5-10-2012, 19:14
Функции на кората
челен дял. Основната функция на фронталните дялове е контролът на произволните движения, координацията на двигателните механизми на експресивната реч, както и осигуряването на "творческо" или абстрактно мислене.
Фронталния лоб може да бъде разделен на пет области:
- мотор,
- премотор,
- фронтално моторно зрение,
- Моторната област на речта на Broca
- и фронтална асоциативна зона
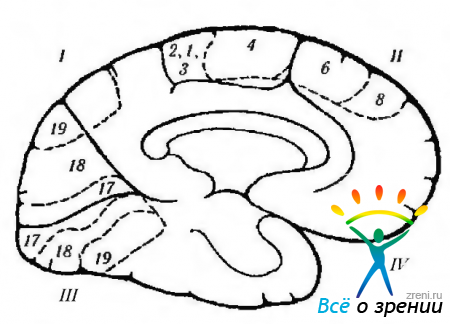
Ориз. 4.1.12.Схематично представяне на функциите на кората голям мозък, локализирани на медиалната повърхност, и нарушаването им по време на различни заболявания. придружено от възбуждане или разрушаване на кората (според Crosby et al., 1962): 1 - париетален лоб. Централна извивка (4). Наляво тилната област: -алексия без аграфия. Заден кортекс: -затруднено превеждане на погледа; - Синдром на Балинт (двустранен), 11 - челен лоб. Допълнително двигателно поле (6): + повдигане на противоположната ръка, обръщане на очите и главата към ръката; + свиване на зеницата; + забавяне на речта. Фронтално моторно очно поле (8). Колан gyrus: + разширяване на зеницата; + лакримация. орбитална повърхност. III - тилен лоб. първичен зрителен кортекс. Контралатерално зрение: - дясностранна хемианопия. IV-темпорален лоб: парализа на движенията на очите; ипсилатерална хемиплегия; хидроцефалия; децеребрална ригидност; педункулярна халюциноза; контралатерална хемианопсия; синдром на Парино. Под повърхността: - разпознаване на лица. Забележка. Цитоархитектоничните полета съответстват на класификацията на Бродман. Плюс показва резултата от електрическа стимулация на кората, а минус - резултата от разрушаването на полето
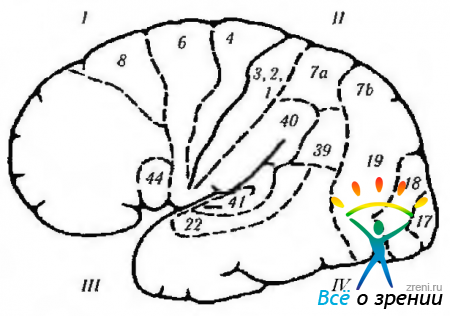
Ориз. 4.1.13.Страничната повърхност на мозъка (обозначенията са подобни на тези, дадени на предишната фигура): 1 - челен лоб. Моторно поле (4). Премоторно поле (6). Моторно очно поле (8): + контралатерална сакада; + затваряне на противоположното око; + гомолатерално приятелско отклонение на окото. Моторно речево поле (44): - двигателна (моторна) афазия. 11 - париетален лоб: - (- проста халюциноза; - илюзии; - метаморфопсия: - загуба на внимание; - агнозия; - апраксия; - отслабване на оптокинетичния нистагъм; - хемианопия в долния квадрант. Супрамаргинален гирус (40): - амнезия; - жаргон афазия Angular gyrus (39): - аграфия; - акалкулия; - загуба на способност да се прави разлика между ляво и дясно правилната страна; агнозия на пръстите. соматична чувствителност. III-темпорален лоб: + зрителни халюцинации; + дежавю; + нарушено обоняние; + гърчове; + автоматизми; - горна квадрианопсия; -загуба на паметта; - Синдром на Klüver-Bucy. първичен слухов кортекс. Вторична слухова кора: -слухова афазия. Planum temporale: - дислексия. IV-окципитален лоб (17, 18, 19): + усещане за светлина; -(-усещане за цветове; -(-усещане за движение; + усещане за линейни образи. Визуални асоциативни полета (18, 19): -(-контралатерални движения на очите като плавно преследване; + вергентни движения; -синдром на Антон
Първична двигателна зона(моторна зона; 4-то поле по Бродман).
двигателен регион b е пред централната бразда, която разделя фронталния лоб от теменния лоб. Състои се от голям брой гигантски пирамидални клетки на Бетц.
Представлява движения, а не отделни мускули. Клетките, които контролират движенията, са разположени по такъв начин, че движенията на долната половина на тялото са представени в горната част на гируса, а горната половина в долната. Невронните аксони се спускат през вътрешната капсула и педункулите на мозъка под формата на кортикобулбарни и кортикоспинални пътища.
Отстраняване на моторната кора води до парализа.
Премоторна зона(поле b) лежи директно пред основното двигателен кортекс, заемащ задната част на трите хоризонтално ориентирани фронтални извивки.
Той образува влакната на фронтопонтоцеребеларните пътища и също така дава множество влакна на базалните ганглии.
Предполага се, че премоторният кортекс участва в планирането на двигателните действия. Това се доказва от факта, че само когато се мисли за планираното действие, невроните на шестото поле се възбуждат. Стимулирането на тези неврони води до повдигане на ръцете обратната страна, а главата и очите се обръщат по посока на повдигнатия крайник. Зениците се разширяват и говорът е нарушен (вижте фиг. 4.1.12, 4.1.13).
Унищожаването на тази област е придружено псевдомозъчна атаксия на противоположната страна. Способността за копиране на заучени движения (апраксия) също е нарушена. В този случай отделните движения не са повредени, но пациентът не е в състояние да ги произведе в желаната последователност.
Фронтална моторна зрителна зона(поле 8 по Бродман).
Тази област (поле 8 и частично 6 и 9) заема задната част на средната фронтална извивка (gyrus frontalis medius).
Фронтална моторна зрителна зона важни за произволните движения на очите. Някои области определят конвергенция, дивергенция и акомодация.
Трябва да се отбележи, че движението на очите, зеницата и клепачите могат да бъдат открити по време на електрическа стимулация на почти всички области на мозъчните полукълба. Електрическата стимулация на областта на предното моторно око води до сакадични движения, за разлика от реакцията на стимулация на полета 18, 19 и 22 (плавно проследяване и вергентни движения). Стимулирането на 8-мо поле при маймуни е придружено от съпътстващо контралатерално отклонение на очите. В този случай зеницата се разширява, а главата и торсът се обръщат в обратна посока. Увреждането на фронталния лоб причинява обратима загуба на способността за контралатерални сакадични движения.
Тази област на кората се отличава с множество връзки.
Аферентни влакнанавлизат в зрителния фронтален кортекс от зрителния туберкул, както и от горния темпорален, перистриален и париетален (област 7) кортекс. Тук са открити неврони, които контролират движенията на очите.
Еферентипроектирани към базалните ганглии, таламуса, претекталната област (включително ядрото на оптичния тракт), горните коликули на квадригемината и част ретикуларна формациямост.
На свой ред трябва да се отбележи, че фронталните зрителни области се проектират върху контралатералните фронтални области и върху кортикалните области от същата страна. Те осигуряват визуално възприятие (на 7-мо поле париетална кораи перистриална кора).
Когато се проектира върху среден мозъквлакната на префронталната кора пресичат предната част на вътрешната капсула и се разделят на дорзален и вентрален път в ростралния диенцефалон.
Дорзалният трансталамичен път пресича дорзомедиалното и интраламиларното ядро на таламуса оптикус и средната част на възглавницата, образувайки в същото време не голям бройсинаптични връзки. Синапси се намират също в претекталните ядра и в горните туберкули на квадригемината.
Вентрален педункуло-тегментален пътпреминава в най-вентралната част на мозъчния ствол, доставяйки влакна към ядрата на хипоталамуса и по-дълбоко - към горните туберкули на квадригемината. В същото време фронталните зрителни области имат топографска проекция върху горните туберкули на квадригемината. Пътеката завършва в ретикуларната формация на моста.
Трети префронтален лигаментвъзниква в областта на диенцефално-мезенцефалния възел, разположен близо до червеното ядро. Този път се проектира върху ядрото на медиалния надлъжен фасцикулус и междинното ядро на Cajal от същата страна, както и върху ядрото на Darkshevich и растралната част на окуломоторното ядро от двете страни.
Различни области на фронталния зрителен кортекс контролират сакадичните движения на очите. различна амплитуда, както и посоката на сакадите. Те осигуряват така нареченото „визуално внимание“, насочвайки погледа към „целта“.
В момента се разследва ролята на фронталния лоб във функционирането на клепачите. Доброволното затваряне на клепачите изисква отпускане на повдигащия клепач и свиване на орбикуларния очен мускул. Има противоречия относно локализацията на областите на мозъка, които контролират волевите движения на клепача. Тези места могат да бъдат разположени в двигателната премоторна или фронтална моторна зрителна област. Стимулирането на тази зона води до затваряне на контралатералния клепач. Едностранното увреждане на недоминиращите фронтални лобове може да бъде придружено от двустранно неуспешно затваряне на клепачите.
Моторна речева зона на Broca(полета 44 и 45 по Бродман).
Тази зона се намира в задната-долна част на фронталния лоб. Тя допринася за инициирането на речта. Увреждането на зоната на Broca води до афазия, която се състои в трудността на свързването в определена последователност от думи, за да се осигури свързана реч (експресивна афазия). Пациентите разбират както писмен, така и говорим език, но обикновено не могат да произнесат нищо. Способността за писане също е нарушена, въпреки че ръката може да се използва за други дейности.
фронтална асоциативна зона. Тази важна област в предната половина на фронталния лоб е отговорна за инициирането на всички видове поведенчески-интелектуални, висцерални и емоционални функции. Промените в паметта, интелекта и личността настъпват при дразнене или масивно разрушаване на фронталните лобове. Това е особено силно изразено в тези случаи. когато е засегнато доминантното полукълбо.
Обонятелен трактсе намира под орбиталната повърхност на фронталния лоб и е единственият сензорен път, който достига до кората без образуване на синапси в таламуса. Едностранен тумор на орбиталната повърхност на фронталния лоб (менингиом на обонятелния сулкус или менингиом на малкото крило) сфеноидна кост) може да доведе до компресия на обонятелния тракт и оптичен нерв. В резултат на това се развива аносмия (липса на обоняние), ипсилатерална атрофия на зрителния нерв и подуване на главата на зрителния нерв от противоположната страна (синдром на Кенеди Фостър).
Увреждането на структурите на лимбичната система във фронталния дял, особено на зъбния гирус, може да причини смущения в емоционалната страна на живота, както и висцерални признаци, включително безпричинно сълзене и промени в реакцията на зеницата към светлинни стимули.
париетален лоб(вижте фиг. 4.1.12, 4.1.13). Като цяло функцията на теменните дялове е свързана с чувствителността, паметта, свързана с речта и ученето, както и с ориентацията на тялото в пространството чрез обработка на визуална информация.
В предния париетален лоб има извивка, разположена успоредно на прецентралната извивка или моторна кора. Това е постцентралната извивка или първичната соматосензорна кора (полета 3, 1 и 2) (виж Фиг. 4.1.13). Останалата част от париеталния лоб може да бъде разделена на два лобула - горен и долен. Полета 5 и 7, разположени на повърхността на горната париетална лобула, са свързани с корелацията на соматосензорната информация, което позволява съзнателно да се оцени теглото, повърхностните свойства, размера и формата на обекта.
Долен париетален лобул(супрамаргинална и ъглова извивка) е тясно свързана с механизмите на речта, тъй като разрушаването на тази област на кората в лявото полукълбо води до нарушено възприятие на речта.
Ролята на париеталния лоб в пространствената ориентация все още не е проучена. Въпреки това, при пациенти с неговото поражение се отбелязва нарушение на тази функция.
Аферентните влакна преминават през париеталния лоб, свързвайки области 18 и 19 с фронталните дялове и мозъчния ствол. Тези пътища включват горния и долния надлъжни снопове.
Предполага се, че париеталната кора (поле 7) осигурява функцията "внимание" към визуалните цели.
Седмото поле получава аференти от cingulate gyrus(g. cinguli), от претекталната област и горните туберкули на квадригемината през ядрата на таламуса. Също така се откриват аференти, преминаващи от фронталната зрителна област и престриатния кортекс към кората на париеталния лобул.
Еферентите се проектират върху претекталната област, горните туберкули на квадригемината, сивото вещество, разположено около силвиевия акведукт, и върху париеталните дялове на двете полукълба на мозъка.
Когато очите се движат, невроните на долния париетален лоб се активират, което показва тяхната роля в контролирането на произволните движения на очите. Дейността продължава докато трае наблюдението на целта.
Клетките на 7-мо поле не определят сакадичните движения, тъй като тяхната активност не се увеличава по време на сакади.
Стимулиране на ъгловия гирус(g. angularis) води при опитни животни до приятелско отклонение на очите. Това са полета 38 и 39, минаващи около задния ръб на средната темпорална извивка (g. temporalis medius). Оперативно отстраняванетази област на кората не води до нарушено движение на очите. Двустранното увреждане на париеталната кора може да доведе до развитие на моторна апраксия на окото.
Поражението на париеталния лоб също е придружено от нарушение на визуалните двигателни механизми, които осигуряват оптокинетичен нистагъм, по време на който очите произвеждат сакади за търсене и фиксиране на целта.
Други двигателни признаци на изолирани лезии на париеталния лоб включват приятелско отклонение на очите в обратна посока от мястото на нараняванес принудително затваряне на клепачите, ниска подвижност на пациента и лош контрол (неразбиране) на крайниците от противоположната страна. Трудностите при разбирането на естеството на действието на крайниците водят до конституционална и окуломоторна апраксия. Пациенти с увреждане на париеталния лоб може да имат затруднения при преместването на погледа си от един визуален обект към друг в рамките на контралатералната половина на зрителното поле.
Друг признак на дисфункция на париеталния лоб е невъзможност за фиксиране на визуален обект.
В допълнение към сензорните функции, които са локализирани в задната устна на централната бразда на Роландо, париеталният лоб е мястото на сложен процес на разпознаване и разбиране. Дифузното увреждане на париеталните лобове завършва с агнозия, зрителна невнимание, метаморфопсия, фотопсия, халюцинации, илюзии и алексия (виж Фиг. 4.1.12, 4.1.13).
| Повече ▼ подробностиза последствията от дисфункция на париеталния лоб са дадени в неврологичната литература. Тук представяме само някои от синдромите.
зрителна агнозия, който често се развива в нарушение на кръвообращението в централната нервна система, се характеризира с нарушение на процеса на разпознаване. Агнозия може да бъде пълна или частична. Визуалната агнозия се диагностицира в случаите, когато пациентът може да идентифицира обект само тактилно. Разпознаването и разбирането на визуалните образи и цветовото значение на изображението често са нарушени.
Двустранно увреждане на париеталния лоб (по-често при сенилна деменция, омекване на темпоро-окципиталната и париеталната област на кората, наранявания на задните части на двете полукълба, енцефалит) се придружава от зрителна агнозия и зрителна атаксия (синдром на Балинт).
Визуална пространствена агнозияхарактеризиращ се с трудности при оценката на размера на обекта и разстоянието до него. Отбелязва се и появата на неправилно решение на "синтетични" пространствени проблеми като четене на карта. Едновременната агнозия се свежда до факта, че пациентите запазват способността си да разпознават отделни обекти, но нямат способността да възприемат група от обекти (изображения) като цяло.
Астерогностияхарактеризиращ се с факта, че пациентът може да анализира обекти по външния им вид, но не е в състояние да ги характеризира тактилно. Това се развива в резултат на нарушаване на сложното взаимодействие между тактилна и визуална информация в теменните дялове на кората.
Дифузните лезии на париеталните дялове могат да доведат до илюзии и метаморфопсии. Тези изкривявания включват микропсия, макропсия. Пациентите отбелязват движението на неподвижни цели, неправилно оценяват разстоянието до обектите. ДА СЕ специални видовеилюзиите включват папинопсия (нарушение на зрителното възприятие под формата на запазване или повторна поява на визуален образ след изчезването на обект от зрителното поле), халюцинаторна полиопия и др.
Илюзиите се развиват в резултат на дифузни нарушения на кората, а халюцинациите и фотопсиите се развиват с развитието на тумор или токсично увреждане на кората.
Характеристиките на патологията на париеталния лоб също могат да зависят от това дали патологичният фокус е локализиран в доминантното (обикновено ляво) или недоминантно полукълбо. Доминиращият париетален лоб е отговорен за развитието на речеви нарушения като слухова сензорна афазия, зрителна агнозияи аграфии.
Увреждане на доминиращия ъглов гирус(по-често в резултат на нарушения на кръвообращението в басейна на ъгловата артерия) развиват нистагъм, зрително-агностични нарушения, аграфия, акалкулия (неспособност за добавяне или изваждане на числа), дигитална агнозия, невъзможност да се прави разлика между дясната и лявата страна ( Синдром на Герстман; синдром на ангуларния гирус). Може би развитието на хомонимна хемианопсия.
Ако недоминиращият ъглов гирус е повреден, взаимодействието на пациента с околната среда се нарушава. В резултат на това се развива зрителна дезориентация и топографска агнозия.
Отпред на ъгловата извивка и през супратемпоралната бразда е супрамаргинален извивка. В доминантното полукълбо тази област е срещата на визуалните и слуховите компоненти на езика.
Увреждането на задните париетални асоциационни зони често завършва развитие на апраксия(затруднено изпълнение на "фини" специализирани движения). Това очевидно се дължи на неразбиране на необходимата последователност от движения за изпълнение на определена функция (т.е. за пациента не е ясно какво и в каква последователност трябва да се извърши). Пациентът не може да начертае проста диаграма (конструктивна апраксия). По-малко сериозна форма на тази патология е тактилната агнозия (неспособност за разпознаване на обект при палпация). Понякога това заболяване се нарича астерогностия. В същото време няма признаци на загуба на тактилна или проприоцептивна чувствителност и интеграцията на визуална и соматосензорна информация е нарушена. Апраксия и астерогностия често се развиват след увреждане на лявото полукълбо на мозъка. Астерогностията обикновено е ограничена до противоположната страна на тялото, докато апраксия обикновено е двустранна.
Зрителната и слуховата информация са тясно свързани с функцията на езика. Поради тази причина увреждането на области 39 и 40, както и на части от областта на слуховата асоциация, обикновено води до афазия, известна като афазия на Вернике(Вернике). Ако увреждането е ограничено до ъгловата извивка, способността за разбиране на писмен език (алексия) и способността за писане (аграфия) се губят. В същото време пациентите разбират речта. Това състояние обикновено се развива след нараняване.
Интегрирането на визуална и соматосензорна информация е важно за формирането на "образ на тялото", т.е. разбиране на характеристиките на позицията на тялото в пространството. Значително увреждане на полета 7, 39 и 40 на едното полукълбо може да доведе до развитие на "неразбиране" или "пренебрегване" на противоположната половина на тялото. Пациентите не могат да превържат или развържат собствените си крайници. Те също не разпознават собствените си крайници.
Тилен лоб(полета 17, 18 и 19). По протежение на жлеба на шпора (fissura calcarina) е основната зрителна зона (поле 17). В съседство с него е "оценъчната" визуална зона (поле 18), за която се смята, че преобразува сигналите, получени от поле 17, във визуални изображения.
Зона 17 на тилната кора е зона за приемане и обработка на визуална информацияи има важни връзкис двигателни зрителни зони на кората. Именно на това ниво на зрителния кортекс се оценява значението на това, което се вижда. Дразненето на тази област предизвиква халюцинации и съновидения.
Двигателните връзки на тилния лоб също заслужават внимание, тъй като те участват в оптокинетичния нистагъм, акомодационния рефлекс и регулацията на бинокулярното зрение.
Полета 18 и 19 са взаимозависими и имат повече връзки с останалата част от мозъка. Смята се, че зона 18 е по-заинтересована от интегрирането на визуална информация. получава се в 17-то поле, докато поле 19 превежда тази информация в по-сложна умствена и двигателна дейност.
Скорошни анатомични и физиологични изследванияразкри, че има поне, 10 подрегиона на полета 18 и 19, чиито неврони имат способността да реагират на визуални стимули. Всеки от тези подрегиони е свързан със 17-то поле. Те обаче не са свързани помежду си. Увреждането на тези подрегиони не води до слепота. Някои са нарушени зрителни функции, като анализ на цветовете, възприятие за целостта на обект и др.
зрителна асоциативна кора, разположен в някои области на темпоралния лоб, също участва в обработката на визуална информация. Това се отнася в най-голяма степен за поле 21 (в средната темпорална кора). Именно това поле осигурява разпознаване на изображението и неговото увреждане завършва с агнозия.
Горните свойства на зрителния кортекс се осигуряват и от множество хоризонтални и вертикални връзки между неврони в различни слоеве на зрителния кортекс. Благодарение на тяхното присъствие се отбелязва изключителната пластичност на зрителната кора при обработката на визуална информация от различен тип.
Структурната организация на зрителната кора и нейните функции ще бъдат дадени по-подробно в следващия раздел.
темпорален лоб(полета 41, 22).
Тази част от мозъка свързан с възприемането на слухови усещания, участва в речевата функциячрез слухов контрол на речта и също играе роля в оценката на пространството и функцията на паметта. Отдавна е установено, че напречният темпорален гирус (поле 41) е основният слухов център. Малко поле (22). заобикаляща тази извивка, се счита за "ментална" или "оценъчна", слухов център. Дразненето на по-голямата част от темпоралния лоб, особено по дължината на средния темпорален гирус, причинява усещания, които пациентът описва като слухови. Пациентите с увреждане на "менталното поле" на темпоралния лоб понякога губят способността да оценяват звуковите тонове. В същото време пациентът не разпознава мотивите, а музикалните звуци се възприемат от него като случаен шум.
Поле 22 на лявото полукълбо също е свързано с функцията на речта, тъй като увреждането на тази област води до загуба на способността да се разбира значението на думите. Има също така доказателства, че темпоралният лоб е свързан с "вестибуларната" чувствителност (усещане за баланс), тъй като стимулацията на задния горен темпорален гирус при пациент в съзнание причинява замаяност и усещане за въртене.
Дълбоките лезии в темпоралния лоб често включват най-ниските оптични радиационни влакна от латералното геникуларно тяло. При което зрително увреждане. Най-честата е хемианопсията на горния квадрант.
С поражението на темпоралния или психомоторния лоб се развива епилепсия, придружена от зрителни халюцинации, усещането за deja vu и deja pense („вече видяно“ - фр.), при което визуалното преживяване и мислите имат странна приятелска връзка, появяват се автоматизми, които съпътстват амнезия, компулсивно поведение, гняв и тревожност.
При пациенти с едностранно увреждане на първичния слухова коралека загуба на слуха. При нарушение на слуховото разпознаване (слухова агнозия) се засягат асоциативните слухови полета. Пациентите обикновено съобщават, че чуват звуци, но не могат да ги идентифицират.
Област, участваща в разпознаването на лица, се намира на вентралната повърхност на темпоралния лоб. Увреждане на тази област (по-често в нарушение на кръвообращението в областта на базиларната артерия на мозъка), което, като правило, се простира до съседния тилен лоб (полета 20, 21), двете полукълба, придружено от развитие на прозопагнозия(невъзможност за разпознаване на лица). По принцип това състояние не е придружено от други неврологични признаци. Пациентът може да чете и назовава предмети. В случаите, когато пациентът чуе гласа на познат човек, той може веднага да го идентифицира.
Останалата част от горната темпорална извивка е заета от зона 22 (слухова асоциативна кора), която получава голям брой аференти от полета 41 и 42, за да даде влакна към париеталните и тилните дялове. Зона 22 също участва в езиковите функции. Увреждането на горната част на 22-ро поле води до затруднения в разбирането на езика.
Темпоралните дялове са особено важни за функцията на паметта.. Например, стимулирането на кората на слуховите асоциации при пациенти по време на неврохирургични операции предизвиква сложни спомени, както слухови, така и зрителни. При обширно увреждане на темпоралния лоб настъпва нарушение на паметта.
лимбичен лоб(фиг. 4.1.14).
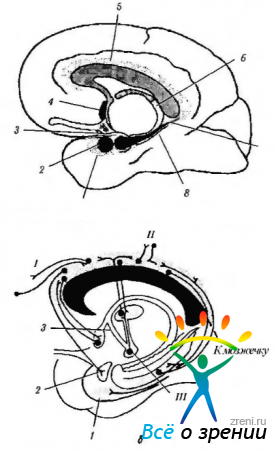
Ориз. 4.1.14.Лимбична система (според Brodsl, 1992): a - местоположението на структурите на лимбичната система (1 - енториално поле; 2 - амигдала; 3 - кука; 4 - септално ядро; 5 - cingulate gyrus; b - fornix; 7 - хипокампус; 8 - парахипокампален gyrus); b - връзки на cingulate gyrus (I - фронтална асоциативна кора; II - париетална и темпорална асоциативна кора; III - мастоидно тяло). Сингуларният гирус е свързан с асоциативните полета на кората на главния мозък и лимбичната система, осигурявайки взаимодействие между тях.
Лимбичната система е комплекс от образувания на терминала, диенцефалона и средния мозък. Всъщност " лимбична система” съвпада с използваното преди това понятие – „обонятелен мозък”.
Ако новият кортекс (неокортекс) контролира пространствено-времевите отношения на организма с околната среда и също така е отговорен за формално-логическото мислене и стереогностичните способности, тогава лимбичната система определя емоционалните функции и мотивацията за действиекакто и процеси на учене и памет. Освен това контролира хомеостазата. Неговата важна функция е организирането на поведенческите реакции на индивида в отговор на влиянието на външната среда и промените във вътрешната среда на организма. Тези реакции са насочени към запазване на индивида и се осъществяват чрез долните центрове на диенцефалона.
Лимбичният лоб също има нещо общо с функцията на обонянието, тъй като дразненето или увреждането на областта на хипокампа е придружено от обоняние.
Образува се лимбичният лоб:
- подкаузална област (area subcallosa),
- cingulate gyrus (gyrus cinguli),
- провлак на cingulate gyrus
- парахипокампална извивка (gyrus parahippocampalis),
- хипокампус (рог на амон, назъбен гирус и основа на хипокампуса или субикулум)
- и диагоналния лигамент на Broca.
Подкоровите структури на лимбичната система включват сливиците (corpus amygdaloideum), септалните ядра и предното таламично ядро.
Аферентните и еферентните връзки на структурите на лимбичната система, както помежду си, така и с други части на мозъка, са изключително разнообразни. Най-силно изразени са реципрочните му връзки с хипоталамуса.
Хипоталамус и мамиларни теласвързан с хипокампуса и септалната област чрез форникса, с амигдалата чрез крайната ивица и амигдалофугалния сноп и с обонятелния мозък чрез медиалния сноп преден мозък. Лимбичната система е свързана със средния мозък чрез хипоталамуса и мамиларните тела.
Лимбичната система комуникира с неокортекса във фронталната и темпорален лоб. Темпорални областиса основно отговорни за предаването на информация от зрителния, слуховия и соматичния кортекс към амигдалата и хипокампуса. Фронталните области служат като основен отдел на неокортекса, регулиращ лимбичните зони.
Базални ганглиипредставляват подкорово натрупване на неврони под формата на група ядра (фиг. 4.1.15; 4.1.16).
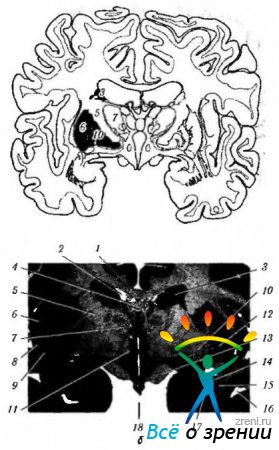
Ориз. 4.1.15.Базални ганглии: а - схематично представяне на местоположението на структурите, свързани с базалните ганглии; b - челен участък на мозъка, преминаващ през структурите, свързани с базалните ганглии (4 - corpus callosum; 2 - рог страничен вентрикул; 3- тяло на опашното ядро; 4-свод; 5-вътрешна капсула; 6 - островче; 7 - визуален туберкул; 8- черупка; 9 - ограда; 10 - бледа топка; 11 - хипотуберална област; 12 - безименно вещество: 13 - зрителен тракт; 14 - задната долна част на опашното ядро; 15 - сливица: 16 - темпорален рог на страничния вентрикул; 17 - челен рог на страничния вентрикул; 18 - трета камера)
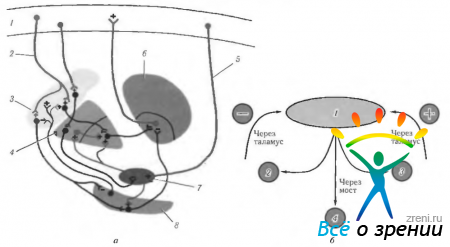
Ориз. 4.1.16.Най-важните аферентни, еферентни и вътрешни връзки на базалните ганглии и връзката им с малкия мозък (обяснение в текста): a - (1-моторна кора; 2 - еферентни влакна от кората до стриатума; 3 - стриатум; 4 - бледа топка; 5 - аферентни влакна; 6 - зрителен туберкул; 7 - субталамично ядро; 8 - substantia nigra); b - (1 - мозъчна кора (моторна); 2 - базални ганглии; 3 - малък мозък; 4-моторни и интеркаларни неврони)
Те контролират движението на тялото. Тази функция на базалните ганглии е установена въз основа на клинични наблюдения. Когато са повредени, има нарушение двигателна активносткакто под формата на невъзможност да се инициират необходимите движения, така и невъзможност за потискане на движенията.
При болестта на Паркинсон, която възниква при увреждане на базалните ганглии, пациентите имат "неизразително" лице. Това е свързано с нарушена двигателна активност на лицевите мускули и контрол на движенията на очите, по-специално нарушения на сакадичните движения. Именно поради тази причина ще разгледаме по-подробно невроанатомията на тази област.
Има три основни натрупвания на подкорови ядра, които се наричат стриатум ( corpus striatum), ограда (claustrum) и амигдала (corpus amigdaloideum).
- Corpus striatum се състои от две части - опашато ядро (nucleus caudatus) и лещовидно ядро (nucleus lentiformis).
Каудално ядролежи над и медиално на лещовидното ядро, отделено от последното от слой бяло вещество, наречен вътрешна капсула (capsula interna). Удебелената предна част на каудалното ядро, неговата глава (caput nuclei caudati) образуват страничната стена преден рогстраничен вентрикул, докато задната тънка част на каудалното ядро (corpus et cauda nuclei caudati) се простира назад по дъното на централната част на страничната камера. От медиалната страна nucleus caudatus е в непосредствена близост до визуалния хълм, отделен от него с ивица бяло вещество (stria terminalis). Отпред и отдолу главата на каудалното ядро достига предната перфорирана субстанция (substantia perforata anterior). В този момент главата е свързана с лещовидното ядро (с част, наречена putamen). В допълнение към тази широка връзка на двете ядра от вентралната страна има и тънки ивици сиво вещество, осеяни с бели снопчета на вътрешната капсула. Те са довели до наименованието "набраздено тяло" (corpus striatum).
Лещовидно ядро(nucleus lentiformis) лежи латерално от опашното ядро и зрителната могила, отделена от тях с вътрешна капсула. В хоризонтален участък на полукълбото медиалната повърхност на лентиформеното ядро, обърната към вътрешната капсула, има формата на крехкост с връх, насочен към средата. Предната страна на ъгъла е успоредна на каудалното ядро, а задната страна е успоредна на таламуса. Страничната повърхност е леко изпъкнала и е обърната към страничната страна на полукълбото в областта на инсулата. Отпред и вентрално, както вече беше посочено, лентиформеното ядро се слива с главата на опашното ядро.
На предния участък лентиформеното ядро има формата на клин, чийто връх е обърнат към медиалната страна, а основата е странично. Лещовидното ядро е разделено на три сегмента от два успоредни бели слоя (laminae medullares). Странично разположената тъмно сива зона се нарича черупка (putamen), а двете средни, по-светли, заедно се наричат бледа топка (globus pallidus).
Различавайки се вече по своя макроскопичен вид, бледа топка(globus pallidus) също има хистологична структура, която се различава значително от структурата на други части на стриатума.
С оглед на всички тези особености бледото топче се обособява в специална морфологична единица, наречена pallidum, докато обозначението striatum е оставено само за putamen и nucleus caudatus. В резултат на това терминът "лещовидно ядро" губи предишното си значение и може да се използва само в чисто топографски смисъл. В същото време, вместо предишното име corpus striatum, опашните и лещовидните ядра се наричат стриопалидарна система.
Стриопалидарната система е основната част от екстрапирамидната система и освен това е най-висшият регулаторен център автономни функциипо отношение на терморегулацията и въглехидратния метаболизъм, доминиращи над подобни вегетативни центрове в хипоталамуса.
- Ограда(claustrum) е тънка пластинка от сиво вещество, разположена в областта на острова, между него и путамена. Той е отделен от последния от слой бяло вещество, наречен външна капсула (capsula externa).
- амигдала(corpus amygdaloideum) се намира под путамена в предния край на темпоралния лоб. Амигдалата изглежда принадлежи към субкортикалните обонятелни центрове и към лимбичната система. Той завършва в снопа от влакна, идващи от обонятелния лоб и предната перфорирана субстанция (substantia perforata anterior), отбелязана в описанието на зрителната могила, наречена стрия, завършва.
Ролята на амигдалата при хората не е добре разбрана. По принцип ролята на амигдалата е разкрита въз основа на изследване на неврологични пациенти. Благодарение на тези изследвания е установено, че механичната или електрическа стимулация на амигдалата при хората често води до страх или други емоционални реакции. Така човешката амигдала участва в процесите на емоционално възприятие. Показано е също, че при двустранно увреждане на сливиците има загуба на "емоционална" памет, поява на "уплашено" изражение на лицето и нарушение на интеграцията на обонятелната и визуалната информация.
Добре известно е, че човек има способността да определя настроението и емоционалното състояние на друг индивид. IN напоследъкучастие на амигдалата в този комплекс физиологичен процес. Когато определя емоционалното състояние на човек, нашият мозък улавя и анализира фините промени в изражението на лицето и на първо място в посоката на погледа. Невронните механизми, лежащи в основата на тези процеси, не са съвсем ясни. Въпреки това е установено, че "точната" настройка на погледа, психофизиологично оценена като наличие на "внимание", определен Горна часттемпорален кортекс и амигдала. При изследване на пациенти с аутизъм беше установено, че те често имат аномалия на амигдалата или нейното увреждане. Непосредствените механизми, лежащи в основата на социалната преценка на други хора въз основа на ориентацията на погледа, остават до голяма степен неизвестни.
Напоследък substantia nigra и субталамичните ядра също се отнасят към базалните ганглии. Освен това в substantia nigra се разграничават 2 части - ретикуларна (pars reticulata) и компактна (pars compacta).
Морфологични и електрофизиологични методи показват, че повечето отаферентните сигнали, идващи към базалните ганглии, навлизат в стриатума (виж фиг. 4.1.16). Тези сигнали идват от много източници, основните от които са:
- всички области на кората полукълба;
- интраламеларни ядра на таламуса
- substantia nigra (по допаминергичния път).
От вътрешността на бледото кълбо произлиза най-важното от всички еферентни пътищабазални ганглии. Този път завършва главно в зрителния туберкул и покрива на средния мозък.
По този начин базалните ганглии играят ролята на междинна връзка във веригата, свързваща моторните зони на кората с всички останали нейни области. Основната им функция е „планиране“ на физическа активност.
IN последните годиниустанови, че базалните ганглии заедно с някои кортикални области също определят някои когнитивни функции, включително внимание, памет. Понятието "внимание", в най-широкия смисъл на думата, е "процес на избор". Като се има предвид, че процесът на ориентация в пространството и изборът на целта на интерес, в която участват сакадичните движения на очите, е свързан с базалните ганглии, става ясна значителна роля на базалния комплекс в процесите на ориентация чрез сакади. В този случай специално място в контрола на сакадите на окото играе substantia nigra, по-специално неговата ретикуларна част. Както анатомично, така и електрофизиологично е доказано наличието на връзката му с горните туберкули на квадригемината. Основната функция на невроните в substantia nigra е да инхибират активността на невроните на горния коликулус на квадригемината, участващи в генерирането на сакади.
В двигателната активност на очитеучаства и каудалното ядро, което има връзки с substantia nigra и външните туберкули на quadrigemina. Възбуждането на каудалното ядро води до сакадични движения на окото в посока, обратна на възбуденото ядро.
Субталамичното ядро също участва в движението на очите. Върху това ядро се проектира бледа топка, кората на челната област. Влакната се отклоняват от субталамичното ядро до substantia nigra, външния и вътрешния сегмент на globus pallidus.
Функцията на globus pallidus в окуломоторната активност е по-малко ясна. Това се дължи на факта, че globus pallidus има голям брой връзки между различните ядра на базалните ганглии и малко връзки с нервни центроверазположен извън базалните ганглии. Окуломоторните неврони бяха открити само в дорзалната част на globus pallidus, т.е. в частта, получаваща входни данни от опашното ядро.
Ролята на базалните ганглии в движението на очитепотвърдено от множество клинични наблюдения. При болестта на Паркинсон, придружена от дегенерация на базалните ганглии и сивото вещество на средния мозък и прекъсване на фронтомезенцефалните пътища, се развива прогресивна супрануклеарна парализа. При такива пациенти скоростта на сакадичните движения на очите е намалена. Сакадите обикновено са хипометрични, което е свързано с нарушение на функцията "сакадна памет". Често движението на очите не може да бъде завършено изцяло. Вертикалните движения на очите са засегнати по-значително от хоризонталните. Пациентите също показват аксиална ригидност и деменция. Подобни промени са открити и при болестта на Хънтингтън.
corpus callosum(корпус калозум). Corpus callosum е голям пакет от влакна (състоящ се от приблизително 250 милиона влакна), който свързва двете полукълба (фиг. 4.1.17).
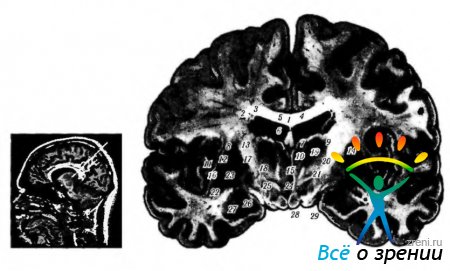
Ориз. 4.1.17.Локализация на ядрата на таламуса: а - ядрен парамагнитен резонанс; b - челен участък на мозъка (1 - тяло на corpus callosum; 2 - тяло на опашното ядро; 3 - централна част на страничния вентрикул; 4 - прозрачна преграда; 5 - хориоиден плексусстраничен вентрикул; 6 - свод; 7 - предна група ядра на зрителната могила; 8- външна капсула; 9 - странична група ядра на таламуса; 10- медиални ядра на таламуса: - черупка; 12- странична медуларна плоча; 13 - вътрешна капсула; 14 - ретикуларно ядро на таламуса; 15 - интерталамичен шип; 16 - странична част на бледата топка; 17 - медиална медуларна плоча; 18 - мамилоталамичен пакет; 19 - HI Пъстървово поле; 20 - gopa incerta; 21 - Н2 Пъстървово поле; 22 - ограда; 23-медиална част на бледата топка; 24 - трета камера; 25 - ядра на хипоталамуса; 26-визуален тракт; 27 - амигдала; 28-тяло на зърното; 29-основата на краката на мозъка)
Основната функция на този лъч е предаването на информация от едно полукълбо в друго.
Основните функции на corpus callosum са:
- Корелация на образите на дясната и лявата половина на зрителното поле.
- Интегриране на усещания, идващи от чифтни крайници, необходими за обучение на двигателна координация.
- Интегриране на процесите на внимание и активиране в полукълбата.
При пациенти, при които corpus callosum е пресечен за терапевтични цели, мозъчната дисфункция в Ежедневиетоне е спазено. Те бяха разкрити само чрез специални тестове. Други пациенти понякога развиват доста значителни поведенчески разстройства. Това най-често се проявява чрез "вербална слепота", "вербална глухота" и загуба на координация на движенията на десния и левия крайник.
Продължава в следващата статия: Анатомия на мозъка? Част 3
И то в очите на цялата столица
Петелът пърхаше от иглата за плетене,
полетя към колесницата
И седна на короната на краля,
Стреснат, клъвнал короната
И скочи... и в същото време
Дадон падна от колесницата -
Той ахна веднъж и умря.
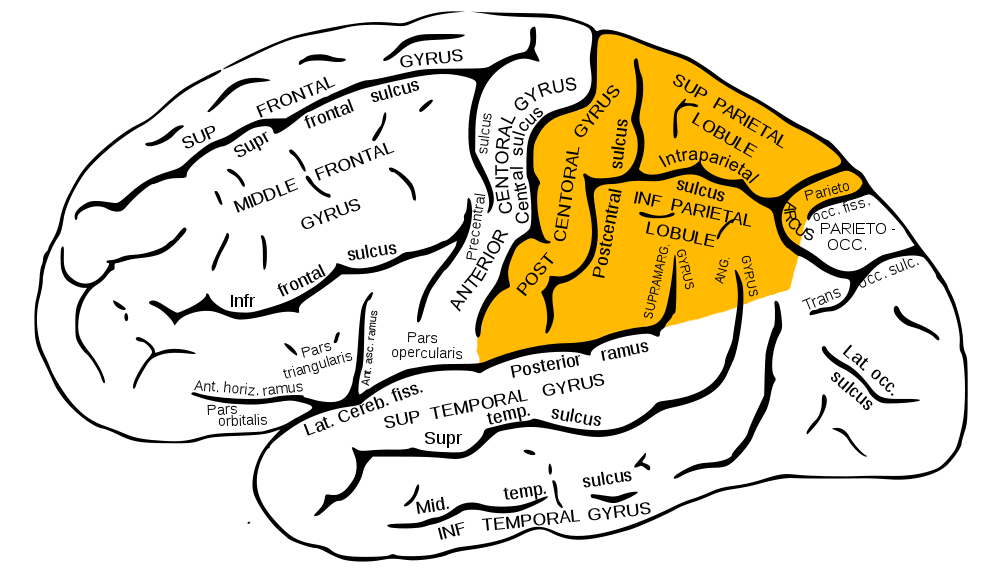
Изглежда, че вече разбрахте какво ще бъде обсъдено в следващия материал от цикъла „Как работи мозъкът“. Вече говорихме за фронталните лобове, темпоралните и париеталните, сега се обръщаме към париеталните лобове. Те също са Lobus parietalis на латински, познат на лекарите.
Париеталните лобове са показани в жълто
Париеталният лоб е разположен точно над тила и се "състои" от три извивки: една вертикална- задна централна (най-стария дял) и две хоризонтални - горна париетална (нова) и долна париетална (нова).
Подобно на структурата на предните лобове, части от човешкото тяло са „проектирани“ върху централната предна извивка на париета: долната трета е лицето, средната трета е ръката и торса, горната част е кракът. Не забравяйте, че дялът е "удвоен", следователно едната му половина е отговорна за една (срещуположна) половина на тялото.
Структурата на париеталните дялове
Освен това в горния париетален гирус има центрове, отговорни за сложни типоведълбока чувствителност: мускулно-ставен, двуизмерно-пространствен усет (дефиниране на цифри, букви, фигури, нарисувани с молив или др. тъп предметвърху човешка кожа), усещане за тежест и обем на движение, усещане за разпознаване на предмети чрез допир.
В долния париетален лоб има праксис центрове, тоест движения, които са станали „автоматични“ в процеса на повторение и упражнения, които се развиват в процеса на учене и постоянна практика, например ходене, хранене, обличане и скоро.
Париеталният лоб участва в обработката и съзнателното възприемане на соматосензорни (от рецептори в мускулите, кожата, ставите и вътрешни органи) информация, която засяга произволните движения.
Лезиите на горната париетална лобула са придружени от развитието на нарушение на способността за разпознаване на предмети чрез докосване със затворени очи. Пациентите описват индивидуалните качества на обекта, но не могат да синтезират неговия образ.
С поражението на долния париетален лобул се нарушава усещането за схемата на тялото. Човек не може да разбере къде е дясната и къде лявата страна, не разпознава собствените си пръсти. Друг вид разстройство е несъзнаването на своя дефект (пациентът твърди, че движи парализирани крайници). Тези пациенти могат да развият псевдополимелия.— усещане за допълнителни крайници или части на тялото. Такива пациенти могат самостоятелно да премахнат „пречещия“ крайник или да допринесат за неговата ампутация.
Когато кората на ъгловия гирус е повредена, пациентът губи чувството за пространствено възприемане на света около него, позицията на собственото си тяло и взаимовръзките на неговите части. Това е придружено от различни психопатологични симптоми: деперсонализация, дереализация. Те могат да се наблюдават при пълно запазване на съзнанието и критичното мислене.
При лезии на париеталния лоб на доминантното полукълбо човек развива дислексия - неспособност да чете, да прави разлика между дясно и ляво, както и дискалкулия - неспособност да прави аритметика. Трябва да се отбележи, че често дискалкулията е самостоятелно заболяване, а не следствие от неврологични или психологически проблеми. В допълнение към тези проблеми е възможна и апраксия - нарушение или невъзможност за извършване на някакво целенасочено действие (добре, например, вземете чаша и пийте), като същевременно запазите елементарните компоненти на действието.
Анастасия Шешукова
Тилният лоб заема задните части на полукълба. На изпъкналата повърхност на полукълбото тилният лоб няма резки граници, отделящи го от париеталните и темпоралните лобове, с изключение на горната част на париетално-тилната бразда, която, разположена на вътрешната повърхност на полукълбото, разделя париеталния лоб от тилния лоб. Браздите и извивките на горната странична повърхност на тилната част са нестабилни и имат променлива структура. На вътрешната повърхност на тилния лоб има шпора, която разделя клина (триъгълна лобула на тилния лоб) от лингвалния гирус и окципитотемпоралния извивка.
Функцията на тилния лоб е свързана с възприемането и обработката на визуална информация, организирането на сложни процеси на визуално възприятие. В този случай горната половина на ретината се проектира в областта на клина, който възприема светлината от долните зрителни полета; в областта на лингуларния гирус е долната половина на ретината, която възприема светлината от горните зрителни полета.
Остров
Островът или така наречената затворена лобула се намира дълбоко в страничната бразда. Островчето е отделено от съседните съседни секции с кръгъл жлеб. Повърхността на островчето е разделена от надлъжната му централна бразда на предна и задна част. Вкусов анализатор е проектиран в островчето.
лимбичен кортекс
На вътрешната повърхност на полукълбата над corpus callosum е cingulate gyrus. Тази извивка, с провлак зад corpus callosum, преминава в извивката близо до морското конче - парахипокампалната извивка. Сингуларният извивка заедно с парагитпокампалния извивка образуват сводестия извивка.
Вътрешните и долните повърхности на полукълбата са обединени в така наречената лимбична (маргинална) кора, заедно с амигдалата от групата на подкоровите ядра, обонятелния тракт и луковицата, областите на фронталните, темпоралните и париеталните лобове на мозъка. кора, както и с хипотубералната област и ретикуларната формация на багажника. Лимбичната кора е комбинирана в една функционална система - лимбично-ретикуларен комплекс. Основната функция на тези части на мозъка е не толкова да осигуряват комуникация с външния свят, колкото да регулират тонуса на кората на мозъка, нагоните и афективния живот. Те регулират сложни, многостранни функции на вътрешните органи и поведенчески реакции. Лимбично-ретикуларният комплекс е най-важната интегративна система на тялото. Лимбичната система също е важна за формирането на мотивациите. Мотивацията (или вътрешната мотивация) включва най-сложните инстинктивни и емоционални реакции (хранителни, защитни, сексуални). Лимбичната система също участва в регулирането на съня и бодърстването.
Лимбичната кора също изпълнява важна функция на обонянието. Миризмата е възприемането на химикали във въздуха. Обонятелен мозъкчовек осигурява обонянието, както и организирането на сложни форми на емоционални и поведенчески реакции. Обонятелният мозък е част от лимбичната система.
Обонятелният мозък се състои от два отдела - периферен и централен. Периферният отдел е представен от обонятелния нерв, обонятелните луковици, първичните обонятелни центрове. Централният отдел включва извивката на морското конче - хипокампуса, зъбната и сводестата извивка.
Обонятелният рецепторен апарат е разположен в носната лигавица. Чрез системата от нервни проводници информацията от рецепторите се предава в кортикалната част на обонятелния анализатор (фиг. 8).
Ориз. 8. Обонятелен анализатор (диаграма):
1 - обонятелен епител, биполярни обонятелни клетки; 2 - обонятелна крушка; 3 - обонятелен тракт; 4 - първични обонятелни центрове; 5 - визуален туберкул; 6 - кортикален обонятелен център; 7 - corpus callosum
Кортикалната секция на обонятелния анализатор се намира в cingulate gyrus, gyrus на морското конче и в куката на морското конче, които заедно образуват затворена пръстеновидна област. Периферната част на обонятелния анализатор е свързана с кортикалните области на двете полукълба.
Физиологичният механизъм на възприемане на миризма от обонятелния анализатор не е напълно изяснен. Има две основни хипотези, обясняващи природата на този процес от различни позиции. Според една от хипотезите взаимодействието между молекулите на одоранта и хеморецепторите става като ключ и ключалка, т.е. видът на молекулата съответства на специален рецептор. Друга хипотеза се основава на предположението, че молекулите на миризлива субстанция имат определена вълна на колебание, към която обонятелните рецептори са „настроени“. Молекулите, които имат подобни вибрации, трябва да имат обща вълна и съответно да дават подобни миризми.
Терминът "обонятелен мозък" по отношение на човешката физиология е донякъде произволен и не разкрива напълно неговата многостранна и универсална функция. „Поставянето“ на централното звено на обонятелния мозък в мозъчните полукълба не е случайно и е резултат от огромната „информационна“ роля, която е изиграло обонянието в процеса на еволюцията при адаптирането към външната среда и регулирането на комплекса. поведенчески реакции. Получаване на храна, избор на индивид от противоположния пол, грижа за потомството, целостта на територията, организиране на групови общности в рамките на един вид - всички тези ежедневни функции при много животни се извършват с прякото участие на фино изградена система за обонятелна рецепция и , въз основа на това, способността на редица животни да изпращат фини диференцирани специфични миризливи вещества- сигнали-информатори.
Универсалните форми на поведенчески реакции при животните, проявяващи се в ежедневната грижа за местообитанието, за потомството, създават впечатлението, че те са надарени с разум. Привидната интелигентност е просто резултат от реагиране на външни стимули. Но самите тези стимули и реакциите към тях напълно отговарят на биологичните нужди на животните.
В живота на хората обонянието е загубило биологичната информационна стойност, която е имало при животните. Човешката обонятелна система е проектирана както да изпълнява тясна, „собствена“ функция, така и за един вид „зареждане“ на емоции. Силата на въздействието на миризмите върху емоционалната сфера, че те са най-важният „хранителен субстрат на емоциите“, е добре известна от древността в историята на човечеството.
Обонянието на човек може да варира. По правило тези вариации са незначителни, но в някои случаи остротата на миризмата може да бъде много висока (дегустатори в парфюмерийната индустрия).
Тъй като обонятелният анализатор играе важна роля в регулирането на емоциите, неговият централен участък се отнася към лимбичната система, образно наречена „общ знаменател“ за много емоционални и висцеросоматични реакции на тялото.
Големи полукълба на мозъка
са най-голямата част от мозъка. Те покриват малкия мозък и мозъчния ствол. Мозъчните полукълба съставляват приблизително 78% от общата маса на мозъка.
В процеса на онтогенетичното развитие на организма мозъчните полукълба се развиват от крайния мозъчен мехур на невралната тръба, поради което тази част на мозъка се нарича още теленцефалон.
Мозъчните полукълба се делят на средна линиядълбока вертикална цепка отдясно и ляво полукълбо. В дълбочината на средната част двете полукълба са свързани помежду си с голяма адхезия - corpus callosum. Всяко полукълбо има дялове: челен, париетален, времеви, тилен.
Лобовете на мозъчните полукълба са разделени един от друг с дълбоки бразди. Най-важни са три дълбоки бразди: централната (на Роланд), разделяща фронталния дял от теменния; латерална (Sylvian), отделяща темпоралния лоб от париеталния, и париетално-окципитален, разделящ теменния лоб от тилната на вътрешната повърхност на полукълбото.
Всяко полукълбо има горно-латерална (конвексна) - конвекситална, долна - базална и вътрешна - медиална повърхност. Всеки лоб на полукълбото има мозъчни извивки, разделени една от друга с бразди. Отгоре полукълбото е покрито с кора - тънък слой сиво вещество, което се състои от нервни клетки.
Cortex- най-младата формация на централната нервна система в еволюционно отношение. При човека достига най-високо развитие. Мозъчната кора е от голямо значение за регулирането на жизнената дейност на тялото, за осъществяването на сложни форми на поведение и формирането на нервно-психични функции.
Под кората е бели кахъриполукълба, той се състои от процеси на нервни клетки - проводници. Поради образуването на мозъчни извивки, общата повърхност на мозъчната кора се увеличава значително. Общата площ на кората на полукълба е 1200 cm2, като 2/3 от повърхността му е разположена в дълбините на браздите и 1/3 на видимата повърхност на полукълбата. Всеки дял на мозъка има различно функционално значение.
Cortex се състои от 4 дяла, разделени един от друг с бразди. Основните бразди, разделящи фронталните, париеталните и темпоралните лобове, са Роландов и Силвиев.
Лобове на мозъчната кора :
челен (челен ),
париетален (париетален ),
тилен (тилен ),
времеви (времеви ),
Във връзка с акциите се говори за локални системи на мозъка.
МЕСТНИ СИСТЕМИ НА МОЗЪКА:
Тилната област на мозъкаизпълняват функциите на организиране на визуалното възприятие. Първични зони на тилната кора - елементарни функциивизия.
Вторични отдели на тилната кора - оптико-гностични функции.
Темпорални области на мозъка"отговорен" за организацията на слуховото възприятие. Основните зони на темпоралната кора са елементарните функции на слуха.
Вторични зони на темпоралната кора - акустико-гностични функции.
Сензомоторни и премоторни части на мозъка- организация на движенията. 1. Постцентрални части на мозъка - аферентна организация на движенията. 2. Премоторни зони на кората - еферентна организация (програмиране) на движението.
3. Моторни зони на кората - двигателен анализатор, изпълнение на двигателна програма. Фронтални дялове на мозъка (префронтални области)извършват регулирането на умствената дейност, т.е. регулиране на състояния на активност, произволни движения и действия, когнитивни процеси и емоционална и лична сфера, като цяло са отговорни за мисленето, интелектуалната дейност. Париетални области на мозъкаса представени от първични зони (всички видове кожно-кинестетична чувствителност), вторични зони (визуално-пространствени представи, представи за схемата на тялото, соматогноза, стереогнозис) и третични зони.
челен дялзаема предните части на полукълба. Той е отделен от теменния лоб от централната бразда, а от темпоралния лоб от страничната бразда. Във фронталния лоб има четири извивки: една вертикална - прецентрална и три хоризонтални - горна, средна и долна фронтална извивка.
Свивките са разделени една от друга с бразди. На долната повърхност на фронталните лобове се разграничават директният и орбиталният гирус. Директният гирус се намира между вътрешния ръб на полукълбото, обонятелния жлеб и външния ръб на полукълбото.
В дълбините на обонятелната бразда лежат обонятелната луковица и обонятелният тракт. Фронталния лоб на човек съставлява 25-28% от кората, средната маса на челния лоб е 450 g.
Функцията на фронталните лобове е свързана с организацията на произволните движения, двигателните механизми на речта, регулирането на сложни форми на поведение и мисловни процеси. Няколко функционално важни центъра са концентрирани в извивките на фронталния лоб. Предният централен гирус е "представяне" на първичната двигателна зона със строго определена проекция на частите на тялото. Лицето е "разположено" в долната трета на гируса, ръката в средната трета, кракът навътре горна трета. Багажникът е представен в задните отдели на горния фронтален гирус. По този начин човек се проектира с главата надолу и надолу в предния централен гирус.
Преден централен гирусзаедно със съседните задни участъци на фронталните извивки, той играе много функционално важна роля. Той е центърът на доброволните движения. В дълбините на кората на централния гирус от т.нар пирамидални клетки -централен двигателен неврон- започва основният двигателен път - пирамидалният или кортикоспиналният път. Периферните процеси на моторните неврони излизат от кората, събират се в един мощен сноп, преминават през централното бяло вещество на полукълбата и навлизат в мозъчния ствол през вътрешната капсула; в края на мозъчния ствол те частично се пресичат (преминават от едната страна на другата) и след това се спускат в гръбначен мозък. Тези клонове завършват в сива материягръбначен мозък. Там те влизат в контакт с периферния двигателен неврон и му предават импулси от централния двигателен неврон. Импулсите на произволно движение се предават по пирамидалния път.
В задните отдели на горната фронтална извивка има и екстрапирамидален център на кората, който е тясно свързан анатомично и функционално с образуванията на така наречената екстрамирамидна система. Екстрапирамидна система- двигателна система, която подпомага осъществяването на произволно движение. Това е система за "осигуряване" на произволни движения. Филогенетично по-стара от пирамидната система, човешката екстрапирамидна система осигурява автоматично регулиране на „заучените“ двигателни действия, поддържане на общия мускулен тонус, „готовност“ на периферния двигателен апарат за движение, преразпределение на мускулния тонус по време на движения. Освен това участва в поддържането на нормална поза.
В задната част на средната фронтална извивка е фронталният окуломоторен център, който контролира приятелското, едновременно въртене на главата и очите (центърът на въртене на главата и очите в обратна посока). Дразненето на този център води до обръщане на главата и очите в обратна посока. Функцията на този център е от голямо значение за осъществяването на така наречените ориентировъчни рефлекси, които са много важни за запазването на живота на животните.
В задната част на долната фронтална гирус е двигателен център на речта(центърът на Брок).
Фронталната кора на мозъчните полукълба също получава Активно участиевъв формирането на мислене, организирането на целенасочени дейности, дългосрочното планиране.
темпорален лобзаема долната странична повърхност на полукълбата. Темпоралният лоб е отделен от фронталния и париеталния дял чрез странична бразда.
На горната странична повърхност на темпоралния лоб има три извивки - горен, средно аритметичноИ нисък. Горната темпорална извивка е разположена между силвиевата и горната темпорална бразда, средната извивка е между горната и долната темпорална бразда, а долната извивка е между долната темпорална бразда и напречната церебрална фисура. На долната повърхност на темпоралния лоб се разграничават долната темпорална извивка, латералната окципитотемпорална извивка, хипокампалната извивка (крака на морско конче).
Функция на темпоралния лобсвързани с възприемането на слухови, вкусови, обонятелни усещания, анализ и синтез на звуци на речта, механизми на паметта. Основният функционален център на горната странична повърхност на темпоралния лоб е разположен в горната темпорална извивка. Тук е слуховият или гностичен център на речта (центърът на Вернике).
В горната темпорална извивка и на вътрешната повърхност на темпоралния лоб е зоната на слуховата проекция на кората. Зоната на обонятелната проекция се намира в гируса на хипокампа, особено в предната му част (така наречената кука). До обонятелните проекционни зони има и вкусови.
Темпоралните дялове играят важна роля в организацията на комплекса умствени процеси, по-специално памет.
париетален лобзаема горните странични повърхности на полукълбото. От предния париетален лоб, отпред и отстрани, той е ограничен от централната бразда, от темпоралната отдолу - от страничната бразда, от тилната - от въображаема линия, минаваща от горния ръб на париетално-тилната бразда до долния ръб на полусферата.
На горната странична повърхност на париеталния лоб има три извивки: една вертикална - задна централна и две хоризонтални - горна париетална и долна париетална. Частта от долния париетален извивка, която обгръща задната част на латералната бразда, се нарича супрамаргинална (супрамаргинална), а частта, обграждаща горната темпорална извивка, възлова (ъглова) област.
Париеталният лоб, подобно на фронталния лоб, съставлява значителна част от мозъчни полукълба. Във филогенетично отношение в него се разграничава стар участък - задната централна извивка, нова - горната париетална извивка и по-нова - долната париетална извивка. Функцията на париеталния лоб е свързана с възприемането и анализа на чувствителни стимули, пространствена ориентация. Няколко функционални центъра са концентрирани в извивките на париеталния лоб.
В задния централен гирус центровете на чувствителност се проектират с проекция на тялото, подобна на тази в предния централен гирус. В долната трета на гируса се проектира лицето, в средната трета - ръката, торса, в горната трета - крака. В горния париетален гирус има центрове, които отговарят за сложни видове дълбока чувствителност: мускулно-ставно, двуизмерно-пространствено усещане, усещане за тежест и обем на движение, усещане за разпознаване на обекти чрез докосване.
По този начин кортикалната част на чувствителния анализатор е локализирана в париеталния лоб.
Праксисните центрове са разположени в долния париетален лоб. Праксисът се разбира като целенасочени движения, които са станали автоматизирани в процеса на повторения и упражнения, които се развиват в процеса на учене и постоянна практика през индивидуалния живот.
Ходене, хранене, обличане, механично писане, различни видоветрудовата дейност (например движенията на водача за управление на кола, косене и др.) са праксис.
Праксис- най-висшата проява на двигателната функция на човека. Осъществява се в резултат на комбинираната дейност на различни територии. мозъчната кора.
Тилен лобзаема задните области на полукълбата. На изпъкналата повърхност на полукълбото тилният лоб няма резки граници, отделящи го от париеталните и темпоралните лобове, с изключение на горната част на париетално-тилната бразда, която, разположена на вътрешната повърхност на полукълбото, разделя париеталния лоб от тилния лоб.
Браздите и извивките на горната странична повърхност на тилната част са нестабилни и имат променлива структура.
На вътрешната повърхност на тилния лоб има шпора, която разделя клина (триъгълна лобула на тилния лоб) от лингвалния гирус и окципитотемпоралния извивка.
Функцията на тилния дял е свързана с възприятието и обработката визуална информация, организация на сложни процеси на зрително възприятие. В този случай горната половина на ретината се проектира в областта на клина, който възприема светлината от долните зрителни полета; в областта на лингуларния гирус е долната половина на ретината, която възприема светлината от горните зрителни полета.
Остров, или т.нар затворена лобула, намира се в дълбочината на страничната бразда. Островчето е отделено от съседните съседни секции с кръгъл жлеб.
Повърхността на островчето е разделена от надлъжната му централна бразда на предна и задна част. В о вкусов анализатор.
Обонятелен анализатор
Нервни клетки, възприемащи обонятелни дразнения, се намират в лигавицата на горните части на носната кухина. Оттук аксоните на тези клетки отиват в черепната кухина и влизат в обонятелните луковици. От тях се изпращат нервни влакна темпорален лоб (вътрешна повърхност)къде са нервните клетки обонятелен анализатор.
Анализатор на вкуса
Този анализатор започва в нервните окончания на вкусовите рецептори на езика, които са представени в тях от вкусови рецептори. Нервните влакна, напускащи вкусовите рецептори, отиват в мозъка и завършват, подобно на нервите на обонятелния анализатор, на вътрешна повърхност темпорален лоб.
corpus callosum- дъговидна тънка пластина, филогенетично млада, свързва средните повърхности двете полукълба. Удължената средна част на corpus callosum преминава в удебеляване отзад, а отпред се извива и извива дъговидно надолу.
Corpus callosum свързва филогенетично най-младите части на полукълбата и играе важна роля в обмена на информация между тях.
МОЗЪЧЕН СТЪБЛО ИЛИ МОЗЪЧЕН СТЪБЛО -
традиционно обособена система от мозъчни области, която е разширена формация, която продължава гръбначния мозък.
Мозъчният ствол винаги включва продълговатия мозък, моста и средния мозък. Често включва малкия мозък, понякога диенцефалона.
Медула -
отдел на мозъка. Съществува и традиционното име bulbus (луковица, поради формата на този отдел).
Продълговатият мозък навлиза в мозъчния ствол.
Отвън, от вентралната (лицева) страна, има пирамиди (те съдържат кортикоспиналния тракт - пътя от кората до моторните неврони на гръбначния мозък) и маслини (те съдържат ядрата на долната маслина, свързани с поддържането на баланса) . От дорзалната страна: тънки и клиновидни снопове, завършващи с туберкули на тънките и клиновидни ядра (превключете информацията за дълбока чувствителност съответно на долната и горната половина на тялото), долната половина на ромбовидната ямка, което е дъното на четвъртия вентрикул и въжените тела, които го разделят, или долните крака на малкия мозък.
Вътре са и ядрата от VIII до XII (и едно от ядрата VII) на черепните нерви, част от ретикуларната формация, медиалната бримка и други възходящи и низходящи пътища.
Има формата на пресечен конус.
Благодарение на изследванията на учени като R. Magnus и I. F. Klein беше установено, че в продълговатия мозък има сложна система от рефлексни центрове, които осигуряват определено положение в тялото поради статични и статично-кинетични рефлекси. Тези рефлекси всъщност са механизми за преразпределение на мускулния тонус по такъв начин, че да се поддържа удобна поза за животното (постурално-тонични рефлекси) или връщане към дадена от неудобна (коригиращи рефлекси) и балансът се поддържа и по време на ускорението (стато-кинетични рефлекси). Изпълнението на тези рефлекси става с участието на такива образувания на багажника като ретикуларната формация, червеното ядро и вестибуларните ядра.
Ретикуларна формация - това е формация, която върви от гръбначния мозък към таламуса в рострална (кортекс) посока. Освен че участва в обработката на сензорната информация, ретикуларната формация има активиращ ефект върху кората на главния мозък, като по този начин контролира дейността на гръбначния мозък. За първи път механизмът на действие на ретикуларната формация върху мускулен тонусе установен от R. Granit: той показа, че ретикуларната формация е в състояние да промени активността на γ-моторните неврони, в резултат на което ихаксоните (γ-еференти) причиняват свиване на мускулните вретена и в резултат на това увеличаване в аферентни импулси от мускулни рецептори. Тези импулси, влизайки в гръбначния мозък, предизвикват възбуждане на α-моторните неврони, което е причина за мускулния тонус.
Установено е, че в изпълнението на тази функция на ретикуларната формация участват два клъстера от неврони: неврони на ретикуларната формация на моста и неврони на ретикуларната формация на продълговатия мозък. Поведение на невроните на ретикуларната формация продълговатия мозъкподобно на поведението на невроните в ретикуларната формация на моста: те предизвикват активирането на α-моторните неврони на мускулите флексори и следователно инхибират активността на α-моторните неврони на мускулите екстензори. Невроните на ретикуларната формация на моста действат точно обратното, възбуждат α-моторните неврони на екстензорните мускули и инхибират активността на α-моторните неврони на флексорните мускули. Ретикуларната формация има връзка с малкия мозък (част от информацията от него отива към невроните на продълговатия мозък (от ядрата на корковия и сферичен малък мозък) и от палатката към невроните на моста) и с мозъчна кора, от която получава информация. Това предполага, че ретикуларната формация е колектор на неспецифичен сензорен поток, вероятно участващ в регулирането на мускулната активност.
важно функционално значение ретикуларен, или мрежеста формация на мозъчния ствол, която се развива във връзка с появата на система от вагус, вестибуларен и тригеминални нерви.
Ретикуларната формация се състои от нервни клетки с различна големина и форма, както и гъста мрежа нервни влакна, отиващи в различни посоки и разположени главно в близост до камерната система. На ретикуларната формация се придава първостепенно значение в кортикално-подкоровите взаимоотношения. Намира се на средни етажи. продълговатия мозък,хипоталамус, тегментум сиво вещество, мост.
Многобройни колатерали от всички аферентни (сензорни) системи се приближават до ретикуларната формация. Чрез тези колатерали всяко дразнене от периферията, насочено към определени зони на кората по специфични пътища на нервната система, достига и до ретикуларната формация. Неспецифичните възходящи системи (т.е. пътища от ретикуларната формация) осигуряват възбуждане на мозъчната кора, активиране на нейната дейност.
Наред с възходящите неспецифични системи през мозъчния ствол преминават низходящи неспецифични системи, които влияят на гръбначните рефлексни механизми.
Ретикуларната формация е тясно свързана с лимбичната система, както и с кората на главния мозък. Благодарение на това се формира функционална връзка между висшите части на централната нервна система и багажника. мозък. Тази система се нарича лимбично-ретикуларен комплекс или лимбично-ретикуларна ос. Този сложен структурно-функционален комплекс осигурява интегрирането на най-важните функции, в изпълнението на които участват различни части на мозъка.
Известно е, че будното състояние на кората се осигурява от специфични и неспецифични системи. Реакцията на активиране се поддържа от постоянна доставка на импулси от рецепторите слухови, визуален, обонятелен, вкусови кожно-кинестетични анализатори. Тези стимули се предават по специфични аферентни пътища към различни части на кората. От всички абитуриенти таламус, а след това многобройни колатерали към ретикуларната формация се отклоняват в кората на мозъчните полукълба на аферентните пътища, което осигурява неговата възходяща активираща активност.
От своя страна ретикуларната формация получава импулси от малък мозък, подкорови ядра, лимбична системакоито осигуряват емоционално адаптивни поведенчески реакции, мотивационни форми на поведение. Въпреки това, степента на осигуряване на адаптивни безусловни рефлексни реакции от неспецифична система при хора и животни е различна. Ако при животните подкоровите образувания и лимбичната система играят водеща роля в задоволяването на жизнените нужди на организма за неговото оцеляване при заобикаляща среда, тогава при хората, поради доминирането на кората, дейността на дълбоките структури на мозъка (подкорови образувания, лимбична система, ретикуларна формация) в по-голяма степен, отколкото при животно, е подчинена на мозъчната кора. Ретикуларната формация играе важна роля в регулацията на мускулния тонус. Мускулният тонус се регулира от два вида ретикулоспинални пътища. Бързопроводимият ретикулоспинален тракт регулира бързите движения; бавно провеждащ ретикулоспинален път - бавни тонични движения.
Ретикуларната формация на продълговатия мозък участва в появата на децеребрална ригидност. При пресичане на мозъчния ствол над продълговатия мозък намалява активността на невроните, които имат инхибиторен ефект върху моторните неврони на гръбначния мозък, което води до рязко повишаване на тонуса на скелетните мускули.
Функции на продълговатия мозък
Защитни рефлекси (например кашлица, кихане).
Жизненоважни рефлекси (напр. дишане).
регулиране на съдовия тонус.
Рефлексни центрове на продълговатия мозък:
защитно (кашляне, кихане и др.)
центрове за контрол на тона скелетни мускулиза поддържане на стойката на човек.
съкращаване или удължаване на времето на спиналния рефлекс
храносмилане
сърдечна дейност
Pons
Варолиевият мост (от името на Констанцо Варолиа), или мостът - част от мозъка, заедно с малкия мозък е част от задния мозък. принадлежи на мозъка,
В основата на моста има низходящи пътеки: кортикоспинален пирамидален тракт, кортикобулбарен, кортико-мостови трактове.
среден мозък(lat.Mesencephalon) -част от мозъка, древният визуален център. Включени в мозъчния ствол.
функции на средния мозък
1. мотор,
2. сензорни (зрение, слух),
3. Регулиране на актовете на дъвчене и преглъщане,

