa sopeutuessaan ulkoiseen ympäristöön ja säätelemällä komplekseja. käyttäytymisreaktiot. Ruoan saaminen, vastakkaista sukupuolta olevan yksilön valitseminen, jälkeläisistä huolehtiminen, alueen koskemattomuus, ryhmäyhteisöjen järjestäminen lajin sisällä - kaikki nämä päivittäiset toiminnot monissa eläimissä suoritetaan suoraan osallisena hienosti rakennettuun hajuvastaanottojärjestelmään ja Tämän perusteella useiden eläinten kyky lähettää hienovaraisia erilaistettuja erityisiä hajuisia aineita- Signaalien ilmoittajat.
Eläinten yleiset käyttäytymisreaktioiden muodot, jotka ilmenevät elinympäristön ja jälkeläisten päivittäisessä hoidossa, luovat vaikutelman, että heillä on järki. Näennäinen älykkyys on yksinkertaisesti seurausta reagoimisesta ulkoisiin ärsykkeisiin. Itse nämä ärsykkeet ja vastaukset niihin vastaavat kuitenkin täydellisesti eläinten biologisia tarpeita.
Ihmisten elämässä hajuaisti on menettänyt biologisen informaatioarvon, joka sillä oli eläimissä. Ihmisen hajujärjestelmä on suunniteltu suorittamaan sekä kapeaa, "omaa" toimintoa että eräänlaista tunteiden "latausta". Hajujen emotionaaliseen sfääriin kohdistuvan vaikutuksen voimakkuus, että ne ovat tärkein "tunteiden ravintoalusta", on tiedetty hyvin muinaisista ajoista lähtien ihmiskunnan historiassa.
Ihmisen hajuaisti voi vaihdella. Yleensä nämä vaihtelut ovat merkityksettömiä, mutta joissakin tapauksissa hajun terävyys voi olla erittäin korkea (hajuvesiteollisuuden maistajat).
Koska hajuanalysaattorilla on tärkeä rooli tunteiden säätelyssä, sen keskusosaan viitataan limbiseen järjestelmään, jota kutsutaan kuvaannollisesti "yhteiseksi nimittäjäksi" monille kehon tunne- ja visserosomaattisille reaktioille.
Suuret aivopuoliskot
ovat aivojen suurin osa. Ne peittävät pikkuaivot ja aivorungon. Aivopuoliskot muodostavat noin 78 % aivojen kokonaismassasta.
Organismin ontogeneettisen kehityksen prosessissa aivopuoliskot kehittyvät hermoputken terminaalisesta aivorakosta, joten tätä aivojen osaa kutsutaan myös telencephaloniksi.
Aivopuoliskot on jaettu keskiviiva syvä pystyhalkio oikealla ja vasen aivopuolisko. Keskiosan syvyydessä molemmat pallonpuoliskot on liitetty toisiinsa suurella tartunnalla - corpus callosumilla. Jokaisella pallonpuoliskolla on lohkoja: edestä, parietaalinen, ajallinen, takaraivo-.
Aivopuoliskon lohkot erotetaan toisistaan syvien uurteiden avulla. Kolme syvää uraa ovat tärkeimpiä: keskimmäinen (Rolandin), joka erottaa etulohkon päällimmäisestä; lateraalinen (Sylvian), joka erottaa ohimolohkon parietaalista, ja parietaali-okcipitaalinen, erottaen sivulohkon takaraivosta pallonpuoliskon sisäpinnalla.
Jokaisella pallonpuoliskolla on ylempi lateraalinen (kupera) - kupera, alempi - tyvipinta ja sisäinen - mediaalinen pinta. Jokaisessa pallonpuoliskossa on aivokierteitä, jotka on erotettu toisistaan uurteiden avulla. Ylhäältä puolipallo on peitetty kuorella - ohuella kerroksella harmaata ainetta, joka koostuu hermosolut.
Aivokuori- keskushermoston nuorin muodostelma evoluution kannalta. Ihmisillä se saavuttaa korkeimman kehityksensä. Aivokuorella on suuri merkitys kehon elintärkeän toiminnan säätelyssä, monimutkaisten käyttäytymismuotojen toteuttamisessa ja neuropsyykkisten toimintojen muodostumisessa.
Kuoren alla on valkea aine pallonpuoliskot, se koostuu hermosolujen prosesseista - johtimista. Aivomurtumien muodostumisen vuoksi aivokuoren kokonaispinta kasvaa merkittävästi. Puolipallon aivokuoren kokonaispinta-ala on 1200 cm2, jonka pinnasta 2/3 sijaitsee uurteiden syvyyksissä ja 1/3 puolipallojen näkyvällä pinnalla. Jokaisella aivojen lohkolla on erilainen toiminnallinen merkitys.
Aivokuori koostuu 4 lohkosta, jotka on erotettu toisistaan vaoilla. Pääurat, jotka erottavat etu-, parietaali- ja ohimolohkot, ovat Rolandov ja Sylviev.
Aivokuoren lohkot :
edestä (edestä ),
parietaalinen (parietaalinen ),
takaraivo (takaraivo- ),
ajallinen (ajallinen ),
Osakkeiden yhteydessä puhutaan aivojen paikallisista järjestelmistä.
AIVOJEN PAIKALLISET JÄRJESTELMÄT:
Aivojen takaraivoalueet suorittaa visuaalisen havainnon organisointitoiminnot. Takkaran aivokuoren ensisijaiset vyöhykkeet - perustoiminnot näkemys.
Toissijaiset osat takaraivokuoren - opto-gnostiset toiminnot.
Aivojen ajalliset alueet"vastuussa" kuulohavainnon järjestämisestä. Temporaalisen aivokuoren ensisijaiset vyöhykkeet ovat kuulon perustoiminnot.
Temporaalisen aivokuoren toissijaiset vyöhykkeet - akustis-gnostiset toiminnot.
Aivojen sensorimotoriset ja esimotoriset osat- liikkeiden järjestäminen. 1. Postcentral osat aivoissa - afferentti organisointi liikkeitä. 2. Premotoriset aivokuoren vyöhykkeet - liikkeen efferenttiorganisaatio (ohjelmointi).
3. Aivokuoren moottorivyöhykkeet - moottorianalysaattori, moottoriohjelman toteutus. Aivojen etulohkot (prefrontaaliset alueet) suorittaa henkisen toiminnan säätelyä, ts. toimintatilojen säätely, vapaaehtoiset liikkeet ja toimet, kognitiiviset prosessit sekä tunne- ja henkilösfääri ovat yleensä vastuussa ajattelusta, älyllisestä toiminnasta. Aivojen parietaaliset alueet Ne edustavat ensisijaiset vyöhykkeet (kaiken tyypit iho-kinesteettinen herkkyys), toissijaiset vyöhykkeet (visuaalis-spatiaaliset esitykset, ideat kehosta, somatognoosista, stereognoosista) ja tertiäärisiä vyöhykkeitä.
etulohko miehittää pallonpuoliskojen etuosat. Sen erottaa parietaalilohkosta keskussulkus ja ohimolohkosta lateraalisulkus. Etulohkossa on neljä gyriä: yksi pystysuora - precentraalinen ja kolme vaakasuoraa - ylä-, keski- ja alaosa.
Kierteet on erotettu toisistaan vaoilla. Etulohkojen alapinnalla erotetaan suora ja orbitaalinen gyrus. Suora gyrus sijaitsee pallonpuoliskon sisäreunan, hajuuran ja puolipallon ulkoreunan välissä.
Hajuvaon syvyyksissä sijaitsevat hajutulppa ja hajutie. Henkilön etulohko muodostaa 25-28% aivokuoresta, otsalohkon keskimääräinen massa on 450 g.
Otsalohkojen toiminta liittyy vapaaehtoisten liikkeiden järjestämiseen, puheen motorisiin mekanismeihin, monimutkaisten käyttäytymismuotojen säätelyyn ja ajatteluprosesseihin. Useita toiminnallisesti tärkeitä keskuksia on keskittynyt otsalohkon konvoluutioihin. Keskimmäinen etummainen gyrus on ensisijaisen motorisen alueen "esitys", jossa on tiukasti määritelty kehon osien projektio. Kasvot "sijaitsevat" gyrusen alemmassa kolmanneksessa, käsi keskimmäisessä kolmanneksessa, jalka sisään ylempi kolmannes. Runko on edustettuna ylemmän frontaalisen gyrusin takaosissa. Siten henkilö projisoituu ylösalaisin ja alaspäin keskikyrukseen.
Keskimmäinen etummainen gyrus Yhdessä etupyörän vierekkäisten takaosien kanssa sillä on toiminnallisesti erittäin tärkeä rooli. Se on vapaaehtoisten liikkeiden keskus. Keskisen gyrusen aivokuoren syvyyksissä ns pyramidisolut -keskusmotorinen neuroni- päämoottoritie alkaa - pyramidaalinen tai kortikospinaalinen polku. Motoristen neuronien perifeeriset prosessit nousevat aivokuoresta, kerääntyvät yhdeksi voimakkaaksi nipuksi, kulkevat pallonpuoliskojen keskeisen valkoisen aineen läpi ja tulevat aivorunkoon sisäisen kapselin kautta; aivorungon päässä ne ylittävät osittain (kulkevat puolelta toiselle) ja laskeutuvat sitten selkäydin. Nämä oksat päättyvät harmaa aine selkäydin. Siellä ne joutuvat kosketuksiin perifeerisen motorisen neuronin kanssa ja välittävät impulsseja sille keskusmotorisesta neuronista. Vapaaehtoisen liikkeen impulssit välittyvät pyramidin polkua pitkin.
Yläkerran takaosassa on myös aivokuoren ekstrapyramidaalinen keskus, joka on anatomisesti ja toiminnallisesti tiiviisti yhteydessä ns. Ekstrapyramidaalinen järjestelmä- moottorijärjestelmä, joka auttaa mielivaltaisen liikkeen toteuttamisessa. Tämä on järjestelmä mielivaltaisten liikkeiden "tarjoamiseksi". Koska ihmisen ekstrapyramidaalinen järjestelmä on fylogeneettisesti vanhempi kuin pyramidaalinen järjestelmä, se tarjoaa "oppittujen" motoristen toimintojen automaattisen säätelyn, yleisen lihasjänteen ylläpidon, perifeerisen motorisen laitteen "valmiuden" liikkumiseen, lihasjänteen uudelleen jakautumisen liikkeiden aikana. Lisäksi se osallistuu normaalin asennon ylläpitämiseen.
Keskimmäisen frontaalisen gyrusen takaosassa on etuosan silmämotorinen keskus, joka ohjaa pään ja silmien ystävällistä, samanaikaista pyörimistä (pään ja silmien pyörimiskeskus vastakkaiseen suuntaan). Tämän keskuksen ärsytys saa pään ja silmät kääntymään vastakkaiseen suuntaan. Tämän keskuksen toiminnalla on suuri merkitys niin kutsuttujen suuntautumisrefleksien toteuttamisessa, jotka ovat erittäin tärkeitä eläinten elämän säilymisen kannalta.
Takaosassa alemman frontaalisen gyrus on motorinen puhekeskus(Brockin keskusta).
Myös aivopuoliskojen etukuori vastaanottaa Aktiivinen osallistuminen ajattelun muodostuksessa, määrätietoisen toiminnan organisoinnissa, pitkän tähtäimen suunnittelussa.
ajallinen lohko miehittää pallonpuoliskojen alemman sivupinnan. Ohimolohko on erotettu etu- ja parietaalilohkosta lateraalisella uralla.
Ohimolohkon ylemmällä sivupinnalla on kolme kiertettä - ylempi, keskiverto Ja alempi. Ylämäinen temporaalinen kypärä sijaitsee ylemmän ja ylemmän ohimoraon välissä, keskimmäinen kypärä on ylemmän ja alemman ohimohaavan välissä ja inferior gyrus on alemman ohimoraon ja poikittaisen aivohalkeaman välissä. Ohimolohkon alapinnalla erotetaan alempi temporaalinen gyrus, lateraalinen occipitotemporaalinen gyrus, hippokampaalinen gyrus (merihevosen jalat).
Temporaalilohkon toiminta liittyvät kuulo-, maku-, hajuaistimuksiin, puheäänien analysointiin ja synteesiin, muistimekanismeihin. Ohimolohkon ylemmän lateraalipinnan päätoiminnallinen keskus sijaitsee ylemmässä ohimokivussa. Tässä on kuulo eli gnostinen puheen keskus (Wernicken keskus).
Ylämäisessä temporaalisessa gyrusssa ja ohimolohkon sisäpinnalla on aivokuoren kuuloprojektioalue. Hajuprojektioalue sijaitsee hippokampuksen gyrus-alueella, erityisesti sen etuosassa (ns. koukku). Hajuprojektioalueet ovat myös makuisia.
Ohimolohkoilla on tärkeä rooli kompleksin järjestämisessä henkisiä prosesseja, erityisesti muisti.
parietaalinen lohko miehittää pallonpuoliskon ylemmän sivupinnan. Frontaalisesta parietaalilohkosta, edestä ja sivulta, sitä rajoittaa keskisulkus, temporaalisesta alhaalta - lateraalisesta uurresta, takaraivosta - kuvitteellinen viiva, joka kulkee parietaali-okcipitaalisen uurteen yläreunasta puolipallon alareuna.
Parietaalilohkon ylemmällä lateraalipinnalla on kolme kiertettä: yksi pystysuora - posterior keskus ja kaksi vaakasuoraa - ylempi parietaalinen ja alempi parietaalinen. Alemman parietaalisen gyrusen osaa, joka ympäröi lateraalisen sulcusin takaosan, kutsutaan supramarginaaliksi (supramarginaaliksi) ja yläpäätä ympäröivää osaa. ajallinen gyrus, solmu (kulma)alue.
Parietaalilohko, kuten otsalohko, muodostaa merkittävän osan aivopuoliskot. Fylogeneettisin termein siinä erotetaan vanha osa - takamyrsky, uusi - ylempi parietaalinen gyrus ja uudempi - alempi parietaalinen gyrus. Parietaalilohkon toiminta liittyy herkkien ärsykkeiden havaitsemiseen ja analysointiin, avaruudelliseen suuntautumiseen. Parietaalilohkon kierteisiin on keskittynyt useita toiminnallisia keskuksia.
Takaosassa keskikehän herkkyyskeskukset projisoidaan kehon projektiolla, joka on samankaltainen kuin anteriorisessa keskigyrusessa. Gyrusen alemmassa kolmanneksessa kasvot projisoidaan, keskikolmanneksessa - käsivarsi, vartalo, ylemmässä kolmanneksessa - jalka. Ylimmässä parietaalisessa gyrusssa on keskuksia, jotka vastaavat monimutkaisista syvien herkkyyden tyypeistä: lihas-nivel-, kaksiulotteinen-avaruustuntemus, painon- ja liiketilavuuden tunne, esineiden tunnistamisen tunne koskettamalla.
Siten herkän analysaattorin kortikaalinen osa sijoittuu parietaalilohkoon.
Praxiskeskukset sijaitsevat alemmassa parietaalilohkossa. Praxis ymmärretään määrätietoisina, toisto- ja harjoitusprosessissa automatisoituneina liikkeinä, jotka kehittyvät oppimisprosessissa ja jatkuvassa harjoittelussa yksilön elämän aikana.
Kävely, syöminen, pukeutuminen, mekaaninen kirjoittaminen, erilaisia työtoiminta (esim. kuljettajan liikkeet autoa ajaessaan, niitto jne.) ovat käytäntöjä.
Praxis- ihmisen motorisen toiminnan korkein ilmentymä. Se toteutetaan eri alueiden yhdistetyn toiminnan tuloksena. aivokuori.
Okcipital lohko miehittää pallonpuoliskojen takaosat. Aivopuoliskon kuperalla pinnalla takaraivolohkossa ei ole teräviä rajoja, jotka erottaisivat sen parietaali- ja ohimolohkosta, lukuun ottamatta parietaali-okcipitaalisen uurteen yläosaa, joka sijaitsee puolipallon sisäpinnalla, erottaa parietaalilohkon takaraivolohkosta.
Takarautalohkon ylemmän lateraalipinnan uurteet ja kierteet ovat epävakaita ja niillä on vaihteleva rakenne.
Takalaukun lohkon sisäpinnalla on kannusura, joka erottaa kiilan (niskakyrskyn kolmiomaisen lohkon) linguaalista ja niskakyrskystä.
Oksipitaalilohkon toiminta liittyy havainnointiin ja käsittelyyn visuaalista tietoa, visuaalisen havainnon monimutkaisten prosessien organisointi. Tässä tapauksessa verkkokalvon ylempi puolisko heijastuu kiilan alueelle, joka havaitsee valon alemmista näkökentistä; lingulaarisen gyrusen alueella on verkkokalvon alapuoli, joka havaitsee valon ylemmistä näkökentistä.
Saari, tai ns suljettu lobule, sijaitsee sivuvaon syvyydessä. Saaristo on erotettu vierekkäisistä vierekkäisistä osista pyöreällä uralla.
Saaren pinta on jaettu pitkittäiskeskiurallaan etu- ja takaosaan. Saarella on ennustettu makuanalysaattori.
Hajuanalysaattori
Hermosolut, jotka havaitsevat hajuärsytystä, sijaitsevat nenäontelon yläosien limakalvolla. Sieltä näiden solujen aksonit menevät kallononteloon ja menevät hajusoluihin. Niistä lähetetään hermosäikeitä ohimolohko (sisäpinta) missä ovat hermosolut hajuanalysaattori.
Maku-analysaattori
Tämä analysaattori alkaa kielen makuhermojen hermopäätteistä, joita niissä edustavat makuhermot. Makuhermoista lähtevät hermosäidut menevät aivoihin ja päättyvät hajuanalysaattorin hermien tapaan klo. sisäpinta ajallinen lohko.
corpus callosum- kaareva ohut levy, fylogeneettisesti nuori, yhdistää mediaanipinnat molemmat pallonpuoliskot. Corpus callosumin pitkänomainen keskiosa siirtyy takaa paksuuntumaan ja edestä kaareva ja kaareva alaspäin.
Corpus callosum yhdistää aivopuoliskojen fylogeneettisesti nuorimmat osat ja sillä on tärkeä rooli niiden välisessä tiedonvaihdossa.
Aivorunko TAI AIVOKANNA -
perinteisesti erottuva aivoalueiden järjestelmä, joka on selkäydintä jatkava laajennettu muodostelma.
Aivorunkoon kuuluvat aina ydin, poni ja väliaivot. Usein se sisältää pikkuaivot, joskus välilihakset.
Ydin -
aivojen osasto. Siellä on myös perinteinen nimi bulbus (sipuli, tämän osaston muodosta).
Medulla oblongata tulee aivorunkoon.
Ulkopuolella, vatsan (kasvojen) puolella, on pyramidit (ne sisältävät kortikospinaalikanavan - polun aivokuoresta selkäytimen motorisiin neuroniin) ja oliiveja (ne sisältävät alemman oliivin ytimiä, jotka liittyvät tasapainon ylläpitämiseen) . Selkäpuolella: ohuet ja kiilanmuotoiset niput, jotka päättyvät ohuiden ja kiilanmuotoisten ytimien tuberkoihin (vaihda kehon ala- ja yläpuoliskon syväherkkyystiedot), rombisen kuopan alapuoli, joka on neljännen kammion pohja ja sitä erottavat köysikappaleet tai pikkuaivojen alaraajat.
Sisällä ovat myös kallohermojen ytimet VIII - XII (ja yksi ytimistä VII), osa verkkokalvon muodostusta, mediaalista silmukkaa ja muita nousevia ja laskevia polkuja.
Se on katkaistun kartion muotoinen.
Tällaisten tutkijoiden, kuten R. Magnuksen ja I. F. Kleinin, tutkimuksen ansiosta todettiin, että ytimessä on monimutkainen refleksikeskusten järjestelmä, joka tarjoaa tietyn aseman kehossa staattisten ja staattis-kineettisten refleksien vuoksi. Nämä refleksit ovat itse asiassa mekanismeja lihasten sävyn uudelleenjakaumiseksi siten, että eläimelle säilyy mukava asento (asentotoniset refleksit) tai palautuu epämukavalta (korjaavat refleksit) ja tasapaino säilyy myös kiihdytyksen aikana (stato-kineettiset refleksit). Näiden refleksien toteutus tapahtuu sellaisten runkomuodostelmien, kuten retikulaarimuodostelman, punaisen tuman ja vestibulaarisen ytimien, osallistuessa.
Retikulaarinen muodostuminen - se on muodostelma, joka kulkee selkäytimestä talamukseen rostral (aivokuoren) suunnassa. Sensorisen tiedon käsittelyyn osallistumisen lisäksi retikulaarinen muodostus aktivoi aivokuorta ja säätelee siten selkäytimen toimintaa. Ensimmäistä kertaa verkkokalvon muodostumisen vaikutusmekanismi lihasten sävy perusti R. Granit: hän osoitti, että retikulaarinen muodostus pystyy muuttamaan γ-motoristen hermosolujen toimintaa, minkä seurauksena ihaksonit (γ-efferentit) aiheuttavat lihaskarojen supistumista ja sen seurauksena lisääntymistä lihasreseptoreiden afferenteissa impulsseissa. Nämä impulssit, jotka tulevat selkäytimeen, aiheuttavat α-motoristen hermosolujen virittymisen, mikä on syy lihasjänteyteen.
On osoitettu, että kaksi hermosolujen klusteria osallistuu tämän verkkokalvomuodostelman toiminnon suorittamiseen: sillan retikulaarimuodostelman hermosolut ja pitkittäisytimen retikulaarimuodostelman neuronit. Retikulaarimuodostelman hermosolujen käyttäytyminen ydinjatke samanlainen kuin hermosolujen käyttäytyminen sillan retikulaarisessa muodostelmassa: ne aiheuttavat flexor-lihasten α-motoristen hermosolujen aktivaatiota ja siten estävät ojentajalihasten α-motoristen neuronien toimintaa. Sillan retikulaarisen muodostelman neuronit toimivat täsmälleen päinvastoin, kiihottavat ojentajalihasten α-motorisia hermosoluja ja estävät taivutuslihasten α-motoristen neuronien toimintaa. Retikulaarimuodostelmalla on yhteys pikkuaivoon (osa sen tiedosta menee ytimeen (korkki- ja pallomaisen pikkuaivojen ytimistä) ja teltasta sillan hermosoluihin) ja aivokuori, josta se saa tietoa. Tämä viittaa siihen, että retikulaarinen muodostus on epäspesifisen sensorisen virtauksen kerääjä, joka mahdollisesti osallistuu lihastoiminnan säätelyyn.
tärkeä toiminnallinen merkitys verkkomainen tai aivorungon verkostomuodostusta, joka kehittyy vagus-, vestibulaari- ja kolmoishermot.
Verkkomuodostelma koostuu erikokoisista ja -muotoisista hermosoluista sekä tiheästä verkostosta hermosäikeitä, joka kulkee eri suuntiin ja sijaitsee pääasiassa lähellä kammiojärjestelmää. Retikulaariselle muodostukselle annetaan ensisijainen merkitys aivokuoren ja aivokuoren välisissä suhteissa. Se sijaitsee keskikerroksissa. ydinjatke,hypotalamus, tegmentum harmaa aine, pons.
Lukuisat sivut kaikista afferenteista (sensorisista) järjestelmistä lähestyvät retikulaarista muodostumista. Näiden sivuelementtien kautta myös periferiasta tuleva ärsytys, joka suuntautuu tietyille aivokuoren alueille hermoston tiettyjä reittejä pitkin, saavuttaa myös retikulaarimuodostelman. Epäspesifiset nousevat järjestelmät (eli reitit retikulaarisesta muodostumisesta) saavat aikaan aivokuoren virittymisen ja sen toiminnan aktivoinnin.
Nousevien epäspesifisten järjestelmien ohella aivorungon läpi kulkevat laskevat epäspesifiset järjestelmät, jotka vaikuttavat selkärangan refleksimekanismiin.
Retikulaarinen muodostus liittyy läheisesti limbiseen järjestelmään sekä aivokuoreen. Tästä johtuen keskushermoston korkeampien osien ja vartalon välille muodostuu toiminnallinen yhteys. aivot. Tätä järjestelmää kutsutaan limbi-retikulaariseksi kompleksiksi tai limbi-retikulaariseksi akseliksi. Tämä monimutkainen rakenteellinen ja toiminnallinen kompleksi varmistaa tärkeimpien toimintojen yhdistämisen, joiden toteuttamiseen aivojen eri osat osallistuvat.
Tiedetään, että aivokuoren valvetila saadaan aikaan spesifisillä ja epäspesifisillä järjestelmillä. Aktivointireaktiota ylläpitää jatkuva impulssien syöttö reseptoreista kuulo, visuaalinen, haju, maku ja ihon kinesteettiset analysaattorit. Nämä ärsykkeet välittyvät tiettyjä afferentteja pitkin aivokuoren eri osiin. Kaikilta osallistujilta talamus, ja sitten lukuisat retikulaarisen muodostelman sivut lähtevät afferenttien aivopuoliskojen aivokuoreen, mikä varmistaa sen nousevan aktivoivan toiminnan.
Retikulaarinen muodostus puolestaan vastaanottaa impulsseja pikkuaivot, subkortikaaliset ytimet, limbinen järjestelmä jotka tarjoavat emotionaalisesti mukautuvia käyttäytymisvasteita, motivoivia käyttäytymismuotoja. Kuitenkin aste, jolla ei-spesifinen järjestelmä tuottaa mukautuvat ehdottomat refleksireaktiot ihmisillä ja eläimillä, on erilainen. Jos eläimissä aivokuoren alaisilla muodostelmilla ja limbisellä järjestelmällä on johtava rooli organismin elintärkeiden tarpeiden täyttämisessä sen selviytymiselle ympäristöön, niin ihmisellä aivokuoren dominanssin vuoksi aivojen syvien rakenteiden (subkortikaaliset muodostelmat, limbinen järjestelmä, retikulaarinen muodostus) toiminta on enemmän kuin eläimellä alisteinen aivokuori. Retikulaarisella muodostuksella on tärkeä rooli lihasjänteen säätelyssä. Lihasten sävyä säätelevät kahden tyyppiset retikulospinaaliset kanavat. Nopeasti johtava retikulospinaalitie säätelee nopeita liikkeitä; hitaasti johtava retikulospinaalinen reitti - hitaat tonic liikkeet.
Medulla oblongatan retikulaarinen muodostuminen liittyy decerebrate-jäykkyyden esiintymiseen. Kun aivorunko leikataan pitkittäisytimen yläpuolelle, selkäytimen motorisiin hermosoluihin inhiboivan vaikutuksen omaavien neuronien aktiivisuus laskee, mikä johtaa luurankolihasten jyrkkään kohoamiseen.
Medulla oblongatan toiminnot
Suojarefleksit (esim. yskiminen, aivastelu).
Tärkeät refleksit (esim. hengitys).
verisuonten sävyn säätely.
Medulla oblongatan refleksikeskukset:
suojaava (yskiminen, aivastelu jne.)
äänensäätökeskukset luurankolihas ylläpitämään henkilön asentoa.
selkärangan refleksiajan lyhentäminen tai pidentäminen
ruoansulatus
sydämen toimintaa
Pons
Pons Varolii (Constanzo Varolian puolesta) tai silta - aivojen osa, on yhdessä pikkuaivojen kanssa osa takaaivoja. kuuluu aivoihin,
Sillan pohjassa on laskevia polkuja: kortikospinaalinen pyramiditie, kortikobulbaari, kortikobridge-tie.
keskiaivot(lat. Mesencephalon) - osa aivoja, muinainen visuaalinen keskus. Sisältyy aivorunkoon.
keskiaivojen toiminnot
1. moottori,
2. sensorinen (näkö, kuulo),
3. pureskelun ja nielemisen säätely,

