Skeleta-muskuļu sistēma kalpo, lai noturētu ķermeni noteiktā stāvoklī un pārvietotos telpā. Skeleta-muskuļu sistēma sastāv no skeleta sistēmas (skeleta), saitēm, locītavām un skeleta muskulis. Kauli, saites un locītavas ir pasīvie kustību orgānu elementi. Muskuļi ir motora sistēmas aktīvā daļa.
Somatomotorās nervu sistēmas evolūcija
Tālāk mēs aplūkosim tikai somatomotoro nervu sistēmu, kas inervē skeleta muskuļus. Animācija: muskuļu kontrakcijas molekulārais mehānisms. Somatomotorā nervu sistēma mainās, pakāpeniski attīstās evolūcijas gaitā. Attīstība notiek paralēli skeleta muskuļu veidošanai un kustību mehānismam, jo struktūras ir atkarīgas viena no otras. Evolūcijas laikā mēs varam runāt par dzīvniekiem tikai tad, ja tie darbojas muskuļu parādās cilvēka ķermenī. Uz triecienu reaģē primitīvi, vienkārši konstruēti dzīvnieki ar tīklveida nervu sistēmu vidi ar spēcīgu kustību to visu muskuļu masa, kuras mērķis bija izvairīties kaitīgās ietekmes vai pieiet pie ēdiena.
Skelets kalpo kā atbalsts un aizsardzība visam ķermenim un atsevišķi ķermeņi, un daudzi kauli ir arī spēcīgas sviras, ar kuru palīdzību tiek veiktas dažādas ķermeņa un tā daļu kustības telpā. Muskuļi iekustina visu kaulu sviru sistēmu. Skelets veido ķermeņa pamatu un nosaka tā izmēru un formu. skeleta daļas, piemēram, galvaskauss, ribu būris un iegurnis mugurkauls, kalpo kā vieta dzīvībai svarīgu orgānu - smadzeņu, plaušu, sirds, zarnu u.c. uzglabāšanai un aizsardzībai. Skelets ir iesaistīts vielmaiņā, jo īpaši, lai uzturētu noteiktu līmeni. minerālu sastāvs asinis. Turklāt vairākas vielas, kas veido kaulus (kalcijs, fosfors, citronskābe utt.), Ja nepieciešams, viegli nonāk vielmaiņas procesos. Kaulu vielas organisko bāzi galvenokārt veido olbaltumvielas, savukārt minerālvielu veido kalcija un fosfora sāļi.
Viņu vienkāršā nervu sistēma un muskuļu sistēma vairs nebija atļauta diferencēta kustība. Evolūcijas gaitā attīstījās arvien perfektākas kustības. Tie kļuva iespējami, pateicoties attīstībai, diferenciācijai nervu sistēma un kustības mehānisms un to harmonizācija.
Mīkstmieši, kas dzīvo bez fiksēta skeleta, iziet cauri viļņveidīgām muskuļu gredzenu peristaltiskām kontrakcijām. Muskuļu gredzenu segmentus kontrolē ganglija šūnas, kas ir gareniski savienotas ar nervu šķiedras lai sinhronizētu kustību.
Palielinot, kaulu cietajām un gludajām virsmām ir poraina struktūra. Pastāv dažādi veidi kaulu audi, kas ir visbiežāk dažādas daļas viens kauls: kompakts slānis un poraina viela. Kauli, piemēram, skriemeļi, augšstilba kakls, epifīze rādiuss, sastāv galvenokārt no porainas vielas. Sūkļainajā vielā kaulu sijas ir izvietotas izliektu plākšņu veidā, kas savienotas ar šķērseniski vai slīpi virzošiem šķērsstieņiem. Ekstremitāšu garie cauruļveida kauli sastāv galvenokārt no vielas, kurā kaulu plāksnes atrodas ļoti cieši.
Sākotnēji izkliedēta lokalizēta muskuļu šķiedras vēlāk veido mijiedarbojošus muskuļu saišķus; muskuļus. Fiksētā skeleta evolūcija ļāva strauji attīstīties, kas posmkājiem tika atrisināts atšķirīgi kā ārējais skelets, mugurkaulniekiem - kā iekšējais skelets. Lai gan abi efektīvi palīdz ķermeņa kustībai, pēdējais risinājums izrādījās izdevīgāks, jo tas ļāva netraucēti augt, ļaujot izveidot lielākas, sarežģītākas dzīvas būtnes.
Posmkāju nervu sistēma ir segmentēta. Ekstremitāšu aktivitāti mediālajā segmentā kontrolē segmentālie gangliji. Posmkāju gangliji var darboties neatkarīgi. Lai gan ganglijs galvā ir lielāks par otru, tas nekontrolē citu gangliju darbību. Tā izmērs drīzāk nodrošina acis, antenas un mutes instrumentus, kas tiek glabāti uz galvas. Kad posmkāja galva ir nogriezta, dzīvnieks var pārvietoties arī vairākas dienas, līdz nomirst bez mutes. Starpganglioniskie savienojumi nodrošina ekstremitāšu kustību koordināciju.
Kauli kā citi iekšējie orgāni sastāv no šūnām. Ir īpašas šūnas, kas pastāvīgi iznīcina kaulu vielu (osteoklastus); šūnas, kas atjauno, atjauno kaulus (osteoblastus), un šūnas, kas ir atbildīgas par kaula skeleta veidošanos un kaulu audu (osteocītu) mineralizāciju.
Cilvēka dzīves laikā kaulos nepārtraukti notiek kaulaudu pārstrukturēšanās procesi: kādā atsevišķi paņemtā nelielā kaulaudu laukumā kaulaudi tiek iznīcināti, pēc tam izņemto veco kaulu aizstāj ar tieši tādu pašu daudzumu jauna kaula. . Veseliem cilvēkiem kaulu audu iznīcināšanas un jauna kaula veidošanās procesi kvantitatīvi ir vienādi. Šo šūnu darbību kontrolē daudzas bioloģiski aktīvas vielas, piemēram, vairogdziedzera un epitēlijķermenīšu hormoni, virsnieru garozas hormoni, D3 vitamīns un, visbeidzot, dzimumhormoni (estrogēni un progesterons). Kaulu audu augšana un attīstība notiek līdz 16-25 gadiem. Pēc maksimālā kaulu masas līmeņa sasniegšanas līdz 30-40 gadu vecumam sākas tā nenozīmīgs zudums, kas ir 0,2-0,5% gadā.
Salīdzinot ar viņu sarežģīta funkcija, ganglijā tiek atrasts pārsteidzoši maz nervu šūnu. Viss ekstremitātes muskulis parasti tiek apgādāts ar četriem neironiem. Savienojumi starp neironiem nodrošina savstarpēju inervāciju starp saliecējiem un ekstensoriem un kustību koordināciju ar citām ekstremitātēm.
Pirmie mugurkaulnieki ir dzīvnieki, kas dzīvo ūdenī; Zivis. Sākotnēji viņu skeleta muskuļi sastāvēja tikai no aksiāliem muskuļiem, kurus absorbēja mugurkaula skābe un tika iegūti no miotomas. Muskuļi nodrošina tikai ķermeņa saliekšanu un pagriešanos. Pirmā viendabīgā aksiālā muskuļu masa vēlāk tiek segmentēta; tādējādi attīstījās atsevišķi muskuļi. Tad muskuļi tika iestiepti arī ķermeņa sienā. Kā nākamais evolūcijas rezultāts, spuras un pēc tam, kad daži uzdrošinājās nokļūt kontinentā, izvietoja ekstremitātes.
30-40 gadu vecumā kaulu zudums ir 0,5% gadā. Un pēc menopauzes sākuma sievietēm gadā tiek zaudēti 3-5% kaulu masas.
Cilvēka ķermenī ir izolēts arī mīksts skelets (skelets), kas piedalās orgānu noturēšanā pie kauliem. Mīkstais skelets ietver fascijas, saites, orgānu saistaudu kapsulas un citas struktūras. Lielākā daļa muskuļu ir pievienoti kauliem. Muskuļi ietver kustībā esošos skeleta kaulus un veic darbu. Daudzi muskuļi, kas ieskauj ķermeņa dobumus, aizsargā iekšējos orgānus.
Muskuļi, kas nodrošināja šīs struktūras, izlauzās no ķermeņa sienas. Ekstremitāšu attīstība bija lēciens mugurkaulnieku kustības sistēmas attīstībā. Ekstremitātes ļauj dzīvām būtnēm pārvietoties efektīvāk, tāpēc ir būtiski uzlabojušies to dzīves apstākļi, pielāgošanās videi. Evolūcijas gaitā to struktūra un izmērs ir pastāvīgi uzlaboti. Ekstremitātes, kas ir ievērojami lielākas par ķermeņa izmēru, pieļauj divu kāju dzīvi. Rezultātā viņi varēja labāk orientēties ar augstākiem maņu orgāniem galvā, un rokas, atbrīvotas no pārvietošanās slodzes, varēja izmantot citiem mērķiem.
Kaulu stāvoklis ir atkarīgs no to slodzes. Labi attīstīti muskuļu audi stiprina locītavas un veicina normālu kaulu attīstību un darbību. Gan muskuļi, gan kauli zaudē savu masu, ja slodze uz tiem ir pārāk maza. Tāpēc, lai ilgstoši uzturētu muskuļu un skeleta sistēmu veselīgā stāvoklī, nepieciešams pastāvīgi trenēties, veikt dažādas fiziski vingrinājumi. Katrs cilvēks pēc 30 gadu vecuma ir lemts fiziskajai audzināšanai.
Tie un atbilstošā roku mazo locītavu pārkārtošanās rosināja izmantot instrumentus, kas arī paātrināja smadzeņu attīstību, kas bija svarīgākais antropoīdu pielāgošanās orgāns. Divpusība īpaši uzsvēra motoriskās sistēmas attīstību un vienlaikus veicināja, jo ķermeņa vertikālā pozīcija prasa daudz lielāku kustību koordināciju nekā stāvēšana četru pēdu augstumā.
Ekstremitāšu paplašināšanās mainīja arī muskuļus. Daudz lielākā pieejamā vietā ir atļauta gan reizināšana, gan ekstremitāšu pagarināšana. Tie palīdz palielināt gan muskuļu spēku, gan mazāku kustību veiktspēju. Kustību mehānisma attīstība ir devusi jaunas iespējas mugurkaulniekiem. Viņu kustība kļūst perfektāka, ātrāka, precīzāka un atbilstošāka. Nepieciešami izstrādātas nervu struktūras, kas kontrolē skeleta muskuļu darbību. Kustību kustību veicināja vienlaikus attīstot maņu orgānus.
Tas ir īpaši svarīgi sievietēm, kuru kauli ir mazāk blīvi nekā vīriešiem. Turklāt sievietes vecumā ir vairāk pakļautas muskuļu masas zudumam.
Var kontrolēt kaulu masu, veidošanās pakāpi un zudumu labs uzturs. Kalcijs un D vitamīns stiprina kaulus un novērš to plaisāšanu, lūzumu un citus ievainojumus. Nepieciešamā kalcija dienas deva pieaugušajiem ir aptuveni 1200-1500 mg. Sievietes, tāpat kā vīrieši, līdz 20 gadu vecumam iegūst 50% no kaulu masas. Kombinācijā ar regulārām fiziskām aktivitātēm kalcijs veicina veselīgu, spēcīgu kaulu masu. Vecumā no 20 līdz 30 gadiem kaulu masa nedaudz palielinās, un pēc 30 gadiem sākas tā zuduma process.
Ar viņu palīdzību cilvēks var iegūt arvien pilnīgāku un precīzāku informāciju no apkārtējās vides. Lai izmantotu šīs iespējas, smadzenēs izplatījās neironi, kas atbalsta kustību koordināciju, un, lai tie būtu aktīvi, tie veidoja arvien sarežģītākus un intensīvākus savienojumus ar maņu orgāniem. Aksiālos muskuļus sākotnēji kontrolēja tikai mugurkaula motoriskie neironi. Pēc atsevišķu muskuļu atdalīšanas kontroli pār atsevišķiem muskuļiem pārņēma smadzenes, kas attīstās mugurkaulniekiem un spēlē centrālo kontroles lomu.
Saistībā ar jūsu ķermeņa bioloģisko funkciju, īpaša uzmanība veselīga ēšana jādod sievietēm. Grūtniecība un barošana ar krūti nepieciešams papildu kalcijs. Ja sievietes asinis nesatur nepieciešamo summu No šī minerāla kauli kļūst par papildu kalcija avotu, kas galu galā noved pie to vājināšanās. smēķēšana un pārmērīga lietošana alkoholiskie dzērieni veicina arī kaulu zudumu. Hormons estrogēns regulē kalcija izskalošanos no kauliem un tādējādi rada normālus apstākļus ilgstošai kaulu augšanai. Tomēr menopauzes un pēcmenopauzes periodā sievietes ķermenis pārstāj ražot estrogēnu, kas ir galvenais paātrinātā kaulu masas zuduma cēlonis. Regulāras fiziskās aktivitātes un palielināta ikdienas kalcija deva palīdzēs novērst šo procesu pēcmenstruālā periodā. Papildus kalcija uzņemšanai kā uztura bagātinātājam ikdienas uzturā ieteicams iekļaut pārtiku, kas satur šo elementu.
Pirmais mugurkaulnieku autocentrs bija lokomotoriskais reģions, kas atradās mesencefalonā. Lai gan evolūcija ir attīstījusies līdz jaunām smadzeņu zonām, kas specializējas motoru kontrolē, retikulospastiskais trakts saglabājas arī cilvēkiem. Papildus muskuļu tonusa kontrolei viņš galvenokārt nodarbojas ar ritmisku kustību izpildi.
Evolūcijas laikā mugurkaulnieku smadzenes sākotnēji dominē mūsdienu smadzeņu stumbra daļā. Tā kā smadzeņu funkcijas kļuva sarežģītākas, bija nepieciešams vairāk nervu šūnu, lai tās kontrolētu. Tā kā tobrīd smadzenēs nebija pietiekami daudz vietas, jauni neironi veidoja jaunu koloniju līdz šim atradušo smadzeņu priekšā. Nākamajā solī veidojas trīs kodoli, kas atbild par redzi: divi aiz un viens virs vidussmadzenēm. Iepriekšējiem mugurkaulniekiem ir trīs acis. Abu sānu acu informāciju apstrādāja divi bazalokatīvi sānu dzimumlocekļi, kurus iepriekš minētās parietālās acis šķērso caur augšējo kodolu.
Uz programmu fiziskās aktivitātes jāiekļauj svara treniņš. Svarīgs faktors, kas nosaka muskuļu un skeleta sistēmas veselību, ir optimāla svara uzturēšana.
Īpaša uzmanība jāpievērš kājām. Kājām ir ļoti liela nozīme balsta un kustību aparāta sistēmā, kā arī visa organisma veselībā. Pareiza aprūpe aiz kājām papildus citām higiēnas procedūrām ietver rūpīgu apavu izvēli. Ērti un pareizi izvēlēti apavi ir lieliska daudzu pēdu slimību profilakse.
Arī sānu acis un to kodoli cilvēkam saglabājās kā redzes orgāni, tajā pašā laikā parietālā acs un tās kodols pamazām atkāpās, zaudēja redzi un nogrima dziļumā kā epitalāms. Nākamā funkcija diencefalonā bija somatosensorā. Rāpuļiem somatosensorās funkcijas ārstēšana turpināja attīstīties; attīstījās muguras un priekšējie talāmu kodoli. Tajā pašā laikā anteroventrālais kodols bija pirmais meencefalona koordinācijas centrs.
Pēdējais talāmu apgabals, kas attīstījās, bija Pulvinar Talami, kas apstrādāja maņu orgānu koordināciju. Nezīdītājiem tas pārņēma visu smadzeņu trīs sensoro daivu lomu. Bazālie gangliji, kas koordinē kustību sistēmu, jau ir atrasti abiniekiem. Tie strauji attīstījās, bet tikai zīdītājiem.
"Skeleta-muskuļu sistēma" un citi raksti no sadaļas Skeleta-muskuļu sistēmas slimības
Brīvprātīgās kustības funkcionālā struktūra. No iepriekš minētā izriet, ka jebkuras kustības nodrošināšanā ir iesaistītas dažādas sastāvdaļas, tāpēc viens no galvenajiem jautājumiem ir, kā nodrošināt izpildaparāta saņemto vienreizējo komandu. Neatkarīgi no konkrētās kustības stratēģijas un taktikas, programmas, kas nodrošina programmu, galvenais uzdevums ir visu komandas komponentu koordinēšana.
Urāna telencefalona nogulsnes, kas izstieptas no diēna šūnas sānu sienām, jau veidojas abiniekiem. Pirmās telencefālās struktūras atbilst bazālo gangliju un zīdītāju limbiskās sistēmas augiem. Viņu galvenie uzdevumi bija ožas sajūta un noteiktas, īpaši seksuālas, uzvedības koordinēšana. Smadzeņu garoza ir zīdītāju "izgudrojums". Saskaņā ar iepriekš minēto, tiek atrastas tādas jaunas funkcijas, kuru neironi nav atraduši pietiekami daudz vietas starpencefalonā.
Tā kā diencefalons vairs nevarēja augt, no neiroepitēlija caurules sienas, kas izauga no tās sienas, radās jauni neironi. Sānu kambari pakāpeniski attīstījās, un encefalona daivas no to sienām. Pirmie zīdītāji bija mazi, stieņveida grauzēji, kas sākumā šaudījās apkārt un pēc tam kļuva kā maisiņš. Dinozauru laikmetā viņi patvērās metro alās. Atbilstoši savam dzīvesveidam tumšās alās viņi pārvietojās galvenokārt pēc smaržas.
CNS ir vairākas ģenētiski fiksētas programmas (piemēram, kustību staigāšanas programma, kuras pamatā ir aktivitāte mugurkaula - kas attiecas uz mugurkaulu.");" onmouseout = "nd();" href="javascript:void(0);">mugurkaulaģenerators). Šādas vienkāršas programmas tiek apvienotas sarežģītākās sistēmās, piemēram, taisnas pozas uzturēšanai. Šī asociācija rodas mācīšanās rezultātā, kas tiek nodrošināta, piedaloties priekšējai garozai. puslodes.
Sarežģītākā un filoģenētiski jaunākā ir spēja veidot kustību secību un paredzēt tās īstenošanu. Šīs problēmas risinājums ir saistīts ar frontālo asociatīvo sistēmu, kas iegaumē un saglabā atmiņā šādas kustību secības. Šīs kodēšanas augstākais atspoguļojums cilvēkā ir kustības pamatjēdzienu verbalizācija jeb verbālais pavadījums.
Kustības kontroles sistēmas vispārējā likumsakarība ir atgriezeniskās saites izmantošana. Tas ietver ne tikai proprioceptīvu atgriezenisko saiti no kustības, kas ir sākusies, bet arī Aktivizācija - uzbudinājums vai paaugstināta aktivitāte, pāreja no miera stāvokļa uz aktīvu stāvokli. onmouseout = "nd();" href="javascript:void(0);">aktivizēšana atlīdzības vai soda sistēmas. Turklāt tiek iekļauta arī iekšējā atgriezeniskā saite, t.i. informācija par motora sistēmas pamatā esošo līmeņu darbību vai pašas motora komandas efektīvā kopija. Šāda veida atgriezeniskā saite ir nepieciešama jaunu kustību koordināciju attīstībai. Dažādas sarežģītības un ātruma kustībām atgriezenisko saiti var aizvērt dažādos līmeņos. Tāpēc abi vadības veidi - programmēšana un izsekošana - var pastāvēt vienā un tajā pašā kustības vadības sistēmā.
Noslēgumā vēlams minēt izcilā fiziologa N.A. Bernsteins, ka kustības "...nav telpiskas, bet gan semantiskas un darbības līmeņa ķēžu motoriskās sastāvdaļas tiek diktētas un atlasītas atbilstoši objekta semantiskajai būtībai un tam, kas ar to būtu jādara."
Lai izdzīvotu, divas vissvarīgākās funkcijas ir smarža un reprodukcija. Tādējādi šīs divas funkcijas ir risinātas divos jaunos diencefalona jaunattīstības reģionos. viņu paliekas, limbiskā sistēma un smakojošā garoza, kas pilda tās pašas funkcijas, ir atrodama arī cilvēkiem mediālajā centrālajā garozā. Šīs divas funkcijas ir cieši saistītas morfoloģiski un funkcionāli katram mugurkaulniekam, pat cilvēkiem ar ievērojami samazinātu smaku. Parfimērijas industrija balstās uz šādu attieksmi.
Corpus Callosum puslodes nav vecāko izdzīvojušo zīdītāju, kloako un putnu smadzenēs. Nākamais evolūcijas solis bija ievērojama somatosensorās sistēmas attīstība. Šie nakts grauzēji, līdzīgi mūsdienu žurkām un pelēm, joprojām dzīvoja tumšajā pazemē. Viņi varēja ātri un droši pārvietoties pa tumšajiem pazemes kanāliem gari mati uz sejas, jo viņi varēja tos izmantot kā radaru, izmantojot ļoti jutīgus spiediena receptorus, kas savienoti. Šai precīzai mehāniskai “attēla analīzei” viņiem bija nepieciešama jauna neironu populācija.
10.4. Kustību organizācijas elektrofizioloģiskās korelācijas
Dažādu aspektu pētīšanai tiek izmantotas elektrofizioloģiskās metodes motora aktivitāte, un jo īpaši tie, kas nav pieejami tiešai novērošanai. Vērtīgu informāciju par kustību organizācijas fizioloģiskajiem mehānismiem sniedz metodes smadzeņu garozas zonu mijiedarbības novērtēšanai, lokālo EEG un ar kustību saistīto potenciālu analīzei un neironu aktivitātes fiksēšanai.
Smadzeņu biopotenciālu starpzonālo savienojumu izpēte ļauj izsekot atsevišķu garozas zonu mijiedarbības dinamikai dažādos kustību izpildes posmos, apgūstot jaunas motorikas, identificēt starpzonu mijiedarbības specifiku laikā. dažādi veidi kustības.
Šie neironi migrēja uz jaunizveidoto teleencefalonu un izveidoja pirmo īsto daivu - parietālā daiva. Šis atloks palika arī cilvēka somatosensorais centrs. Oposiem un līdzīgām sugām smadzeņu garozu galvenokārt izmanto ādas dziedināšanai. Veciem zīdītājiem signāli virzījās no muskuļu vārpstām uz jaunizveidotajām smadzenītēm un no turienes caur smadzenīšu kodoliem uz talāmu.
Nākamajā evolūcijas posmā trušiem līdzīgu grauzēju smadzenes, kas jau devās uz zemes virsmu, saskārās ar jaunu izaicinājumu: viņiem bija jāieklausās no attāluma un jāatrod potenciālais uzbrūkošais ienaidnieks, lai tikai aizbēgtu laiks. Jaunais akustiskais centrs ir pārcēlies no apakšējā colliculus uz temporālā daiva, kas attīstās no neiroepitēlija caurules sienas, kas izaug no dienosifalona un vietas trūkuma dēļ šūpojas uz leju. Daļas sānu kambara, kas satur divas pinuma asinsvadu sienas, šūnu barotnes un apakšējās kukurūzas, kā arī parietālo un temporālā daiva, kas attīstās no tās sienas, ir vecākās smadzeņu garozas daļas.
Telpiskā sinhronizācija (PS), t.i. No dažādiem smadzeņu garozas punktiem fiksētā elektrisko svārstību sinhronā dinamika atspoguļo tādu smadzeņu struktūru stāvokli, kurā tiek veicināta ierosmes izplatīšanās un radīti apstākļi starpzonu mijiedarbībai. PS reģistrācijas metodi izstrādāja izcilais krievu fiziologs M.N. Livanovs.
Atsevišķu zonu EEG ritmisko komponentu un to telpisko un laika attiecību pētījumi cilvēkiem, veicot brīvprātīgas kustības, deva reāla iespēja tuvoties funkcionālās mijiedarbības centrālo mehānismu analīzei, kas veidojas sistēmas līmenī motoriskās aktivitātes laikā. Ritmisko kustību veikšanas laikā reģistrētās EEG korelācijas analīze parādīja, ka cilvēkiem kortikālajā kustību organizācijā piedalās ne tikai motorās garozas centri, bet arī frontālās un apakšējās parietālās zonas.
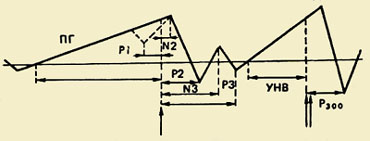
Brīvprātīgo kustību apgūšana un to apmācība izraisa kortikālo biopotenciālu starpcentrālo korelāciju pārdali. Treniņa sākumā kopējais skaits krasi palielinās kopīgās aktivitātēs iesaistītie centri, un motoro zonu EEG ritmisko komponentu sinhronās attiecības ar priekšējo un aizmugurējo. Garozas asociatīvās zonas - zonas, kas saņem informāciju no receptoriem, kas uztver dažādu modalitātes kairinājumu, un no visām projekcijas zonām. ");" onmouseout = "nd();" href="javascript:void(0);">asociatīvs apgabali. Apgūstot kustību, kopējais PS līmenis ievērojami samazinās, un, gluži pretēji, palielinās savienojumi starp motoriskajām zonām un apakšējām parietālajām. 
Ir svarīgi atzīmēt, ka mācību procesā tiek pārkārtots dažādu garozas zonu biopotenciālu ritmiskais sastāvs: EEG sāk reģistrēties lēni ritmi, kas biežumā sakrīt ar kustību izpildes ritmu. Šos cilvēka EEG ritmus sauc par "marķētiem". Tādas pašas iezīmētās svārstības tika konstatētas bērniem pirmsskolas vecums kad viņi veic ritmiskas kustības uz ergogrāfa.
Sistemātiski cilvēka EEG pētījumi cikliskās (periodiski atkārtojas) un acikliskās motoriskās aktivitātes īstenošanas laikā atklāja būtiskas izmaiņas smadzeņu garozas elektriskās aktivitātes dinamikā. EEG tiek novērota gan lokālās, gan attālās biopotenciālu sinhronizācijas palielināšanās, kas izpaužas kā periodisko komponentu jaudas palielināšanās, auto- un krustenisko korelogrammu frekvenču spektra izmaiņās, noteiktā saskaņojumā. frekvenču spektru un funkciju maksimumi Saskaņotība - EEG frekvences indikatoru sinhronizācijas pakāpe starp dažādām smadzeņu garozas daļām.");" onmouseout = "nd();" href="javascript:void(0);">saskaņotība tajā pašā frekvencē.
PS un reakcijas laiks. Reakcijas laiks ir viens no vienkāršākajiem motora indikatoriem. Tāpēc īpaši interesants ir fakts, ka pat vienkāršai motoriskai reakcijai var būt dažādas fizioloģiskas korelācijas atkarībā no tās ilguma palielināšanās vai samazināšanās. Tādējādi, salīdzinot smadzeņu EEG spektrālo komponentu starpcentrālo korelācijas attiecību attēlu ar vienkāršas motoriskās reakcijas laiku, izrādījās, ka asociatīvo zonu EEG spatiotemporālo attiecību pārkārtošanās ir saistīta ar reakciju. laiks līdz noteiktam stimulam. Ar straujām reakcijām veselam cilvēkam visbiežāk augstas biopotenciālu korelācijas radās abās apakšējās parietālajās zonās (nedaudz vairāk ar kreiso smadzeņu puslodi). Ja reakcijas laiks palielinājās, tam sekoja biopotenciālu sinhronizācija frontālie reģioni garoza un kreisās puslodes apakšējais parietālais reģions tika izslēgts no mijiedarbības. Turklāt tika atrasta sakarība starp fāzes nobīdes lielumiem Alfa ritms - galvenais elektroencefalogrammas ritms relatīvā miera stāvoklī, ar frekvenci diapazonā no 8 - 14 Hz un vidējo amplitūdu 30 - 70 μV. ");" onmouseout = "nd();" href="javascript:void(0);">alfa ritms reģistrēts frontālajā, precentrālajā un pakauša zonas smadzenes un vienkāršas motora reakcijas ātrums.
Svarīgi atzīmēt, ka biopotenciālu sinhronizācijas palielināšanās notiek cilvēkam jau pirmsdarba stāvoklī koncentrēšanās procesā pirms motoriskās darbības, kā arī kustību garīgās izpildes laikā.
PS un kustības specifika. Papildus nespecifiskajam biopotenciālu PS pieaugumam tika atzīmēts tā izteiktais selektīvais pieaugums starp garozas zonām, kas tieši iesaistītas konkrēta motora akta organizēšanā. Piemēram, tiek konstatēta lielākā elektriskās aktivitātes līdzība: kustinot rokas - starp frontālo reģionu un muskuļu motorisko attēlojumu. augšējās ekstremitātes; kustinot kājas - starp frontālo reģionu un muskuļu motorisko attēlojumu apakšējās ekstremitātes. Ar precīzām darbībām, kurām nepieciešama precīza telpiskā orientācija un vizuālā kontrole (šaušana, paukošana, basketbols), tiek uzlabota mijiedarbība starp vizuālo un motorisko zonu.
Sportistu dažādu smadzeņu daļu biopotenciālu sarežģītā PS dinamika tika atklāta, veicot dažādi vingrinājumi un parādīta EEG ritmisko komponentu mijiedarbības pieauguma atkarība no motoriskās aktivitātes veida, no sportistu kvalifikācijas, no cilvēka spējas risināt taktiskās problēmas, no situācijas sarežģītības. Tādējādi augsti kvalificētiem sportistiem starpcentru mijiedarbība ir daudz intensīvāka un skaidrāk lokalizēta. Izrādījās arī, ka sarežģītāki motora uzdevumi prasa vairāk augsts līmenis EEG ritmu telpiskā sinhronizācija, un taktisko uzdevumu risināšanas laiks korelē ar starpcentru mijiedarbības pieauguma ātrumu. Šajā gadījumā motora reakcija seko pēc maksimālās biopotenciālu sinhronizācijas sasniegšanas smadzeņu garozā.
Kopā cilvēka smadzeņu biopotenciālu PS pētījumi ir ļāvuši konstatēt, ka, veicot vienkāršas un sarežģītas motoriskās darbības, dažādi smadzeņu centri iesaistās mijiedarbībā, veidojot sarežģītas savstarpēji saistītu zonu sistēmas ar aktivitātes perēkļiem ne tikai projekcijā, bet arī asociatīvajās zonās, īpaši frontālajā un apakšējā parietālajā. Šīs starpcentrālās mijiedarbības ir dinamiskas un mainās laikā un telpā, veicot motora darbību.
10.5. Smadzeņu potenciālu komplekss, kas saistīts ar kustībām
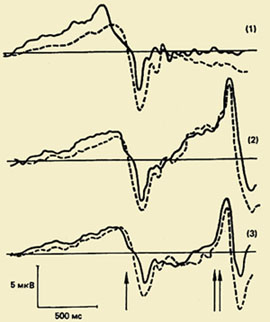 Viens no svarīgākajiem virzieniem motora akta psihofizioloģijas izpētē ir ar kustībām saistīto smadzeņu potenciālu svārstību kompleksa izpēte (PMSD). Šīs parādības nozīme izpratnei fizioloģiskie mehānismi kustību organizācija ir ļoti apjomīga, jo PMSD izpēte ļauj atklāt smadzeņu garozā notiekošo procesu slēpto secību kustību sagatavošanas un izpildes laikā, un informācijas apstrādes procesu hronometrija ir metožu kopums smadzeņu garozā notiekošo procesu mērīšanai. uz mērījumiem balstītas informācijas apstrādes procesa atsevišķu posmu ilgums fizioloģiskie rādītāji, jo īpaši izsaukto un ar notikumu saistīto potenciālu komponentu latentie periodi.");" onmouseout = "nd();" href="javascript:void(0);"> pulkstenisšie procesi, t.i. noteikt savus laika ierobežojumus.
Viens no svarīgākajiem virzieniem motora akta psihofizioloģijas izpētē ir ar kustībām saistīto smadzeņu potenciālu svārstību kompleksa izpēte (PMSD). Šīs parādības nozīme izpratnei fizioloģiskie mehānismi kustību organizācija ir ļoti apjomīga, jo PMSD izpēte ļauj atklāt smadzeņu garozā notiekošo procesu slēpto secību kustību sagatavošanas un izpildes laikā, un informācijas apstrādes procesu hronometrija ir metožu kopums smadzeņu garozā notiekošo procesu mērīšanai. uz mērījumiem balstītas informācijas apstrādes procesa atsevišķu posmu ilgums fizioloģiskie rādītāji, jo īpaši izsaukto un ar notikumu saistīto potenciālu komponentu latentie periodi.");" onmouseout = "nd();" href="javascript:void(0);"> pulkstenisšie procesi, t.i. noteikt savus laika ierobežojumus.
PMSD komponentu sastāvs. Pirmo reizi šis komplekss, kas atspoguļo kustības sagatavošanas, izpildes un izvērtēšanas procesus, reģistrēts 60. gados. Izrādījās, ka pirms kustības notiek lēna negatīva svārstība – gatavības potenciāls (RP). Tas sāk attīstīties 1,5 - 0,5 s pirms kustības sākuma. Šis komponents ir reģistrēts galvenokārt abu pusložu centrālajā un fronto-centrālajā vadā. 500-300 ms pirms kustības sākuma PG kļūst asimetrisks - tā maksimālā amplitūda tiek novērota precentrālajā reģionā, Kontralaterāls - attiecas uz pretējo ķermeņa pusi.");" onmouseout = "nd();" href="javascript:void(0);"> kontralaterāls kustība. Apmēram pusei pieaugušo uz šo lēno negatīvo svārstību fona īsi pirms kustības sākuma tiek reģistrēta neliela amplitūdas pozitīva sastāvdaļa. To sauc par "premotoru pozitivitāti" (PMP). Nākamā strauji augošā negatīvā amplitūdas svārstība, tā sauktais motora potenciāls (MP), sāk attīstīties 150 ms pirms kustības sākuma un sasniedz maksimālo amplitūdu virs kustīgās ekstremitātes motora attēlojuma apgabalā smadzeņu garozā. Šis potenciālu komplekss beidzas ar pozitīvu komponentu aptuveni 200 ms pēc kustības sākuma.
Komponentu funkcionālā nozīme. Ir vispāratzīts, ka gatavības potenciāls (RP) rodas motorajā garozā un ir saistīts ar kustību plānošanas un sagatavošanas procesiem. Tas pieder pie smadzeņu potenciāla lēno negatīvo svārstību klases, kuru rašanās ir izskaidrojama ar attiecīgo garozas zonu neironu elementu aktivāciju.
Hipotēzes par funkcionālā vērtība PMP ir dažādi.
Šīs svārstības tiek uzskatītas gan par centrālās komandas nodošanas no garozas muskuļiem atspoguļojumu, gan kā garozas relaksācijas rezultāts pēc noteikta kustību organizācijas posma pabeigšanas, gan kā procesu atspoguļojums. otras ekstremitātes saistīto kustību nomākšanai un kā atgriezeniskā saite no muskuļu aferentiem. Pašlaik daži autori uzskata, ka PMP ir tikai motora potenciāla sākuma atspoguļojums.
Reģistrējot MP pērtiķiem, MP tika identificētas divas apakškomponentes. Pirmais apakškomponents ir saistīts ar motorās garozas aktivāciju, kas saistīta ar kustības uzsākšanu (sinaptiskā aktivitāte piramīdveida neironi), bet otrs - ar Brodmaņa lauku 2.3. un 4. lauka aktivizēšanu - atsevišķi smadzeņu garozas apgabali, kas atšķiras pēc to šūnu struktūras (citoarhitektonikas) un funkcijām. Piemēram, lauki 17,18,19 ir smadzeņu garozas vizuālie apgabali, kuriem ir dažādas struktūras un funkcijas vizuālās uztveres nodrošināšanā.");" onmouseout = "nd();" href="javascript:void(0);">autors Brodmens . MP reģistrācija cilvēkam ar epilepsiju ļāva tajā atšķirt trīs sastāvdaļas. Pirmo komponentu sauca par iniciācijas potenciālu. Tam ir liela amplitūda un tas notiek pēc kustības sākuma precentrālajā kontralaterālajā garozā. Otrs, kas rodas pēc miogrammas sākuma un ir vairāk lokalizēts kontralaterālajā somatosensorajā laukā, var būt saistīts gan ar kustību ierosināšanu, gan sensoro atgriezenisko saiti. Trešais komponents atspoguļo impulsu, kas nāk no muskuļu aferentiem uz garozu.
Pozitīvais potenciāls, kas seko MP, tiek uzskatīts par reversās aferentācijas atspoguļojumu, kas nāk no perifērajiem receptoriem, augšupejošās aktivitātes no motora centriem, salīdzināšanas darbību starp motoru programmu un tās izpildes neironu attēlu vai kortikālo procesu. relaksācija pēc kustības.
Gaidīšanas vilnis. Papildus PMSD ir aprakstīta vēl viena elektrofizioloģiska parādība, kas būtībā ir tuvu gatavības potenciālam. Tas ir par par potenciāla negatīvajām svārstībām, kas reģistrētas smadzeņu garozas priekšējās daļās laika posmā starp brīdinājuma un sprūda (nepieciešama reakcija) signālu darbību. Šīm svārstībām ir vairāki nosaukumi: gaidīšanas vilnis, E-vilnis, nosacītā negatīvā novirze (CNV). E-vilnis parādās 500 ms pēc brīdinājuma signāla, tā ilgums palielinās, palielinoties intervālam starp pirmo un otro stimulu. E-viļņa amplitūda palielinās tieši proporcionāli motora reakcijas ātrumam uz sākuma stimulu. Tas palielinās līdz ar uzmanības spriedzi un gribas piepūles palielināšanos, kas norāda uz šīs elektrofizioloģiskās parādības saistību ar motora aktivitātes un uzvedības brīvprātīgas regulēšanas mehānismiem kopumā.
10.6. nervu darbība
Funkcionālās kortikālās kolonnas. Cilvēka garozas motoriskajā zonā atrodas tā sauktās milzu piramīdas Beza šūnas - smadzeņu garozas piramīdas šūnas. onmouseout = "nd();" href="javascript:void(0);">Betz šūnas, kas ir sakārtoti atsevišķās kolonnās. Piramīdveida šūnas, kas veic līdzīgas funkcijas, atrodas blakus, pretējā gadījumā būtu grūti izskaidrot precīzu garozas somatotopisko organizāciju. Šādas motoru kolonnas spēj ierosināt vai kavēt funkcionāli viendabīgu grupu Motoru neirons (motoru neirons) - nervu šūna, kuras aksons inervē muskuļu šķiedras. onmouseout = "nd();" href="javascript:void(0);">motoriskie neironi.
Atsevišķu piramīdas šūnu aktivitātes reģistrēšana, izmantojot implantētus mikroelektrodus dzīvniekiem, kuri veic dažādas kustības, ļāva konstatēt fundamentāli svarīgu faktu. Kortikālie neironi, kas regulē jebkura muskuļa darbību, nav koncentrēti tikai vienā kolonnā. Motora kolonna lielākoties ir funkcionāla neironu apvienība, kas regulē vairāku muskuļu darbību, kas iedarbojas uz noteiktu locītavu. Tādējādi motora garozas piramīdveida neironu kolonnās kustības tiek attēlotas ne tik daudz, cik muskuļi.
Motoru programmu neironu kodi. Informācijas kodēšana neironā tiek veikta pēc tā izlādes biežuma. Neironu impulsu aktivitātes analīze dažādu motoru programmu izstrādes laikā dzīvniekiem parādīja, ka neironi ir iesaistīti to konstrukcijā. dažādas nodaļas motoru sistēma, vienlaikus veicot noteiktas funkcijas. Saskaņā ar dažām idejām motoru programmu iekļaušana notiek tā saukto komandu neironu aktivizēšanas dēļ. Savukārt komandu neironi atrodas augstāko garozas centru kontrolē. Inhibīcija ir nervu process, kas ir pretējs ierosināšanai; izpaužas noteiktai ķermeņa sistēmai raksturīgu darbību pavājināšanās vai pārtraukšanā. onmouseout = "nd();" href="javascript:void(0);">Palēnināt komandu neirons noved pie tā vadītās programmas apstāšanās, ierosmes, gluži pretēji, pie nervu ķēdes aktivizēšanas un motoriskās programmas aktualizācijas.
Komandneironu iesaistīšanos smadzeņu integrālajā darbībā nosaka pašreizējā motivācija un konkrēta motora programma, kas vērsta uz šīs motivācijas apmierināšanu. Motora programma, lai pēc būtības būtu adaptīvs, jāņem vērā visas signāli nozīmīgās ārējās vides sastāvdaļas, attiecībā pret kurām tiek veikta mērķtiecīga kustība, t.i. pamatojoties uz multisensoru principu Konverģence ir neironu grupas aksonu asociācija, kas rodas sinapšu veidošanās dēļ uz viena un tā paša postsinaptiskā neirona. onmouseout = "nd();" href="javascript:void(0);">konverģence.
Terminu vārdnīca
- pārvietošanās
- motorais neirons
- artikulācija
- aktivizēšana
- saskaņotību
- Brodmaņa lauki
- informācijas apstrādes procesu hronometrija
Jautājumi pašpārbaudei
- Kāda ir atšķirība starp statisko un dinamisko ķermeņa attēlu?
- Kurām smadzeņu struktūrām ir izšķiroša loma brīvprātīgas kustības organizēšanā?
- Kā piramidālās un ekstrapiramidālās sistēmas funkcijas atšķiras?
- Ko dod smadzeņu potenciāls, kas saistīts ar kustību, lai izprastu kustību organizācijas smadzeņu mehānismus?
Bibliogrāfija
- Batuev A.S. Motora analizatora funkcijas. L.: Nauka, 1970. gads.
- Batuev A.S. Augstāks nervu darbība. Maskava: Augstskola, 1991.
- Bernstein N.A. Esejas par kustību fizioloģiju un darbības fizioloģiju. Maskava: Nauka, 1966.
- Blūms F., Lizersons A., Hofstadters L. Smadzenes, prāts un uzvedība. M.: Mir, 1988.
- Dudel J., Ruegg I., Schmidt R., Janig V. Cilvēka fizioloģija. T. 1 / Red. R. Šmits un G. Tevs. M.: Mir, 1985.
- Cilvēka un dzīvnieku fizioloģijas vispārējais kurss / Red. A.A. Nozdračevs. Maskava: Augstskola, 1991.
- Sologubs E.B. Cilvēka smadzeņu elektriskā darbība motoriskās aktivitātes procesā. L.: Nauka, 1973. gads.
- Khrizman T.P. Bērna kustība un smadzeņu elektriskā aktivitāte. Maskava: Pedagoģija, 1973.
- Evarts E. Smadzeņu mehānismi, kas kontrolē kustību // Brain. M.: Mir, 1982.
Kursa darbu un eseju tēmas
- Mācības N.A. Bernsteins par kustības struktūru.
- Kustību psihofizioloģiskās tipoloģijas.
- Cilvēka roku kustības un to neirofizioloģiskie mehānismi.
- Brīvprātīgās kustības un to neirofizioloģiskie mehānismi.
- Ar kustībām saistīto smadzeņu potenciālu izpētes vēsturiskie posmi.
- Neironu aktivitātes nozīme kustību veidošanā.

