рефлекс- отговор на тялото на дразнене на рецепторите, извършвано от нервната система.
рефлексна дъга - пътя, по който преминава нервният импулс по време на изпълнението на рефлекса. Състои се от 5 части:
- рецептор - чувствителна формация, способна да реагира на определен вид стимул;
- чувствителен неврон провежда импулси към мозъка;
- интеркаларният неврон свързва чувствителни и изпълнителни неврони, намира се в гръбначния мозък или мозъка;
- изпълнителен (двигателен) неврон провежда импулси от мозъка към изпълнителните органи;
- изпълнителна агенция- мускул (контрахира), жлеза (секрети) и др.
Безусловни рефлекси
- присъства от раждането
- по време на живота те не се променят и не изчезват;
- са еднакви във всички организми от един и същи вид;
- адаптиране на тялото към постоянни условия.
Условни рефлекси
- придобити по време на живота;
- може да се промени или изчезне;
- всеки организъм има свой собствен;
- адаптиране на тялото към променящите се условия.
Тестове
1) Коя връзка служи като начало на рефлексната дъга
А) интеркаларен неврон
Б) чувствителен неврон
Б) рецептор
Г) изпълнителен неврон
2. В каква последователност компонентите на рефлексната дъга се включват в изпълнението на рефлекса
А) изпълнителен орган, двигателен неврон, интеркаларен неврон, сетивен неврон, рецептор
Б) интеркаларен неврон, сензорен неврон, двигателен неврон, рецептор, изпълнителен орган
В) рецептор, сензорен неврон, интеркаларен неврон, двигателен неврон, изпълнителен орган
Г) сензорен неврон, интеркаларен неврон, рецептор, изпълнителен орган, двигателен неврон
3. Началният етап на рефлексната дъга в слюнчения рефлекс
а) слюнчена жлеза
Б) рецептор
Б) интеркаларен неврон
Г) изпълнителен неврон
4. Действието на стимулите предизвиква появата нервен импулс V
А) сензорни неврони
Б) двигателни неврони
Б) рецептори
Г) интеркаларни неврони
5. Възбуждането е насочено по чувствителния неврон
А) към централната нервна система
Б) към изпълнителния орган
Б) към рецепторите
Г) към мускулите
6. Рецепторите са чувствителни образувания, които
А) предават импулси към централната нервна система
Б) предават нервни импулси от интеркаларните неврони към изпълнителната
В) възприемат стимули и превръщат енергията на стимулите в процес на нервно възбуждане
Г) получават нервни импулси от сетивните неврони
7. Нервните импулси се предават към мозъка чрез неврони
А) мотор
Б) интеркаларен
Б) чувствителен
Г) изпълнителен
8. Краища на рефлексната дъга
А) изпълнителен директор
Б) чувствителен неврон
Б) рецептор
Г) интеркаларен неврон
9. Провеждат се нервни импулси от рецепторите към централната нервна система
А) сензорни неврони
Б) двигателни неврони
В) сензорни и двигателни неврони
Г) интеркаларни и моторни неврони
10. Нервни образуванияв човешкото тяло, възприемайки дразнения от външната среда, са
А) нерви
Б) рецептори
Б) мозъка
Г) нервни възли
11. Трансформацията на стимулите в нервен импулс става в
а) двигателни нерви
Б) мозъка
Б) рецептори
Г) гръбначен мозък
12. Нервните импулси се предават от човешките сетива чрез неврони
А) чувствителен
Б) двигател
Б) вмъкване
Г) изпълнителен
13. Възприема информация и я преобразува в нервни импулси
А) рецептори
Б) нервни възли
Б) изпълнителни органи
Г) интеркаларни неврони
14. Условен рефлекс
А) се наследява от потомството, но не продължава през целия живот
Б) лесно придобити и изгубени от тялото по време на живота
В) се наследява от потомството от родителите и продължава
Г) е постоянна реакция на организма към строго определени стимули от околната среда
15. Пример за условен рефлекс при котка
А) отделяне на слюнка по време на хранене
Б) отдръпване на лапата по време на изгаряне
Б) реакция на псевдонима
Г) реакция на остър звук
16. Слюноотделяне при хората при вида на лимон - рефлекс
А) условно
Б) безусловно
Б) защитна
Г) показателен
17) Условните рефлекси при хората се формират в процеса
А) индивидуално развитие
Б) видообразуване
В) историческо развитие
Г) образуване на ембриони
18. Пример за условен рефлекс е
А) слюноотделяне при вида на нарязан лимон
Б) слюноотделяне по време на хранене
В) свиване на зеницата при излагане на силна светлина
Г) отдръпване на ръката с внезапна инжекция
19. Реакцията на детето на бутилка мляко е рефлекс, който
А) се предава по наследство
Б) се образува без участието на кората полукълба
Б) се придобиват в течение на живота
Г) продължава през целия живот
20. Защитен рефлекс на кихане
А) не се наследява
Б) е условно
Б) отслабва с времето
Г) характерен за всички индивиди от вида
21. Наричат се реакции, придобити от хората и животните по време на живота и осигуряващи адаптация към променящите се условия на околната среда
А) условни рефлекси
Б) инстинкти
Б) безусловни рефлекси
Г) рефлекси, които се предават по наследство
22. Безусловен рефлекс
А) се предава по наследство
Б) придобити през живота
C) се произвежда за определени сигнали
Г) е в основата на различни външни сигнали
23. Безусловните рефлекси на хората и животните осигуряват
В) овладяване на нови двигателни умения от тялото
24. Човешка реакция към зелен цвятсветофара е рефлекс
А) вродени
Б) придобити
Б) безусловно
Г) наследени
25. Маркирайте стомашен сокв отговор на рецепторите за хранителни стимули устната кухина
А) вълнение
Б) спиране
Б) безусловен рефлекс
Г) саморегулация
26. Биологичното значение на условните рефлекси за организма е, че те
А) поддържане на постоянството на вътрешната среда
Б) специфични за вида и се запазват през целия живот
Б) насърчаване на взаимодействието функционални системи
Г) осигуряват адаптация към променящите се условия на околната среда
27. В процеса на живот животните придобиват рефлекси
А) безусловно
Б) наследени
В) характерни за всички индивиди от даден вид
Г) позволявайки им да оцелеят при променящи се условия
28. Условен рефлекс при хората е
А) свиване на зеницата при ярка светлина
Б) завъртане на главата до остър звук
В) отделяне на слюнка, когато храната попадне в устата
Г) реакция към значението на думата
29. Пример за условен рефлекс е
1) способността да карате колело
2) слюноотделяне при хранене
3) желание за сън при умора
4) кихане и кашляне
30. Условните рефлекси на хората и животните осигуряват
А) адаптация на организма към постоянни условия на околната среда
Б) адаптиране на тялото към променящия се външен свят
В) развитието на организмите на нови двигателни умения
Г) разграничаване на животните от командите на дресьора
31. Вродени рефлекси
А) са видови характеристики на даден организъм
Б) характерни за отделни индивиди от вида
В) изискват допълнителни условия за изпълнение
Г) не се наследяват
основна форма нервна дейносте рефлекс. Рефлекс - причинно-следствена реакция на тялото към промени във външната или вътрешната среда, осъществявана със задължителното участие на централната нервна системав отговор на рецепторна стимулация. Благодарение на рефлексите възниква възникването, промяната или спирането на всяка дейност на тялото.
Нарича се нервният път, по който се разпространява възбуждането по време на изпълнението на рефлексите рефлексна дъга.
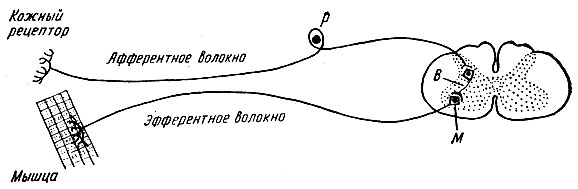
Рефлексните дъги се състоят от пет компонента: 1) рецептор; 2) аферентен нервен път; 3) рефлексен център; 4) еферентен нервен път; 5) ефектор (работно тяло).
Рецептор- Това е чувствително нервно окончание, което възприема дразненето. В рецепторите енергията на стимула се преобразува в енергия на нервен импулс. Разграничете: 1) екстерорецептори- възбудени под въздействието на дразнения от околната среда (рецептори на кожата, очите, вътрешно ухо, назална и устна лигавица); 2) интерорецептори- възприемат стимули от вътрешната среда на тялото (рецептори вътрешни органи, съдове); 3) проприорецептори- реагират на промяна в положението на отделни части на тялото в пространството (рецептори на мускули, сухожилия, връзки, ставни торби).
Аферентен нервен пътпредставени от процеси на рецепторни неврони, които пренасят възбуждания към централната нервна система.
рефлексен центърсе състои от група неврони, разположени на различни нива на централната нервна система и предаващи нервни импулси от аферентния към еферентния нервен път.
еферентен нервен пътпровежда нервните импулси от централната нервна система към ефектора.
Ефектор- изпълнителен орган, чиято дейност се променя под въздействието на нервни импулси, които идват към него чрез образуванията на рефлексната дъга. Ефекторите могат да бъдат мускули или жлези.
рефлексни дъгиможе да бъде проста или сложна. Простата рефлексна дъга се състои от два неврона - възприемащ и ефекторен, между които има един синапс. Диаграма на такава двуневронна рефлексна дъга е показана на фиг. 71.
Пример за проста рефлексна дъга са сухожилните рефлексни дъги, като пателарната рефлексна дъга.
Рефлексните дъги на повечето рефлекси включват не два, а по-голям брой неврони: рецептор, един или повече интеркаларни и ефекторни. Такива рефлексни дъги се наричат сложни, мулти-невронни. Диаграма на сложна (три невронна) рефлексна дъга е показана на фиг. 72.
Сега е установено, че по време на реакцията на ефектора се възбуждат множество нервни окончания, присъстващи в работния орган. Сега нервните импулси от ефектора отново влизат в централната нервна система и я информират за правилната реакция на работния орган. Така рефлексните дъги не са отворени, а пръстеновидни образувания.
Рефлексите са много разнообразни. Те могат да бъдат класифицирани според редица критерии: 1) биологично значение(хранителни, защитни, сексуални); 2) в зависимост от вида на раздразнените рецептори: екстероцептивни, интероцептивни и проприоцептивни; 3) по естеството на реакцията: двигател или мотор (изпълнителен орган - мускул), секреторен (ефектор - желязо), вазомотор (свиване или разширяване на кръвоносните съдове).
Всички рефлекси на целия организъм могат да бъдат разделени на две големи групи: безусловно и условно. Разликите между тях ще бъдат разгледани в глава XII.
Дейността на нервната система има рефлекторен характер. Дори през XVII век. Френският философ и математик Рене Декарт описва рефлексния акт. Той отбеляза реакцията на тялото към дразнене и предположи съществуването на път, по който преминава нервната възбуда. Самият термин "рефлекс" е предложен по-късно - през 18 век - от чешкия учен Й. Прохазка (от лат. "reflex" - отразено действие). По-късно И. М. Сеченов в работата си „Рефлексите на мозъка“ доказва, че реакциите на нервната система към различни видоведразненията протичат по рефлекторния механизъм, т.е. всички съзнателни и несъзнателни действия имат рефлексен произход. Под рефлекс се разбира специфична реакция на тялото към дразнител от вътрешната среда или получен отвън със задължителното участие на централната нервна система. Рефлексите обикновено се наричат функционални единици на нервната дейност.
Механизми на рефлексите на рефлексната дъга
Рефлексите могат да бъдат класифицирани според различни критерии. Така че, в зависимост от нивото на затваряне на дъгата, т.е. според местоположението на рефлексния център рефлексите се разделят на спинални (рефлексът се затваря в гръбначния мозък), булбарни (рефлексен център - медула), мезенцефални (структурата на рефлексната дъга е затворена в средния мозък), диенцефални и кортикални рефлексни центрове са разположени в теленцефалони съответно мозъчната кора. Според ефекторния признак те са соматични, когато еферентният път на рефлекса осигурява двигателна инервация на скелетните мускули, и вегетативни, когато вътрешните органи са ефектори. В зависимост от вида на раздразнените рецептори, рефлексите се разделят на екстероцептивни (ако рецепторът възприема информация от външната среда), проприоцептивни (рефлексната дъга започва от рецепторите на опорно-двигателния апарат) и интероцептивни (от рецепторите на вътрешните органи). Интероцептивните рефлекси от своя страна се делят на висцеро-висцерални (рефлексна дъга свързва два вътрешни органа), висцеро-мускулни (рецепторите са разположени върху мускулно-сухожилния апарат, ефекторът е вътрешен орган) и висцеро-кутанни (рецепторите са локализирани в кожата, работните органи са вътрешността). Според Павлов рефлексите се делят на условни (разработени през живота, специфични за всеки индивид) и безусловни (вродени, видово специфични: хранителни, полови, отбранително-двигателни, хомеостатични и др.).
Как работи рефлексната дъга?
Независимо от вида на рефлекса, неговата рефлексна дъга съдържа рецептор, аферентен път, нервен център, еферентен път, работен орган и обратна връзка. Изключение правят аксоновите рефлекси, чиято структура на рефлексната дъга е разположена в рамките на един неврон: сензорните процеси генерират центростремителни импулси, които, преминавайки през тялото на неврона, се разпространяват по аксона към централната нервна система и по протежение на клона на аксона импулсите достигат до ефектора. Такива рефлекси се приписват на функционирането на метасимпатиковата нервна система, чрез тях например се осъществяват механизми за регулиране на съдовия тонус и активността на кожните жлези.
Функцията за възприемане на дразнене и превръщането му в енергия на възбуждане се изпълнява от рецептори на рефлексни дъги. Рецепторната енергия на възбуждане има характер на локален отговор, който е важен при градирането на възбуждането по сила.
Въз основа на структурата и произхода на рецепторите те могат да бъдат разделени на първични сетивни, вторични сетивни и свободни нервни окончания. При първите самият неврон действа като рецептор (развива се от невроепителия); няма междинни структури между стимула и първия аферентен неврон. Локалният отговор на първичните сензорни рецептори - рецепторният потенциал - също е генераторен потенциал, т.е. индуциране на потенциал за действие през мембраната на аферентното влакно. Първичните сензорни рецептори включват зрителни, обонятелни, хемо- и барорецептори. на сърдечно-съдовата система.
Вторичните чувствителни клетки са специални структури от ненервен произход, които взаимодействат с дендритите на псевдо-униполярни сензорни клетки с помощта на синаптични неврорецепторни контакти. Рецепторният потенциал, възникващ под действието на стимул във вторично чувствителните клетки, не е генератор и не предизвиква появата на потенциал на действие върху мембраната на аферентното влакно. Възбуждащият постсинаптичен потенциал възниква само чрез механизма на освобождаване на медиатора от рецепторната клетка. Градацията на силата на стимула се осъществява чрез отделянето на различни количества от медиатора (колкото повече медиаторът се освобождава, толкова по-силен е стимулът).
Вторичните сензорни клетки включват слухови, вестибуларни, каротидни, тактилни и други рецептори. Понякога, поради особеностите на функциониране, тази група включва фоторецептори, които от анатомична гледна точка и поради произхода си от невроепителия са вторично чувствителни.
Свободните нервни окончания са разклонени дендрити на псевдоуниполярни сензорни клетки и са локализирани в почти всички тъкани на човешкото тяло.
Според енергийната природа на стимула, на който рецепторът реагира, те се разделят на механорецептори (тактилни, барорецептори, волуморецептори, слухови, вестибуларни; те, като правило, възприемат механично дразнене с помощта на клетъчни израстъци), хеморецептори (обонятелни), хеморецептори на кръвоносните съдове, централната нервна система, фоторецептори (възприемат дразнене чрез пръчковидни и конусовидни изходи). редове на клетката), терморецептори (реагират на промяната "топлина-студ" - телца на Руфини и колби на Краузе на лигавиците) и ноцицептори (некапсулирани болкови окончания).
Пострецепторни образувания на рефлексната дъга
Пострецепторното образуване на структурата на рефлексните дъги е аферентен път, образуван от псевдо-униполярен сензорен неврон, чието тяло лежи в спинален ганглий, и се образуват аксони задни корени гръбначен мозък. Функцията на аферентния път е да провежда информация към централната връзка, освен това на този етап информацията се кодира. За тези цели в тялото на гръбначните животни се използва двоичен код, съставен от изблици (залпове) импулси и празнини между тях. Има два основни вида кодиране: честотно и пространствено.
Първият е образуването на различен брой импулси в залп, различен брой залпове, тяхната продължителност и продължителността на паузите между тях в зависимост от силата на стимулацията, приложена към рецептора. Пространственото кодиране осъществява градацията на силата на стимула, като включва различен брой нервни влакна, по които едновременно се извършва възбуждане.
Аферентният път се състои главно от A-α, A-β и A-δ влакна.
Преминавайки през влакната, нервният импулс навлиза в рефлексния център, който в анатомичен смисъл е съвкупност от неврони, разположени на определено ниво на централната нервна система и участващи в образуването на този рефлекс. Функцията на рефлексния център е да анализира и синтезира информация, както и да превключва информация от аферентния към еферентния път.
Функции на рефлексната дъга
В зависимост от отдела на нервната система (соматична и вегетативна), рефлексите, чийто център е разположен в гръбначния мозък, се различават по локализацията на интеркаларните неврони. И така, за соматичната нервна система рефлексният център се намира в междинната зона между предните и задните рога на гръбначния мозък. Рефлексният център на автономната нервна система (тялото на интеркаларните неврони) се намира в задни рога. Соматичните и автономните части на нервната система също се различават по локализацията на еферентните неврони. Телата на двигателните неврони на соматичната нервна система лежат в предните рога на гръбначния мозък, телата на преганглионарните неврони автономна система- на нивото на средните рога.
Аксоните и на двата вида клетки образуват еферентния път на рефлексната дъга. В соматичната нервна система тя е непрекъсната, изградена е от влакна от типа A-α. Единствените изключения са A-γ влакната, които провеждат възбуждане от клетките на гръбначния мозък към интрафузалните влакна на мускулните вретена. Еферентният път на автономната нервна система е прекъснат в автономен ганглийразположени или интрамурални (двойка симпатична част), или близо до гръбначния мозък (самостоятелно или в симпатичен ствол- симпатична част). Преганглионарното влакно принадлежи към В-влакната, постганглионарното влакно към С групата.
Работният орган за соматичната част на нервната система е набраздения скелетен мускул, във вегетативната дъга ефекторът е жлеза или мускул (гладък или набразден сърдечен). Между еферентен пъта работният орган е химически мионеврален или невросекреторен синапс.
Рефлексната дъга се затваря в пръстен поради обратна аферентация - потокът от импулси от ефекторните рецептори обратно към рефлексния център. Функция за обратна връзка - сигнализиране на централната нервна система за извършеното действие. Ако не се изпълнява достатъчно, нервният център се възбужда - рефлексът продължава. Също така, поради обратната аферентация, се осъществява контролът на периферната активност на централната нервна система.
Правете разлика между отрицателна и положителна обратна връзка. Първият, когато изпълнява определена функция, стартира механизъм, който инхибира тази функция. Положителната обратна връзка се състои от допълнително стимулиране на функция, която вече се изпълнява, или инхибиране на функция, която вече е потисната. Положителната обратна аферентация е рядка, тъй като поставя биологичната система в нестабилна позиция.
Простите (моносинаптични) рефлексни дъги се състоят само от два неврона (аферентни и еферентни) и се различават само в проприоцептивните рефлекси. Останалите дъги включват всички горепосочени компоненти.
Рефлексната дъга е верига от неврони от периферен рецептор през централната нервна система до периферен ефектор. Елементите на рефлексната дъга са периферен рецептор, аферентен път, един или повече интерневрони, еферентен път и ефектор.
Всички рецептори участват в определени рефлекси, така че техните аферентни влакна служат като аферентен път на съответната рефлексна дъга. Броят на интерневроните винаги е по-голям от един, с изключение на моносинаптичния рефлекс на разтягане. Еферентният път е представен или от моторни аксони, или от постганглионарни влакна на автономната нервна система, а ефекторите са скелетните мускули и гладките мускули, сърцето и жлезите.
Времето от началото на стимула до отговора на ефектора се нарича рефлексно време. В повечето случаи се определя главно от времето на провеждане в аферентните и еферентните пътища и в централната част на рефлексната дъга, към което трябва да се добави времето на трансформация на стимула в рецептора в разпространяващ се импулс, времето на предаване през синапсите в централната нервна система (синаптично забавяне), времето на предаване от еферентния път към ефектора и времето на активиране на ефектора.
Рефлексните дъги са разделени на няколко вида
1. Моносинаптични рефлексни дъги - в такава дъга участва само един синапс, разположен в централната нервна система. Такива рефлекси са много чести при всички гръбначни животни, те участват в регулацията мускулен тонуси поза (напр. рефлекс в коляното). В тези дъги невроните не достигат до мозъка и рефлексните действия се извършват без негово участие, тъй като те са стереотипни и не изискват мисъл или съзнателно решение. Те са икономични по отношение на броя на участващите централни неврони и не изискват намесата на мозъка.
2. Полисинаптични спинални рефлексни дъги - включват най-малко два синапса, разположени в централната нервна система, тъй като в дъгата е включен трети неврон - интеркаларен, или междинен неврон. Тук има синапси между сетивния неврон и интерневрона и между интеркаларните и моторните неврони. Такива рефлексни дъги позволяват на тялото да извършва автоматични неволни реакции, необходими за адаптиране към промените във външната среда (напр. зеничен рефлексили поддържане на равновесие при движение) и до промени в самото тяло (регулиране на дихателната честота, кръвното налягане и др.).
3. Полисинаптични рефлексни дъги, включващи както гръбначния, така и главния мозък – при този тип рефлексни дъги има синапс в гръбначния мозък между сензорния неврон и неврона, който изпраща импулси към мозъка.
Рефлексите могат да бъдат класифицирани според различни критерии. Така че, в зависимост от нивото на затваряне на дъгата, т.е. според местоположението на рефлексния център рефлексите се делят на спинални (рефлексът се затваря в гръбначния мозък), булбарни (рефлексен център е продълговатия мозък), мезенцефални (рефлексната дъга е затворена в средния мозък), диенцефални и кортикални рефлексни центрове се намират съответно в теленцефалона и кората на главния мозък.
Според ефекторния признак те са соматични, когато еферентният път на рефлекса осигурява двигателна инервация на скелетните мускули, и вегетативни, когато вътрешните органи са ефектори.
В зависимост от вида на раздразнените рецептори, рефлексите се разделят на екстероцептивни (ако рецепторът възприема информация от външната среда), проприоцептивни (рефлексната дъга започва от рецепторите на опорно-двигателния апарат) и интероцептивни (от рецепторите на вътрешните органи).
Интероцептивните рефлекси от своя страна се делят на висцеро-висцерални (рефлексна дъга свързва два вътрешни органа), висцеро-мускулни (рецепторите са разположени върху мускулно-сухожилния апарат, ефекторът е вътрешен орган) и висцеро-кутанни (рецепторите са локализирани в кожата, работните органи са вътрешността).
Според Павлов рефлексите се делят на условни (разработени през живота, специфични за всеки индивид) и безусловни (вродени, видово специфични: хранителни, полови, отбранително-двигателни, хомеостатични и др.).
Независимо от вида на рефлекса, неговата рефлексна дъга съдържа рецептор, аферентен път, нервен център, еферентен път, работен орган и обратна връзка. Изключение правят аксоновите рефлекси, чиято рефлексна дъга се намира в рамките на един неврон: сензорните процеси генерират центростремителни импулси, които, преминавайки през тялото на неврона, се разпространяват по аксона към централната нервна система и по клона на аксона импулсите достигат до ефектора. Такива рефлекси се приписват на функционирането на метасимпатиковата нервна система, чрез тях например се осъществяват механизми за регулиране на съдовия тонус и активността на кожните жлези.
Функцията за възприемане на дразнене и превръщането му в енергия на възбуждане се изпълнява от рецептори на рефлексни дъги. Рецепторната енергия на възбуждане има характер на локален отговор, който е важен при градирането на възбуждането по сила.
Въз основа на структурата и произхода на рецепторите те могат да бъдат разделени на първични сетивни, вторични сетивни и свободни нервни окончания. При първите самият неврон действа като рецептор (развива се от невроепителия); няма междинни структури между стимула и първия аферентен неврон. Локалният отговор на първичните сензорни рецептори - рецепторният потенциал - също е генераторен потенциал, т.е. индуциране на потенциал за действие през мембраната на аферентното влакно. Първичните сензорни рецептори включват зрителни, обонятелни, хемо- и барорецептори на сърдечно-съдовата система.
Вторичните чувствителни клетки са специални структури от ненервен произход, които взаимодействат с дендритите на псевдо-униполярни сензорни клетки с помощта на синаптични неврорецепторни контакти. Рецепторният потенциал, възникващ под действието на стимул във вторично чувствителни клетки, не е генератор и не предизвиква появата на потенциал на действие върху мембраната на аферентното влакно. Възбуждащият постсинаптичен потенциал възниква само чрез механизма на освобождаване на медиатора от рецепторната клетка. Градацията на силата на стимула се осъществява чрез отделянето на различни количества от медиатора (колкото повече медиаторът се освобождава, толкова по-силен е стимулът).
Вторичните сензорни клетки включват слухови, вестибуларни, каротидни, тактилни и други рецептори. Понякога, поради особеностите на функциониране, тази група включва фоторецептори, които от анатомична гледна точка и поради произхода си от невроепителия са вторично чувствителни.
Свободните нервни окончания са разклонени дендрити на псевдоуниполярни сензорни клетки и са локализирани в почти всички тъкани на човешкото тяло.
Според енергийния характер на стимула, на който рецепторът реагира, те се разделят на механорецептори (тактилни, барорецептори, волуморецептори, слухови, вестибуларни; те, като правило, възприемат механично дразнене с помощта на клетъчни израстъци), хеморецептори (обонятелни), съдови хеморецептори, централната нервна система, фоторецептори (възприемат дразнене чрез пръчковидни и конусовидни изходи). редове на клетката), терморецептори (реагират на промяната "топлина-студ" - телца на Руфини и колби на Краузе на лигавиците) и ноцицептори (некапсулирани болкови окончания).
Пострецепторното образуване на рефлексни дъги е аферентен път, образуван от псевдо-униполярен сензорен неврон, чието тяло лежи в гръбначния ганглий, а аксоните образуват задните корени на гръбначния мозък. Функцията на аферентния път е да провежда информация към централната връзка, освен това на този етап информацията се кодира. За тези цели в тялото на гръбначните животни се използва двоичен код, съставен от изблици (залпове) импулси и празнини между тях. Има два основни вида кодиране: честотно и пространствено.
Първият е образуването на различен брой импулси в залп, различен брой залпове, тяхната продължителност и продължителността на паузите между тях в зависимост от силата на стимулацията, приложена към рецептора. Пространственото кодиране осъществява градацията на силата на стимула, като включва различен брой нервни влакна, по които едновременно се извършва възбуждане.
В състава на аферентния път влизат предимно А-Ь, А-с и А-е влакна.
Преминавайки през влакната, нервният импулс навлиза в рефлексния център, който в анатомичен смисъл е съвкупност от неврони, разположени на определено ниво на централната нервна система и участващи в образуването на този рефлекс. Функцията на рефлексния център е да анализира и синтезира информация, както и да превключва информация от аферентния към еферентния път.
В зависимост от отдела на нервната система (соматична и вегетативна), рефлексите, чийто център е разположен в гръбначния мозък, се различават по локализацията на интеркаларните неврони. И така, за соматичната нервна система рефлексният център се намира в междинната зона между предните и задните рога на гръбначния мозък. Рефлексният център на автономната нервна система (телата на интеркаларните неврони) се намира в задните рога. Соматичните и автономните части на нервната система също се различават по локализацията на еферентните неврони. Телата на моторните неврони на соматичната нервна система лежат в предните рога на гръбначния мозък, телата на преганглионарните неврони на автономната система лежат на нивото на средните рога.
Аксоните и на двата вида клетки образуват еферентния път на рефлексната дъга. В соматичната нервна система тя е непрекъсната, изградена е от влакна тип A-b. Единствените изключения са A-g влакна, които провеждат възбуждане от клетките на гръбначния мозък към интрафузалните влакна на мускулните вретена. Еферентният път на автономната нервна система се прекъсва във вегетативния ганглий, разположен или интрамурално (парасимпатиковата част), или близо до гръбначния мозък (отделно или в симпатиковия ствол - симпатиковата част). Преганглионарното влакно принадлежи към В-влакната, постганглионарното влакно принадлежи към С групата.
Работният орган за соматичната част на нервната система е напречно набразден скелетен мускул, във вегетативната дъга ефекторът е жлеза или мускул (гладък или набразден сърдечен). Между еферентния път и работния орган има химичен мионеврален или невросекреторен синапс.
Рефлексната дъга се затваря в пръстен поради обратна аферентация - потокът от импулси от ефекторните рецептори обратно към рефлексния център. Функция за обратна връзка - сигнализиране на централната нервна система за извършеното действие. Ако не се изпълнява достатъчно, нервният център се възбужда - рефлексът продължава. Също така, поради обратната аферентация, се осъществява контролът на периферната активност на централната нервна система.
Правете разлика между отрицателна и положителна обратна връзка. Първият, когато изпълнява определена функция, стартира механизъм, който инхибира тази функция. Положителната обратна връзка се състои от допълнително стимулиране на функция, която вече се изпълнява, или инхибиране на функция, която вече е потисната. Положителната обратна аферентация е рядка, тъй като поставя биологичната система в нестабилна позиция.
Простите (моносинаптични) рефлексни дъги се състоят само от два неврона (аферентни и еферентни) и се различават само в проприоцептивните рефлекси. Останалите дъги включват всички горепосочени компоненти.
Физиологични свойства и функционално значение на нервните влакна
Нервните влакна имат най-висока възбудимост, най-висока скорост на провеждане на възбуждане, най-кратък рефрактерен период и висока лабилност. Това е предвидено високо нивометаболитни процеси и нисък мембранен потенциал.
Функция: провеждане на нервните импулси от рецепторите към централната нервна система и обратно.
Структурни особености и видове нервни влакна
Нервното влакно - аксонът - е покрито с клетъчна мембрана.
Има 2 вида нервни влакна:
Немиелинизирани нервни влакна - един слой Шванови клетки, между тях - цепнатини. клетъчната мембранапрез целия контакт с заобикаляща среда. При прилагане на дразнене възниква възбуждане на мястото на действие на стимула. Немиелинизираните нервни влакна имат електрогенни свойства (способността да генерират нервни импулси) навсякъде.
Миелинизирани нервни влакна – покрити със слоеве от Шванови клетки, които на места образуват възли на Ранвие (участъци без миелин) през 1 мм. Продължителността на прихващане на Ranvier е 1 µm. Миелиновата обвивка изпълнява трофични и изолационни функции (висока устойчивост). Зоните, покрити с миелин, нямат електрогенни свойства. Имат пасиранията на Ранвие. Възбуждането възниква при прихващане на Ranvier най-близо до мястото на действие на стимула. При засечките на Ранвие висока плътност Na-каналите, следователно, при всяко прихващане на Ranvier се получава увеличение на нервните импулси.
Прихващанията на Ranvier действат като повторители (генерират и усилват нервните импулси).
Механизмът на провеждане на възбуждане по нервните влакна
1885 - Л. Герман - възникват кръгови токове между възбудените и невъзбудените участъци на нервното влакно.
Под действието на дразнителя възниква потенциална разлика между външната и вътрешната повърхност на тъканта (области, които носят различни заряди). Между тези зони възниква електрически ток (движението на Na + йони). Вътре в нервното влакно възниква ток от положителния полюс към отрицателния полюс, т.е. токът е насочен от възбудената зона към невъзбудената. Този ток излиза през невъзбудената област и я кара да се презареди. По външната повърхност на нервното влакно токът протича от невъзбудената зона към възбудената област. Този ток не променя състоянието на възбудената зона, тъй като тя е в състояние на рефрактерност.
Доказателство за наличието на кръгови течения: нервното влакно е поставено в разтвор на NaClи регистрирайте скоростта на възбуждане. След това нервното влакно се поставя в масло (съпротивлението се увеличава) - скоростта на проводимост намалява с 30%. След това нервното влакно се оставя във въздуха - скоростта на възбуждане се намалява с 50%.
Характеристики на провеждането на възбуждане по протежение на миелинизирани и немиелинизирани нервни влакна:
миелинови влакна - имат обвивка с висока устойчивост, електрогенни свойства само във възлите на Ранвие. Под действието на стимула възниква възбуждане в най-близкия интерсепт на Ранвие. Прихващане на съсед в състояние на поляризация. Полученият ток причинява деполяризация на съседния интерсепт. Възлите на Ranvier имат висока плътност на Na-канали, следователно във всеки следващ възел възниква малко по-голям (по амплитуда) потенциал на действие, поради което възбуждането се разпространява без намаляване и може да прескочи няколко възела. Това е солидната теория на Тасаки. Доказателството на теорията е, че в нервното влакно са инжектирани лекарства, които блокират няколко прихващания, но след това е записано провеждането на възбуждане. Това е много надежден и печеливш метод, тъй като се елиминират незначителни щети, увеличава се скоростта на възбуждане и се намаляват разходите за енергия;
немиелинизирани влакна - повърхността има електрогенни свойства навсякъде. Следователно малки кръгови токове възникват на разстояние от няколко микрометра. Възбуждането има формата на постоянно движеща се вълна.
Този метод е по-малко печеливш: високи разходи за енергия (за работата на Na-K помпата), по-ниска скорост на възбуждане.
Класификация на нервните влакна
Нервните влакна се класифицират според:
продължителността на потенциала на действие;
структура (диаметър) на влакното;
скорост на възбуждане.
Различават се следните групи нервни влакна:
група А (алфа, бета, гама, делта) - най-късият потенциал на действие, най-дебелата миелинова обвивка, най-високата скорост на възбуждане;
група В - миелиновата обвивка е по-слабо изразена;
Група С - без миелинова обвивка.
Морфологични разлики между дендрити и аксони
1. Отделният неврон има няколко дендрита, аксонът винаги е един.
2. Дендритите винаги са по-къси от аксона. Ако размерът на дендритите не надвишава 1,5-2 mm, тогава аксоните могат да достигнат 1 m или повече.
3. Дендритите плавно се отдалечават от тялото на клетката и постепенно имат постоянен диаметър на значително разстояние.
4. Дендритите обикновено се разклоняват под остър ъгъл, а клоните са насочени встрани от клетката. Аксоните отделят колатерали най-често под прав ъгъл; ориентацията на колатералите не е пряко свързана с позицията на клетъчното тяло.
5. Моделът на дендритно разклоняване в клетки от същия тип е по-постоянен от разклоняването на аксона на тези клетки.
6. Дендритите на зрелите неврони са покрити с дендритни шипове, които отсъстват върху сомата и началната част на дендритните стволове. Аксоните нямат шипове.
7. Дендритите никога нямат месеста обвивка. Аксоните често са заобиколени от миелин.
8. Дендритите имат по-правилна пространствена организация на микротубулите, аксоните са доминирани от неврофиламенти и микротубулите са по-малко подредени
9. В дендритите, особено в техните проксимални части, има ендоплазмен ретикулум и рибозоми, които не са в аксоните.
10. Повърхността на дендритите в повечето случаи е в контакт със синоптични плаки и има активни зони с постсинаптична специализация.
Структурата на дендритите
Ако има сравнително голяма литература за геометрията на дендритите, дължината на техните разклонения, ориентацията, тогава около вътрешна структура, за структурата на отделните компоненти на цитоплазмата им има само отделни разпръснати сведения. Тази информация стана възможна едва с въвеждането на електронномикроскопските изследвания в неврохистологията.
Основните характерни черти на дендрита, които го отличават на електронномикроскопски срезове:
1) липса на миелинова обвивка,
наличието на правилна система от микротубули,
3) наличието на активни зони на синапси върху тях с ясно изразена електронна плътност на цитоплазмата на дендрита,
4) отклонение от общия ствол на дендрита на шиповете,
5) специално организирани зони на разклонителни възли,
6) включване на рибозоми,
7) наличието на гранулиран и негранулиран ендоплазмен ретикулум в проксималните области.
Най-забележителната характеристика на дендритната цитоплазма е наличието на множество микротубули. Те са добре идентифицирани както в напречни, така и в надлъжни разрези. Започвайки от проксималния участък на дендрита, микротубулите вървят успоредно на дългата ос на дендрита до дисталните му клонове. Микротубулите следват в дендрита успоредно един на друг, без да се свързват или пресичат един с друг. В напречните сечения се вижда, че разстоянията между отделните тубули са постоянни. Индивидуалните дендритни тубули се простират на доста големи разстояния, често следвайки криви, които могат да бъдат по протежение на хода на дендритите. Броят на тубулите е относително постоянен на единица площ от напречното сечение на дендрита и е приблизително 100 на 1 µm. Това число е типично за всички дендрити, взети от различни отделицентрална и периферна нервна система, различни видовеживотни.
Функцията на микротубулите е транспортирането на вещества по процесите на нервните клетки.
Когато микротубулите са унищожени, транспортът на вещества в дендрита може да бъде нарушен и по този начин крайните участъци на процесите са лишени от притока на хранителни вещества и енергийни веществаот тялото на клетката. Дендритите, за да поддържат структурата на синаптичните контакти при екстремни условия и по този начин да осигурят функцията на междуневронно взаимодействие, компенсират дефицита хранителни веществапоради съседни на тях структури (синаптични плаки, многослойна миелинова обвивка от меки влакна, както и фрагменти от глиални клетки).
Ако действието на патогенния фактор се елиминира своевременно, дендритите възстановяват структурата и правилната пространствена организация на микротубулите, като по този начин възстановяват системата за транспортиране на вещества, която е присъща на нормалния мозък. Ако силата и продължителността на патогенния фактор са значителни, тогава явленията на ендоцитозата, вместо тяхната адаптивна функция, могат да станат фатални за дендритите, тъй като фагоцитираните фрагменти не могат да бъдат използвани и, натрупвайки се в цитоплазмата на дендритите, ще доведат до нейното необратимо увреждане.
Нарушаването на организацията на микротубулите води до рязка промяна в поведението на животните. При животни, при които в експеримента са унищожени микротубулите в дендритите, се наблюдава дезорганизация на сложни форми на поведение, докато простите условни рефлекси са запазени. При хората това може да доведе до сериозни смущения във висшата нервна дейност.
Фактът, че дендритите са най-чувствителното място към действието на патологичен агент в психично заболяване, свидетелстват някои трудове на американски учени. Оказа се, че при сенилна деменция (цианотична деменция) и болестта на Алцхаймер мозъчните препарати, обработени по метода на Голджи, не разкриват процесите на нервните клетки. Стволовете на дендритите изглеждат изгорени и овъглени. Неоткриването на тези процеси в хистологичните препарати на мозъка вероятно също е свързано с нарушение на системата от микротубули и неврофиламенти в тези процеси.
Намира се в дендритите. Те следват успоредно на дългата ос на дендрита, могат да лежат отделно или да бъдат събрани в снопове, но не са строго разположени в цитоплазмата. Вероятно, заедно с микротубулите, те могат да бъдат еквивалент на неврофибрили.
Всички дендрити на ЦНС се характеризират с увеличаване на повърхността, дължащо се на множество дихотомични разделения. В този случай в зоните на разделяне се формират специални места за разширяване или разклонителни възли.
Нормалният анализ показва, че в разклонителния възел, към който се приближават два дендритни клона, всеки от които носи свой собствен сигнал, могат да бъдат извършени следните операции. Чрез разклонения възел общ багажники по-нататък към тялото на невронния проход:
или сигнал от един клон,
или просто от друг
или резултат от взаимодействието на два сигнала,
или сигналите взаимно се компенсират.
Цитоплазмата на разклонения възел съдържа почти всички компоненти, които са характерни за тялото нервна клетка, а местата рязко се различават по своята структура от цитоплазмата на общия дендритен ствол и разклонения, получени чрез разделяне. Разклонените възли съдържат увеличен брой митохондрии, гранулиран и гладък ретикулум, видими са клъстери от единични рибозоми и рибозоми, събрани в розетки. Тези компоненти (гранулиран и гладък ретикулум, рибозоми) участват пряко в синтеза на протеини. Натрупването на митохондрии на тези места показва интензивността на окислителните процеси.
Функции на дендритите
Бих искал да отбележа, че основните трудности, които изследователят среща при изучаването на функцията на дендритите, е липсата на информация за свойствата на дендритната мембрана (за разлика от мембраната на тялото на неврона) поради невъзможността за въвеждане на микроелектрод в дендрита.
Оценявайки общата геометрия на дендритите, разпределението на синапсите и специалната структура на цитоплазмата в местата на разклоняване на дендритите, може да се говори за специални невронни локуси със собствена функция. Най-простото нещо, което може да се припише на дендритните места в местата на разклоняване, е трофична функция.
От гореизложеното следва, че цитоплазмата на дендритите съдържа много ултра структурни компонентиспособни да изпълняват своите важни функции. В дендрита има определени локуси, където работата му има свои собствени характеристики.
Основната цел на многобройните дендритни клонове на нервната клетка е да осигурят взаимовръзка с други неврони. в мозъчната кора на бозайниците голям дялаксодендричните връзки попадат върху контакти със специални специализирани израстъци на дендрити - дендритни шипове. Дендритните шипове са филогенетично най-младите образувания в нервната система. В онтогенезата те узряват много по-късно от другите нервни структури и представляват най-пластичния апарат на нервната клетка.
По правило дендритният шип има характерна форма в мозъчната кора на бозайниците. (фиг. 2). От основния дендритен ствол се отклонява сравнително тясна дръжка, която завършва с разширение - главата. Вероятно тази форма на дендритния придатък (наличието на глава) е свързана, от една страна, с увеличаване на площта на синаптичен контакт с края на аксона, а от друга страна, тя служи за настаняване на специализирани органели вътре в гръбначния стълб, по-специално, бодливият апарат, който присъства само в дендритните шипове на мозъчната кора на бозайниците. В това отношение изглежда подходяща аналогия с формата на края на синаптичния аксон, когато тънко претерминално влакно образува разширение. Това разширение (синаптична плака) образува обширен контакт с инервирания субстрат и съдържа вътре голям набор от ултраструктурни компоненти (синаптични везикули, митохондрии, неврофиламенти, гликогенови гранули).
Съществува хипотеза (която в частност се споделя и развива от Нобеловия лауреат Ф. Крик), че геометрията на шиповете може да се променя в зависимост от функционалното състояние на мозъка. В този случай тясната шийка на гръбначния стълб може да се разшири, а самият гръбначен стълб се изравнява, което води до повишаване на ефективността на аксо-гръбначния контакт.
Ако формата и размерът на дендритните шипове в мозъчната кора на бозайниците могат да варират до известна степен, тогава най-постоянното в тях е наличието на специфичен гръбначен апарат. Това е комплекс от взаимосвързани тубули (цистерни), разположени, като правило, в главата на гръбначния стълб. Вероятно тази органела е свързана с много важни функции, присъщи на филогенетично най-младите мозъчни образувания, тъй като бодливият апарат се намира главно в мозъчната кора и само при висшите животни.
Въпреки всичко шипът е производно на дендрита, в него липсват неврофиламенти и дендритни тубули, цитоплазмата му съдържа едро или фино гранулиран матрикс. Още едно особеностгръбначен стълб в мозъчната кора е задължителното наличие на синаптични контакти с аксонни окончания върху тях. Цитоплазмата на гръбнака има специални компоненти, които я отличават от дендритните стъбла. В цитоплазмата на гръбначния стълб може да се отбележи особена триада: субсинаптична специализация на активните зони - бодлив апарат - митохондрии. Като се има предвид разнообразието от сложни и важни функции, изпълнявани от митохондриите, могат да се очакват и сложни функционални прояви в "триади" по време на синаптично предаване. Може да се каже, че цитоплазмата на дендритния бодил и спиналният апарат могат да бъдат пряко свързани със синаптичната функция.
Дендритните шипове и краищата на дендритите също са много чувствителни към екстремни фактори. При всякакъв вид отравяне (например алкохолно, хипоксично, тежки метали - олово, живак и др.), Броят на шиповете, намиращи се върху дендритите на клетките на мозъчната кора, се променя. По всяка вероятност бодлите не изчезват, но техните цитоплазмени компоненти са нарушени и те са по-лошо импрегнирани със соли. тежки метали. Тъй като шиповете са един от структурните компоненти на междуневронните контакти, неизправностите в тях водят до сериозно увреждане на мозъчната функция.
В някои случаи, при краткотрайно действие на екстремен фактор, на пръв поглед може да възникне парадорзална ситуация, когато броят на шиповете, открити върху дендритите на мозъчните клетки, не намалява, а се увеличава. И така, това се наблюдава при експериментална церебрална исхемия в началния й период. Успоредно с увеличаването на броя на идентифицираните шипове, функционално състояниемозък. В този случай хипоксията е фактор, който допринася за повишен метаболизъм в нервната тъкан, по-добро използване на резервите, които не се използват в нормална среда, и бързото изгаряне на токсините, натрупани в тялото. Ултраструктурно това се проявява в по-интензивно изследване на цитоплазмата на шиповете, растеж и разширяване на цистерните на гръбначния апарат. Вероятно този феномен на положителния ефект на хипоксията се наблюдава, когато човек, изпитващ голямо физически упражненияв условия на хипоксия, покорява планински върхове. След това тези затруднения се компенсират от по-интензивна продуктивна работа, както на мозъка, така и на други органи.
Образуване на дендрити
Дендритите и техните междуневронни връзки се образуват по време на онтогенетичното развитие на мозъка. Освен това дендритите, по-специално апикалните, при млади индивиди остават свободни за известно време, за да образуват нови контакти. Частите на дендрита, разположени по-близо до тялото на клетката, вероятно са свързани с по-силни и прости естествени условни рефлекси, а краищата са оставени за образуване на нови връзки и асоциации.
В зряла възраст върху дендритите вече няма зони, свободни от междуневронни контакти, но с напредването на възрастта първо страдат краищата на дендритите и по отношение на насищането с контакти
при старите индивиди те приличат на дендрити детство. Това се случва както поради отслабване на процесите на синтез на транспортни протеини в клетката, така и поради нарушено кръвоснабдяване на мозъка. Може би именно тук лежи морфологичната основа за така добре познатата в неврологията и ин ежедневиетофактът, че когато на възрастните хора им е трудно да научат нещо ново, те често забравят текущите събития и помнят много добре миналото. Същото се наблюдава и при отравяне.
Както вече беше отбелязано, увеличаването и усложняването на дендритното дърво във филогенезата е необходимо не само за възприятието Голям бройвходящи импулси, но и за предварителна обработка.
Дендритите на невроните на централната нервна система имат синаптична функция навсякъде и крайните участъци по никакъв начин не са по-ниски от средните в това. Ако говорим сиоколо дисталните (терминални) участъци на апикалните дендрити пирамидални невронимозъчната кора, тогава техният дял в осъществяването на междуневронни взаимодействия е дори по-значим от проксималните. Там до Повече ▼крайните синаптични плаки върху самия ствол и върху клоните на апикалния дендрит се съединяват от повече контакти върху дендритни шипове.
Изучавайки този проблем с помощта на електронна микроскопия, изследователите също се убедиха, че крайните участъци на дендритите са плътно покрити със синаптични плаки и по този начин са пряко включени в междуневронните взаимодействия. електронна микроскопиясъщо показа, че дендритите могат да образуват контакти един с друг. Тези контакти могат да бъдат или паралелни, на които повечето автори приписват електротонични свойства, или типични асиметрични синапси с добре дефинирани органели, които осигуряват химическо предаване. Такива дендро-дендритни контакти едва започват да привличат вниманието на изследователите. И така, дендритът по цялата си дължина изпълнява синаптична функция. Как повърхността на дендрита е адаптирана да осигури контакт с окончанията на аксоните?
Повърхностната мембрана на дендрита е предназначена да се използва максимално за междуневронни контакти. Целият дендрит е осеян с вдлъбнатини, гънки, джобове, има различни неравности от вида на микроизрастъци, шипове, подобни на гъби придатъци и др. Всички тези релефи на дендритните стволове съответстват на формата и размера на входящите синаптични окончания. И в различни отделинервна система и при различните животни релефът на дендритната повърхност има специфични особености. Разбира се, най-забележителният израстък на дендритната мембрана е дендритният шип.
Дендритите са много чувствителни към действието на различни екстремни фактори. Нарушенията в тях водят до много заболявания, като например психични разстройства.
Рефлексната дъга се състои от:
- рецептори - възприемащи дразнене.
- чувствително (центростремително, аферентно) нервно влакно, което предава възбуждане към центъра
- нервният център, където се извършва превключването на възбуждането от сетивните неврони към моторните неврони
- двигателно (центробежно, еферентно) нервно влакно, което пренася възбуждане от централната нервна система към работния орган
- ефектор - работен орган, който извършва въздействие, реакция в отговор на дразнене на рецептора.
Рецептори и рецептивни полета
Рецептор- клетка, възприемаща дразнене.
рецептивно поле- това е анатомичната област, при дразнене се предизвиква този рефлекс.
Рецептивните полета на първичните сензорни рецептори са организирани по най-прост начин. Например, тактилното или ноцицептивното рецептивно поле на повърхността на кожата е разклонение на едно сетивно влакно.
Рецепторите, разположени в различни части на рецептивното поле, имат различна чувствителност към адекватна стимулация. Зоната с висока чувствителност обикновено се намира в центъра на рецептивното поле и чувствителността намалява по-близо до периферията на рецептивното поле.
Рецептивните полета на вторичните сензорни рецептори са организирани по подобен начин. Разликата е, че клоновете на аферентното влакно не завършват свободно, а имат синаптични контакти с чувствителни рецепторни клетки. По този начин са организирани вкусовите, вестибуларните, акустичните рецептивни полета.
припокриващи се рецептивни полета. Същата област на чувствителната повърхност (например кожа или ретина) се инервира от няколко чувствителни нервни влакна, които с разклоненията си припокриват рецептивните полета на отделните аферентни нерви.
Поради припокриването на рецептивните полета общата сензорна повърхност на тялото се увеличава.
Класификация на рефлексите.
По вид образование:
Условно (придобито) - отговаря на името, слюнка от кучето в светлината.
Безусловно (вродено) - мигащо преглъщане, коляно.
По местоположение рецептори:
Екстероцептивни (кожни, зрителни, слухови, обонятелни)
Интероцептивни (от рецепторите на вътрешните органи)
Проприоцептивни (от рецептори в мускули, сухожилия, стави)
За ефектори:
Соматични или двигателни (рефлекси скелетни мускули);
Вегетативни вътрешни органи - храносмилателни, сърдечно-съдови, отделителни, отделителни и др.
По биологичен произход:
Отбранителна или защитна (отговор на разделяне на тактилна болка)
Храносмилателни (дразнещи рецептори в устната кухина.)
Сексуални (хормони в кръвта)
Приблизително (завъртане на главата, тялото)
Мотор
Позотонични (поддържащи пози на тялото)
По броя на синапсите:
Моносинаптични, чиито дъги се състоят от аферентни и еферентни неврони (например коляно).
Полисинаптични, чиито дъги също съдържат 1 или повече междинни неврони и имат 2 или повече синаптични превключвателя. (соматични и вегетативни справки).
Дисинаптичен (2 синапса, 3 неврона).
По естеството на отговора:
Мотор \ двигател (мускулни контракции)
Секреторна (секреция на секреторни жлези)
Вазомоторни (разширяване и стесняване на кръвоносните съдове)
Сърдечен (промяна. Работата на сърдечния мускул.)
Според продължителността:
фазово (бързо) отдръпване на ръката
тонизиращо (бавно) поддържане на позата
Според местоположението на нервния център:
Спинален (включени са SM неврони) - издърпване на ръката от горещите сегменти 2-4, коляно.
рефлекси в мозъка
Булбар (medulla oblongata) - затваряне на клепачите при допир. към роговицата.
Мезенцефален (среден m) - ориентир за зрението.
Диенцефален (среден мозък) - обоняние
Кортикална (кора BP GM) - условно. реф.
Имоти нервни центрове.
1. Едностранно разпространение на възбуждането.
Възбуждането се предава от аферентния към еферентния неврон (причина: структурата на синапса).
Забавяне на предаването на възбуждане.
Балсам Наличието на много синапси също зависи от силата на дразнителя (сумация) и от физическото състояние. ЦНС (умора).
3.Сумиранесумиране на ефектите, под праговите стимули.
Временен: реф. От пред. Imp-sa още не е минал, но следа. Вече пристигна.
Пространствено: смесване на няколко. Затънтена вода Те са обусловени от сови. Изображения. Реф.
Център за улеснение и оклузия.
Центърът на облекчение - възниква под действието на оптималния стимул (max response) - появи се. Помощен център.
Под действието на min irr. (намалена отв. Рекция) имаше оклузия.
Усвояване и трансформиране на ритъма на възбуждане.
Трансформация - промяна в честотата на нервния импулс при преминаване през нервния център. Честотата може да се увеличи или намали.
Усвояване (танц, ежедневие)
Последица
Забавянето на края на реакцията след спиране на стимула. Свързан с кръвоносния нерв. имп. Затворено Вериги от неврони.
краткосрочно (части от секундата)
дълго (секунди)
Ритмична дейност на нервните центрове.
Увеличаване или намаляване на честотата на нервните импулси, свързани със свойствата на синапса и интегративната продължителност на невроните.
8. Пластичност на нервните центрове.
Способността да се възстанови функционалността на дадено свойство за по-ефективно регулиране на функциите, прилагането на нови рефлекси, които преди това не са били характерни за този център, или възстановяването на функ. В основата на слоя от synps е промяна в мол-та улица.
Промени в възбудимостта под въздействието на химикали.
Висока чувствителност към реалната разлика.
Умора на нервните центрове.
Свързано с висока умора на синапсите. Намалете чувствата. Рецептори.
Общи принципи на координационната дейност на централната нервна система.
спирачна-специален нер. процента изразяваща се в намаляване или пълно изчезване на респ. реакции.
Принцип на конвергенция
Конвергенцията е конвергенцията на импулси, идващи през различни аферентни пътища във всеки един централен неврон или нервен център.
2 . Принципът на конвергенцията е тясно свързан с принципа общ краен пътотвори Sherrinkton. Много различни стимули могат да възбудят един и същ двигателен неврон и една и съща двигателна реакция. Този принцип се дължи на неравния брой аферентни и еферентни пътища.
Принцип на дивергенцията
Това е контактът на един неврон с много други.
Облъчване и концентрация на възбуждане.
Разпространението на процеса на възбуждане към други нервни центрове се нарича облъчване (изборен- в една посока , обобщено- обширен).
След известно време облъчването се заменя с феномена на концентрация на възбуждане в същата начална точка на ЦНС.
Процесът на облъчване играе положителна (образуване на нови условни рефлекси) и отрицателна (нарушаване на фините връзки, които са се развили между процесите на възбуждане и инхибиране, което води до нарушение на двигателната активност) роли.
Принципът на реципрочност (забавя)
Възбуждането на някои клетки предизвиква инхибиране на други чрез интеркаларния неврон.
Доминиращ принцип
Ухтомски формулира принципа на доминантността като принцип на работа на нервните центрове. срок доминантенобозначава доминиращия фокус на възбуждане на централната нервна система, който определя текущата активност на тялото.
Принципи на доминиращия фокус :
Повишена възбудимост на нервните центрове;
Устойчивост на възбуждане на възбуждане във времето;
Способността за сумиране на външни стимули;
Инерция (способност за поддържане на възбуда за дълго време след края на действието на дразнене); способността да предизвиква конюгатни инхибиции.

