Reflex- reakcia organizmu na podráždenie receptorov, uskutočňovaná nervovým systémom.
reflexný oblúk - dráha, po ktorej prechádza nervový impulz pri realizácii reflexu. Pozostáva z 5 častí:
- receptor - citlivá formácia schopná reagovať na určitý typ podnetu;
- citlivý neurón vedie impulzy do mozgu;
- interkalárny neurón spája senzitívne a výkonné neuróny, nachádza sa v mieche alebo mozgu;
- výkonný (motorický) neurón vedie impulzy z mozgu do výkonných orgánov;
- výkonná agentúra- sval (kontrakty), žľaza (tajomstvo) atď.
Nepodmienené reflexy
- prítomný od narodenia
- počas života sa nemenia a nezmiznú;
- sú rovnaké vo všetkých organizmoch toho istého druhu;
- prispôsobiť telo konštantným podmienkam.
Podmienené reflexy
- získané počas života;
- môže sa zmeniť alebo zmiznúť;
- každý organizmus má svoje vlastné;
- prispôsobiť telo meniacim sa podmienkam.
Testy
1) Ktorý článok slúži ako začiatok reflexného oblúka
A) interkalárny neurón
B) citlivý neurón
B) receptor
D) výkonný neurón
2. V akom poradí sú zložky reflexného oblúka zahrnuté do realizácie reflexu
A) výkonný orgán, motorický neurón, interkalárny neurón, senzorický neurón, receptor
B) interkalárny neurón, senzorický neurón, motorický neurón, receptor, výkonný orgán
C) receptor, senzorický neurón, interkalárny neurón, motorický neurón, výkonný orgán
D) senzorický neurón, interkalárny neurón, receptor, výkonný orgán, motorický neurón
3. Počiatočné štádium reflexného oblúka v slinnom reflexe
A) slinná žľaza
B) receptor
B) interkalárny neurón
D) výkonný neurón
4. Pôsobenie podnetov vyvoláva výskyt nervový impulz V
A) senzorické neuróny
B) motorické neuróny
B) receptory
D) interkalárne neuróny
5. Vzruch smeruje pozdĺž citlivého neurónu
A) do centrálneho nervového systému
B) výkonným orgánom
B) na receptory
D) do svalov
6. Receptory sú citlivé útvary, ktoré
A) prenášať impulzy do centrálneho nervového systému
B) prenášať nervové impulzy z interkalárnych neurónov do exekutívy
C) vnímať podnety a premieňať energiu podnetov na proces nervovej excitácie
D) prijímať nervové impulzy zo senzorických neurónov
7. Nervové impulzy sa prenášajú do mozgu prostredníctvom neurónov
A) motor
B) interkalárne
B) citlivý
D) výkonný
8. Konce reflexného oblúka
A) výkonný pracovník
B) citlivý neurón
B) receptor
D) interkalárny neurón
9. Vedú nervové impulzy z receptorov do centrálneho nervového systému
A) senzorické neuróny
B) motorické neuróny
C) senzorické a motorické neuróny
D) interkalárne a motorické neuróny
10. Nervové formácie v ľudskom tele, vnímanie podráždení z vonkajšieho prostredia, sú
A) nervy
B) receptory
B) mozog
D) nervové uzliny
11. K premene vzruchov na nervový vzruch dochádza v
A) motorické nervy
B) mozog
B) receptory
D) miecha
12. Nervové impulzy sa prenášajú z ľudských zmyslov prostredníctvom neurónov
A) citlivý
B) motor
B) vložiť
D) výkonný
13. Vnímať informácie a premieňať ich na nervové vzruchy
A) receptory
B) nervové uzliny
B) výkonné orgány
D) interkalárne neuróny
14. Podmienený reflex
A) sa dedí po potomstve, ale nepretrváva po celý život
B) telo ľahko získava a stráca počas života
C) dedí potomstvo po rodičoch a pretrváva pre
D) je neustála reakcia organizmu na presne definované podnety prostredia
15. Príklad podmieneného reflexu u mačky
A) slinenie pri jedle
B) odtiahnutie labky pri popálení
B) reakcia na prezývku
D) reakcia na ostrý zvuk
16. Slinenie u ľudí pri pohľade na citrón - reflex
A) podmienené
B) bezpodmienečné
B) ochranný
D) orientačné
17) V procese sa u ľudí vytvárajú podmienené reflexy
A) individuálny rozvoj
B) tvorba druhov
C) historický vývoj
D) tvorba embryí
18. Príkladom podmieneného reflexu je
A) slinenie pri pohľade na rozrezaný citrón
B) slinenie počas kŕmenia
C) zúženie zrenice pri vystavení silnému svetlu
D) odtiahnutie ruky s náhlou injekciou
19. Reakcia dieťaťa na fľašu mlieka je reflex, ktorý
A) sa dedí
B) sa tvorí bez účasti kôry hemisféry
B) sú získané v priebehu života
D) pretrváva po celý život
20. Ochranný kýchací reflex
A) nezdedí sa
B) je podmienené
B) časom slabne
D) charakteristické pre všetkých jedincov druhu
21. Reakcie získané ľuďmi a zvieratami počas života a zabezpečujúce prispôsobenie sa meniacim sa podmienkam prostredia sa nazývajú
A) podmienené reflexy
B) inštinkty
B) nepodmienené reflexy
D) reflexy, ktoré sú zdedené
22. Nepodmienený reflex
A) sa dedí
B) získané počas života
C) sa vytvára pre určité signály
D) je základom rôznych vonkajších signálov
23. Nepodmienené reflexy ľudí a zvierat poskytujú
C) osvojenie si nových pohybových schopností tela
24. Reakcia človeka na zelená farba semafor je reflex
A) vrodené
B) získané
B) bezpodmienečné
D) zdedil
25. Zvýraznite tráviace šťavy ako odpoveď na potravinové stimulačné receptory ústna dutina
A) vzrušenie
B) brzdenie
B) nepodmienený reflex
D) samoregulácia
26. Biologický význam podmienených reflexov pre telo je v tom, že oni
A) udržiavať stálosť vnútorného prostredia
B) druhovo špecifické a pretrvávajú počas celého života
B) podporovať interakciu funkčné systémy
D) zabezpečiť adaptáciu na meniace sa podmienky prostredia
27. V procese života zvieratá získavajú reflexy
A) bezpodmienečné
B) zdedil
C) charakteristické pre všetkých jedincov daného druhu
D) umožniť im prežiť v meniacich sa podmienkach
28. Podmienený reflex u ľudí je
A) zúženie zrenice pri jasnom svetle
B) otočenie hlavy na ostrý zvuk
C) vylučovanie slín pri vstupe potravy do úst
D) reakcia na význam slova
29. Príkladom podmieneného reflexu je
1) schopnosť jazdiť na bicykli
2) slinenie pri jedle
3) túžba spať, keď je unavený
4) kýchanie a kašeľ
30. Podmienené reflexy ľudí a zvierat poskytujú
A) prispôsobenie organizmu konštantným podmienkam prostredia
B) prispôsobenie tela meniacemu sa vonkajšiemu svetu
C) rozvoj organizmov nových pohybových schopností
D) rozlíšenie medzi zvieratami podľa príkazov trénera
31. Vrodené reflexy
A) sú druhové charakteristiky organizmu
B) charakteristika jednotlivých jedincov druhu
C) vyžadujú dodatočné podmienky na realizáciu
D) sa nededia
hlavná forma nervová činnosť je reflex. Reflex - kauzálna reakcia tela na zmeny vonkajšieho alebo vnútorného prostredia, vykonávaná s povinnou účasťou centrálnej nervový systém ako odpoveď na stimuláciu receptora. V dôsledku reflexov dochádza k výskytu, zmene alebo zastaveniu akejkoľvek činnosti tela.
Nervová dráha, po ktorej sa šíri vzruch pri realizácii reflexov, sa nazýva reflexný oblúk.
Reflexné oblúky pozostávajú z piatich komponentov: 1) receptor; 2) aferentná nervová dráha; 3) reflexné centrum; 4) eferentná nervová dráha; 5) efektor (pracovný orgán).
Receptor- Ide o citlivé nervové zakončenie, ktoré vníma podráždenie. V receptoroch sa energia stimulu premieňa na energiu nervového impulzu. Rozlišujte: 1) exteroreceptory- vzrušený pod vplyvom podráždenia z prostredia (receptory kože, očí, vnútorné ucho nosová a ústna sliznica); 2) interoreceptory- vnímať podnety z vnútorného prostredia tela (receptory vnútorné orgány, nádoby); 3) proprioreceptory- reagovať na zmenu polohy jednotlivých častí tela v priestore (receptory svalov, šliach, väzov, kĺbových vakov).
Aferentná nervová dráha reprezentované procesmi receptorových neurónov, ktoré prenášajú vzruchy do centrálneho nervového systému.
reflexné centrum pozostáva zo skupiny neurónov umiestnených na rôznych úrovniach centrálneho nervového systému a prenášajúcich nervové impulzy z aferentnej do eferentnej nervovej dráhy.
eferentná nervová dráha vedie nervové impulzy z centrálneho nervového systému do efektora.
Efektor- výkonný orgán, ktorého činnosť sa mení pod vplyvom nervových vzruchov, ktoré k nemu prichádzajú cez útvary reflexného oblúka. Efektormi môžu byť svaly alebo žľazy.
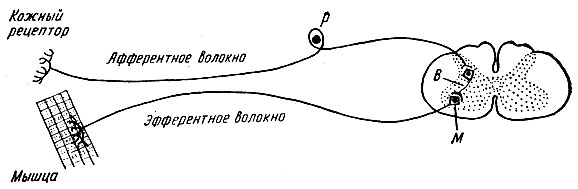
reflexné oblúky môžu byť jednoduché alebo zložité. Jednoduchý reflexný oblúk pozostáva z dvoch neurónov – vnímacieho a efektorového, medzi ktorými je jedna synapsia. Schéma takéhoto dvojneurónového reflexného oblúka je na obr. 71.
Príkladom jednoduchého reflexného oblúka sú reflexné oblúky šľachy, ako je patelárny reflexný oblúk.
Reflexné oblúky väčšiny reflexov nezahŕňajú dva, ale väčší počet neurónov: receptor, jeden alebo viac interkalárnych a efektorových. Takéto reflexné oblúky sa nazývajú komplexné, multineurónové. Schéma komplexného (trojneurónového) reflexného oblúka je na obr. 72.
Teraz sa zistilo, že počas odozvy efektora dochádza k excitácii mnohých nervových zakončení prítomných v pracovnom orgáne. Nervové impulzy teraz z efektora opäť vstupujú do centrálneho nervového systému a informujú ho o správnej reakcii pracovného orgánu. Reflexné oblúky teda nie sú otvorené, ale prstencové útvary.
Reflexy sú veľmi rôznorodé. Môžu byť klasifikované podľa niekoľkých kritérií: 1) biologický význam(potravinové, obranné, sexuálne); 2) v závislosti od typu podráždených receptorov: exteroceptívne, interoceptívne a proprioceptívne; 3) podľa povahy odpovede: motorická alebo motorická (výkonný orgán - sval), sekrečná (efektor - železo), vazomotorická (konstrikcia alebo expanzia krvných ciev).
Všetky reflexy celého organizmu možno rozdeliť do dvoch veľké skupiny: bezpodmienečné a podmienené. Rozdielmi medzi nimi sa budeme zaoberať v kapitole XII.
Činnosť nervového systému má reflexný charakter. Dokonca aj v XVII storočí. Reflexný akt opísal francúzsky filozof a matematik René Descartes. Zaznamenal reakciu tela na podráždenie a navrhol existenciu cesty, po ktorej prechádza nervové vzrušenie. Samotný termín „reflex“ zaviedol neskôr – v 18. storočí – český vedec J. Procházka (z lat. „reflex“ – reflektovaná činnosť). Neskôr I. M. Sechenov vo svojej práci „Reflexy mozgu“ dokázal, že reakcie nervového systému na rôzne druhy podráždenia prebiehajú podľa reflexného mechanizmu, t.j. všetky vedomé a nevedomé akcie sú reflexného pôvodu. Reflexom sa rozumie špecifická reakcia organizmu na dráždivú látku z vnútorného prostredia alebo prijatú zvonka s povinnou účasťou centrálneho nervového systému. Reflexy sa zvyčajne nazývajú funkčné jednotky nervovej aktivity.
Mechanizmy reflexov reflexného oblúka
Reflexy možno klasifikovať podľa rôznych kritérií. Takže v závislosti od úrovne uzavretia oblúka, t.j. podľa umiestnenia reflexného centra sa reflexy delia na spinálne (reflex sa uzatvára v mieche), bulbárne (reflexné centrum - dreň), mezencefalické (štruktúra reflexného oblúka je uzavretá v medzimozgu), diencefalické a kortikálne reflexné centrá sa nachádzajú v. telencephalon a mozgová kôra, resp. Podľa efektorového znaku sú somatické, keď eferentná dráha reflexu zabezpečuje motorickú inerváciu kostrových svalov, a vegetatívne, keď sú efektormi vnútorné orgány. Podľa typu podráždených receptorov sa reflexy delia na exteroceptívne (ak receptor vníma informácie z vonkajšieho prostredia), proprioceptívne (reflexný oblúk vychádza z receptorov pohybového aparátu) a interoceptívne (z receptorov vnútorných orgánov). Interoceptívne reflexy sa zas delia na viscero-viscerálne (reflexný oblúk spája dva vnútorné orgány), visceromaskulárne (receptory sú umiestnené na svalovo-šľachovom aparáte, efektor je vnútorný orgán) a viscero-kutánne (receptory sú lokalizované v koži, pracovných orgánoch – vnútornostiach). Reflexy sa podľa Pavlova delia na podmienené (vyvinuté počas života, špecifické pre každého jedinca) a nepodmienené (vrodené, druhovo špecifické: potravinové, sexuálne, obranno-motorické, homeostatické atď.).
Ako funguje reflexný oblúk?
Bez ohľadu na typ reflexu jeho reflexný oblúk obsahuje receptor, aferentnú dráhu, nervové centrum, eferentnú dráhu, pracovný orgán a spätnú väzbu. Výnimkou sú axónové reflexy, ktorých štruktúra reflexného oblúka sa nachádza v rámci jedného neurónu: senzorické procesy generujú dostredivé impulzy, ktoré sa pri prechode cez telo neurónu šíria pozdĺž axónu do centrálneho nervového systému a pozdĺž vetvy. axónu sa impulzy dostanú k efektoru. Takéto reflexy sa pripisujú fungovaniu metasympatického nervového systému, prostredníctvom ktorých sa vykonávajú napríklad mechanizmy regulácie cievneho tonusu a činnosti kožných žliaz.
Funkciu vnímania podráždenia a jeho premeny na energiu excitácie vykonávajú receptory reflexných oblúkov. Receptorová energia excitácie má charakter lokálnej odozvy, ktorá je dôležitá pri gradácii excitácie silou.
Na základe štruktúry a pôvodu receptorov ich možno rozdeliť na primárne senzorické, sekundárne senzorické a voľné nervové zakončenia. V prvom prípade samotný neurón pôsobí ako receptor (vyvíja sa z neuroepitelu); medzi stimulom a prvým aferentným neurónom nie sú žiadne intermediárne štruktúry. Lokálna odozva primárnych senzorických receptorov - receptorový potenciál - je tiež generátorovým potenciálom, t.j. indukcia akčného potenciálu cez membránu aferentného vlákna. Primárne senzorické receptory zahŕňajú zrakové, čuchové, chemo- a baroreceptory. kardiovaskulárneho systému.
Sekundárne snímacie bunky sú špeciálne štruktúry nenervového pôvodu, ktoré interagujú s dendritmi pseudounipolárnych senzorických buniek pomocou synaptických neuroreceptorových kontaktov. Receptorový potenciál vznikajúci pri pôsobení stimulu v sekundárnych snímacích bunkách nie je generátor a nespôsobuje objavenie sa akčného potenciálu na membráne aferentného vlákna. Excitačný postsynaptický potenciál vzniká len mechanizmom uvoľnenia mediátora receptorovou bunkou. Gradácia sily stimulu sa uskutočňuje vylučovaním rôznych množstiev mediátora (čím viac sa mediátor uvoľňuje, tým silnejší je stimul).
Sekundárne senzorické bunky zahŕňajú sluchové, vestibulárne, karotické, hmatové a iné receptory. Niekedy kvôli zvláštnostiam fungovania do tejto skupiny patria fotoreceptory, ktoré sú z anatomického hľadiska a vzhľadom na svoj pôvod z neuroepitelu sekundárne snímané.
Voľné nervové zakončenia sú rozvetvené dendrity pseudounipolárnych zmyslových buniek a sú lokalizované takmer vo všetkých tkanivách ľudského tela.
Podľa energetickej povahy podnetu, na ktorý receptor reaguje, sa delia na mechanoreceptory (hmatové, baroreceptory, objemové receptory, sluchové, vestibulárne; spravidla vnímajú mechanické dráždenie pomocou bunkových výrastkov), chemoreceptory ( čuchové), cievne chemoreceptory, centrálny nervový systém, fotoreceptory (vnímajú podráždenie cez tyčinkovité a kužeľovité výrastky bunky), termoreceptory (reagujú na zmenu „teplo-chlad“ – Rufiniho telieska a Krauseove banky slizníc) a nociceptory (nezapuzdrené zakončenia bolesti).
Postreceptorové formácie reflexného oblúka
Postreceptorová tvorba štruktúry reflexných oblúkov je aferentná dráha tvorená pseudounipolárnym senzorickým neurónom, ktorého telo leží v spinálny ganglion a tvoria sa axóny zadné korene miecha. Funkciou aferentnej dráhy je viesť informácie do centrálneho spojenia, navyše v tomto štádiu sú informácie zakódované. Na tieto účely sa v tele stavovcov používa binárny kód tvorený výbuchmi (salvami) impulzov a medzier medzi nimi. Existujú dva hlavné typy kódovania: frekvenčné a priestorové.
Prvým je vznik rôzneho počtu impulzov v zhluku, rôzny počet vzplanutí, ich trvanie a trvanie prestávok medzi nimi v závislosti od sily stimulácie aplikovanej na receptor. Priestorové kódovanie vykonáva gradáciu sily stimulu s rôznym počtom nervových vlákien, pozdĺž ktorých sa súčasne vykonáva excitácia.
Aferentná dráha pozostáva hlavne z vlákien A-α, A-β a A-δ.
Po prechode cez vlákna sa nervový impulz dostane do reflexného centra, ktoré je v anatomickom zmysle súborom neurónov umiestnených na určitej úrovni centrálneho nervového systému a podieľajúcich sa na tvorbe tohto reflexu. Funkciou reflexného centra je analyzovať a syntetizovať informácie, ako aj prepínať informácie z aferentnej na eferentnú dráhu.
Funkcie reflexného oblúka
V závislosti od oddelenia nervového systému (somatického a autonómneho) sa reflexy, ktorých centrum sa nachádza v mieche, líšia v lokalizácii interkalárnych neurónov. Takže pre somatický nervový systém je reflexné centrum umiestnené v strednej zóne medzi prednými a zadnými rohmi miechy. Reflexné centrum autonómneho nervového systému (telo interkalárnych neurónov) leží v zadné rohy. Somatické a autonómne časti nervového systému sa líšia aj lokalizáciou eferentných neurónov. Telá motorických neurónov somatického nervového systému ležia v predných rohoch miechy, telá pregangliových neurónov autonómny systém- na úrovni stredných rohov.
Axóny oboch typov buniek tvoria eferentnú dráhu reflexného oblúka. V somatickom nervovom systéme je súvislý, tvoria ho vlákna typu A-α. Výnimkou sú len vlákna A-γ, ktoré vedú vzruch z buniek miechy do intrafúznych vlákien svalových vretien. Eferentná dráha autonómneho nervového systému je prerušená v autonómny ganglion lokalizované alebo intramurálne (pár sympatická časť), alebo v blízkosti miechy (samotnej alebo v sympatický kmeň- sympatická časť). Pregangliové vlákno patrí do B-vlákna, postgangliové vlákno do C skupiny.
Pracovným orgánom pre somatickú časť nervového systému je priečne pruhovaný kostrový sval, vo vegetatívnom oblúku je efektorom žľaza alebo sval (hladký alebo priečne pruhovaný srdcový). Medzi eferentným spôsobom a pracovným orgánom je chemická myoneurálna alebo neurosekrečná synapsia.
Reflexný oblúk sa uzatvára do prstenca v dôsledku reverznej aferentácie - toku impulzov z efektorových receptorov späť do reflexného centra. Funkcia spätnej väzby - signalizácia centrálnemu nervovému systému o vykonanej akcii. Ak sa nevykonáva dostatočne, nervové centrum je vzrušené - reflex pokračuje. Vďaka reverznej aferentácii sa tiež vykonáva kontrola periférnej aktivity centrálneho nervového systému.
Rozlišujte medzi negatívnou a pozitívnou spätnou väzbou. Prvý, pri vykonávaní určitej funkcie, spustí mechanizmus, ktorý túto funkciu inhibuje. Pozitívna spätná väzba pozostáva z ďalšej stimulácie funkcie, ktorá sa už vykonáva, alebo inhibície funkcie, ktorá je už potlačená. Pozitívna reverzná aferentácia je zriedkavá, pretože uvádza biologický systém do nestabilnej polohy.
Jednoduché (monosynaptické) reflexné oblúky pozostávajú iba z dvoch neurónov (aferentného a eferentného) a líšia sa iba proprioceptívnymi reflexmi. Zostávajúce oblúky zahŕňajú všetky vyššie uvedené komponenty.
Reflexný oblúk je reťazec neurónov od periférneho receptora cez centrálny nervový systém k periférnemu efektoru. Prvky reflexného oblúka sú periférny receptor, aferentná dráha, jeden alebo viac interneurónov, eferentná dráha a efektor.
Všetky receptory sú zapojené do určitých reflexov, takže ich aferentné vlákna slúžia ako aferentná dráha zodpovedajúceho reflexného oblúka. Počet interneurónov je vždy väčší ako jeden, s výnimkou monosynaptického strečového reflexu. Eferentná dráha je reprezentovaná buď motorickými axónmi alebo postgangliovými vláknami autonómneho nervového systému a efektormi sú kostrové svaly a hladké svaly, srdce a žľazy.
Čas od začiatku podnetu do odozvy efektora sa nazýva reflexný čas. Vo väčšine prípadov je určený najmä časom vedenia v aferentných a eferentných dráhach a v centrálnej časti reflexného oblúka, ku ktorým treba pripočítať čas premeny vzruchu v receptore na šíriaci sa impulz, čas prenosu cez synapsie v centrálnom nervovom systéme (synaptické oneskorenie), čas prenosu z eferentnej dráhy do efektora a čas aktivácie efektora.
Reflexné oblúky sú rozdelené do niekoľkých typov
1. Monosynaptické reflexné oblúky - na takomto oblúku sa podieľa len jedna synapsia, nachádzajúca sa v centrálnom nervovom systéme. Takéto reflexy sú veľmi bežné u všetkých stavovcov, podieľajú sa na regulácii svalový tonus a držanie tela (napr. trhnutie kolenom). V týchto oblúkoch neuróny nedosahujú mozog a reflexné činy sa vykonávajú bez jeho účasti, pretože sú stereotypné a nevyžadujú myslenie alebo vedomé rozhodnutie. Sú ekonomické, pokiaľ ide o počet zapojených centrálnych neurónov a zbavujú sa zásahu mozgu.
2. Polysynaptické miechové reflexné oblúky - zahŕňajú najmenej dve synapsie umiestnené v centrálnom nervovom systéme, pretože v oblúku je zahrnutý tretí neurón - interkalárny alebo intermediárny neurón. Tu sú synapsie medzi senzorickým neurónom a interneurónom a medzi interkalárnymi a motorickými neurónmi. Takéto reflexné oblúky umožňujú telu vykonávať automatické mimovoľné reakcie potrebné na prispôsobenie sa zmenám vonkajšieho prostredia (napr. pupilárny reflex alebo udržiavanie rovnováhy pri pohybe) a na zmeny v tele samotnom (regulácia dychovej frekvencie, krvného tlaku a pod.).
3. Polysynaptické reflexné oblúky zahŕňajúce miechu aj mozog – pri tomto type reflexných oblúkov existuje v mieche synapsia medzi senzorickým neurónom a neurónom, ktorá vysiela impulzy do mozgu.
Reflexy možno klasifikovať podľa rôznych kritérií. Takže v závislosti od úrovne uzavretia oblúka, t.j. podľa umiestnenia reflexného centra sa reflexy delia na miechové (reflex sa uzatvára v mieche), bulbárne (centrom reflexu je predĺžená miecha), mezencefalické (reflexný oblúk je uzavretý v strednom mozgu), diencefalické a kortikálne reflexné centrá sa nachádzajú v telencefalu a kôre veľkých hemisfér, resp.
Podľa efektorového znaku sú somatické, keď eferentná dráha reflexu zabezpečuje motorickú inerváciu kostrových svalov, a vegetatívne, keď sú efektormi vnútorné orgány.
Podľa typu podráždených receptorov sa reflexy delia na exteroceptívne (ak receptor vníma informácie z vonkajšieho prostredia), proprioceptívne (reflexný oblúk vychádza z receptorov pohybového aparátu) a interoceptívne (z receptorov vnútorných orgánov).
Interoceptívne reflexy sa zas delia na viscero-viscerálne (reflexný oblúk spája dva vnútorné orgány), visceromaskulárne (receptory sú umiestnené na svalovo-šľachovom aparáte, efektor je vnútorný orgán) a viscero-kutánne (receptory sú lokalizované v koži, pracovných orgánoch – vnútornostiach).
Reflexy sa podľa Pavlova delia na podmienené (vyvinuté počas života, špecifické pre každého jedinca) a nepodmienené (vrodené, druhovo špecifické: potravinové, sexuálne, obranno-motorické, homeostatické atď.).
Bez ohľadu na typ reflexu jeho reflexný oblúk obsahuje receptor, aferentnú dráhu, nervové centrum, eferentnú dráhu, pracovný orgán a spätnú väzbu. Výnimkou sú axónové reflexy, ktorých reflexný oblúk sa nachádza v rámci jedného neurónu: senzorické procesy generujú dostredivé impulzy, ktoré sa pri prechode cez telo neurónu šíria pozdĺž axónu do centrálneho nervového systému a pozdĺž vetvy axónu. , impulzy dosiahnu efektor. Takéto reflexy sa pripisujú fungovaniu metasympatického nervového systému, prostredníctvom ktorých sa vykonávajú napríklad mechanizmy regulácie cievneho tonusu a činnosti kožných žliaz.
Funkciu vnímania podráždenia a jeho premeny na energiu excitácie vykonávajú receptory reflexných oblúkov. Receptorová energia excitácie má charakter lokálnej odozvy, ktorá je dôležitá pri gradácii excitácie silou.
Na základe štruktúry a pôvodu receptorov ich možno rozdeliť na primárne senzorické, sekundárne senzorické a voľné nervové zakončenia. V prvom prípade samotný neurón pôsobí ako receptor (vyvíja sa z neuroepitelu); medzi stimulom a prvým aferentným neurónom nie sú žiadne intermediárne štruktúry. Lokálna odozva primárnych senzorických receptorov - receptorový potenciál - je tiež generátorovým potenciálom, t.j. indukcia akčného potenciálu cez membránu aferentného vlákna. Primárne senzorické receptory zahŕňajú zrakové, čuchové, chemo- a baroreceptory kardiovaskulárneho systému.
Sekundárne snímacie bunky sú špeciálne štruktúry nenervového pôvodu, ktoré interagujú s dendritmi pseudounipolárnych senzorických buniek pomocou synaptických neuroreceptorových kontaktov. Receptorový potenciál vznikajúci pri pôsobení stimulu v sekundárnych snímacích bunkách nie je generátorom a nespôsobuje objavenie sa akčného potenciálu na membráne aferentného vlákna. Excitačný postsynaptický potenciál vzniká len mechanizmom uvoľnenia mediátora receptorovou bunkou. Gradácia sily stimulu sa uskutočňuje vylučovaním rôznych množstiev mediátora (čím viac sa mediátor uvoľňuje, tým silnejší je stimul).
Sekundárne senzorické bunky zahŕňajú sluchové, vestibulárne, karotické, hmatové a iné receptory. Niekedy kvôli zvláštnostiam fungovania do tejto skupiny patria fotoreceptory, ktoré sú z anatomického hľadiska a vzhľadom na svoj pôvod z neuroepitelu sekundárne snímané.
Voľné nervové zakončenia sú rozvetvené dendrity pseudounipolárnych zmyslových buniek a sú lokalizované takmer vo všetkých tkanivách ľudského tela.
Podľa energetickej povahy podnetu, na ktorý receptor reaguje, sa delia na mechanoreceptory (hmatové, baroreceptory, volumoreceptory, sluchové, vestibulárne; spravidla vnímajú mechanické dráždenie pomocou bunkových výrastkov), chemoreceptory (čuchové ), chemoreceptory krvných ciev, centrálny nervový systém, fotoreceptory (vnímajú podráždenie cez tyčinkovité a kužeľovité výrastky bunky), termoreceptory (reagujú na zmenu „teplo-chlad“ – Rufiniho telieska a Krauseove banky slizníc) a nociceptory (nezapuzdrené zakončenie bolesti).
Postreceptorová tvorba reflexných oblúkov je aferentná dráha tvorená pseudounipolárnym senzorickým neurónom, ktorého telo leží v miechovom gangliu a axóny tvoria zadné korene miechy. Funkciou aferentnej dráhy je viesť informácie do centrálneho spojenia, navyše v tomto štádiu sú informácie zakódované. Na tieto účely sa v tele stavovcov používa binárny kód tvorený výbuchmi (salvami) impulzov a medzier medzi nimi. Existujú dva hlavné typy kódovania: frekvenčné a priestorové.
Prvým je vznik rôzneho počtu impulzov v zhluku, rôzny počet vzplanutí, ich trvanie a trvanie prestávok medzi nimi v závislosti od sily stimulácie aplikovanej na receptor. Priestorové kódovanie vykonáva gradáciu sily stimulu s rôznym počtom nervových vlákien, pozdĺž ktorých sa súčasne vykonáva excitácia.
Zloženie aferentnej dráhy zahŕňa najmä vlákna A-b, A-c a A-d.
Po prechode cez vlákna sa nervový impulz dostane do reflexného centra, ktoré je v anatomickom zmysle súborom neurónov umiestnených na určitej úrovni centrálneho nervového systému a podieľajúcich sa na tvorbe tohto reflexu. Funkciou reflexného centra je analyzovať a syntetizovať informácie, ako aj prepínať informácie z aferentnej na eferentnú dráhu.
V závislosti od oddelenia nervového systému (somatického a autonómneho) sa reflexy, ktorých centrum sa nachádza v mieche, líšia v lokalizácii interkalárnych neurónov. Takže pre somatický nervový systém je reflexné centrum umiestnené v strednej zóne medzi prednými a zadnými rohmi miechy. Reflexné centrum autonómneho nervového systému (telá interkalárnych neurónov) leží v zadných rohoch. Somatické a autonómne časti nervového systému sa líšia aj lokalizáciou eferentných neurónov. Telá motorických neurónov somatického nervového systému ležia v predných rohoch miechy, telá pregangliových neurónov autonómneho systému ležia na úrovni stredných rohov.
Axóny oboch typov buniek tvoria eferentnú dráhu reflexného oblúka. V somatickom nervovom systéme je súvislý, tvoria ho vlákna typ A-b. Výnimkou sú len vlákna A-g, ktoré vedú vzruch z buniek miechy do intrafúznych vlákien svalových vretien. Eferentná dráha autonómneho nervového systému je prerušená v autonómnom gangliu, lokalizovanom buď intramurálne (parasympatická časť) alebo v blízkosti miechy (samostatne alebo v sympatickom kmeni - sympatická časť). Pregangliové vlákno patrí do B-vlákna, postgangliové vlákno patrí do skupiny C.
Pracovným orgánom pre somatickú časť nervového systému je priečne pruhovaný kostrový sval, vo vegetatívnom oblúku je efektorom žľaza alebo sval (hladký alebo priečne pruhovaný srdcový). Medzi eferentnou dráhou a pracovným orgánom je chemická myoneurálna alebo neurosekrečná synapsia.
Reflexný oblúk sa prstencovo uzatvára v dôsledku reverznej aferentácie – toku impulzov z efektorových receptorov späť do reflexného centra. Funkcia spätnej väzby - signalizácia centrálnemu nervovému systému o vykonanej akcii. Ak sa nevykonáva dostatočne, nervové centrum je vzrušené - reflex pokračuje. Vďaka reverznej aferentácii sa tiež vykonáva kontrola periférnej aktivity centrálneho nervového systému.
Rozlišujte medzi negatívnou a pozitívnou spätnou väzbou. Prvý, pri vykonávaní určitej funkcie, spustí mechanizmus, ktorý túto funkciu inhibuje. Pozitívna spätná väzba pozostáva z ďalšej stimulácie funkcie, ktorá sa už vykonáva, alebo inhibície funkcie, ktorá je už potlačená. Pozitívna reverzná aferentácia je zriedkavá, pretože uvádza biologický systém do nestabilnej polohy.
Jednoduché (monosynaptické) reflexné oblúky pozostávajú iba z dvoch neurónov (aferentného a eferentného) a líšia sa iba proprioceptívnymi reflexmi. Zostávajúce oblúky zahŕňajú všetky vyššie uvedené komponenty.
Fyziologické vlastnosti a funkčný význam nervových vlákien
Nervové vlákna majú najvyššiu excitabilitu, najvyššiu rýchlosť vedenia vzruchu, najkratšiu refraktérnu periódu a vysokú labilitu. Toto je poskytnuté vysoký stupeň metabolické procesy a nízky membránový potenciál.
Funkcia: vedenie nervových impulzov z receptorov do centrálneho nervového systému a naopak.
Štrukturálne vlastnosti a typy nervových vlákien
Nervové vlákno - axón - je pokryté bunkovou membránou.
Existujú 2 typy nervových vlákien:
Nemyelinizované nervové vlákna - jedna vrstva Schwannových buniek, medzi nimi - štrbinovité priestory. bunková membrána počas celého kontaktu s životné prostredie. Pri aplikácii podráždenia dochádza k excitácii v mieste pôsobenia stimulu. Nemyelinizované nervové vlákna majú elektrogénne vlastnosti (schopnosť vytvárať nervové impulzy).
Myelinizované nervové vlákna - pokryté vrstvami Schwannových buniek, ktoré miestami tvoria uzliny Ranviera (oblasti bez myelínu) každý 1 mm. Trvanie zachytenia Ranviera je 1 µm. Myelínový obal plní trofické a izolačné funkcie (vysoká odolnosť). Oblasti pokryté myelínom nemajú elektrogénne vlastnosti. Majú zásahy Ranviera. K excitácii dochádza pri zachytení Ranviera najbližšie k miestu pôsobenia stimulu. Pri zákrokoch Ranviera vysoká hustota Na-kanály preto pri každom zachytení Ranviera dochádza k zvýšeniu nervových impulzov.
Ranvierove zachytenia fungujú ako opakovače (generujú a zosilňujú nervové impulzy).
Mechanizmus vedenia vzruchu pozdĺž nervového vlákna
1885 - L. German - medzi excitovanými a nevybudenými úsekmi nervového vlákna vznikajú kruhové prúdy.
Pri pôsobení dráždidla existuje potenciálny rozdiel medzi vonkajším a vnútorným povrchom tkaniva (oblasti, ktoré nesú rôzne náboje). Medzi týmito oblasťami vzniká elektrický prúd (pohyb iónov Na +). Vo vnútri nervového vlákna vzniká prúd od kladného pólu k zápornému pólu, to znamená, že prúd smeruje z excitovanej oblasti do nevybudenej. Tento prúd vystupuje cez nevybudenú oblasť a spôsobuje jej opätovné nabitie. Na vonkajšom povrchu nervového vlákna prúd tečie z nevzrušenej oblasti do oblasti vzrušenej. Tento prúd nemení stav excitovanej oblasti, pretože je v stave žiaruvzdornosti.
Dôkaz o prítomnosti kruhových prúdov: nervové vlákno je umiestnené v roztok NaCl a registrujte rýchlosť budenia. Potom sa nervové vlákno umiestni do oleja (zvyšuje sa odpor) - rýchlosť vedenia sa zníži o 30%. Potom je nervové vlákno ponechané vo vzduchu - rýchlosť excitácie sa zníži o 50%.
Vlastnosti vedenia vzruchu pozdĺž myelinizovaných a nemyelinizovaných nervových vlákien:
myelínové vlákna - majú plášť s vysokou odolnosťou, elektrogénne vlastnosti iba v uzloch Ranvier. Pri pôsobení stimulu dochádza k excitácii v najbližšom zábere Ranviera. Odpočúvanie susedov v stave polarizácie. Výsledný prúd spôsobí depolarizáciu susedného záchytu. Ranvierove uzly majú vysokú hustotu Na-kanálov, preto v každom ďalšom uzle vzniká o niečo väčší (amplitúdovo) akčný potenciál, vďaka čomu sa excitácia šíri bez úbytku a môže preskočiť cez niekoľko uzlov. Toto je Tasakiho slaná teória. Dôkazom teórie je, že do nervového vlákna boli injikované lieky, ktoré blokujú niekoľko zachytení, ale potom bolo zaznamenané vedenie vzruchu. Ide o vysoko spoľahlivú a ziskovú metódu, pretože sa eliminuje menšie poškodenie, zvyšuje sa rýchlosť budenia a znižujú sa náklady na energiu;
nemyelinizované vlákna - povrch má v celom rozsahu elektrogénne vlastnosti. Preto sa malé kruhové prúdy vyskytujú vo vzdialenosti niekoľkých mikrometrov. Vzruch má formu neustále sa pohybujúcej vlny.
Táto metóda je menej zisková: vysoké náklady na energiu (na prevádzku čerpadla Na-K), nižšia miera budenia.
Klasifikácia nervových vlákien
Nervové vlákna sú klasifikované podľa:
trvanie akčného potenciálu;
štruktúra (priemer) vlákna;
rýchlosť budenia.
Rozlišujú sa tieto skupiny nervových vlákien:
skupina A (alfa, beta, gama, delta) - najkratší akčný potenciál, najhrubší myelínový obal, najvyššia rýchlosť excitácie;
skupina B - myelínová pošva je menej výrazná;
Skupina C - bez myelínového obalu.
Morfologické rozdiely medzi dendritmi a axónmi
1. Jednotlivý neurón má niekoľko dendritov, axón je vždy jeden.
2. Dendrity sú vždy kratšie ako axón. Ak veľkosť dendritov nepresahuje 1,5-2 mm, potom môžu axóny dosiahnuť 1 m alebo viac.
3. Dendrity sa plynule vzďaľujú od tela bunky a postupne majú konštantný priemer na značnú vzdialenosť.
4. Dendrity sa zvyčajne rozvetvujú pod ostrý uhol a vetvy sú nasmerované preč z bunky. Axóny vydávajú kolaterály najčastejšie v pravom uhle, orientácia kolaterál priamo nesúvisí s polohou bunkového tela.
5. Vzor dendritického vetvenia v bunkách rovnakého typu je konštantnejší ako vetvenie axónu týchto buniek.
6. Dendrity zrelých neurónov sú pokryté dendritickými tŕňmi, ktoré chýbajú na sóme a počiatočnej časti dendritických kmeňov. Axóny nemajú tŕne.
7. Dendrity nikdy nemajú kašovitú škrupinu. Axóny sú často obklopené myelínom.
8. Dendrity majú pravidelnejšiu priestorovú organizáciu mikrotubulov, v axónoch dominujú neurofilamenty a mikrotubuly sú menej usporiadané
9. V dendritoch, najmä v ich proximálnych častiach, sú endoplazmatické retikulum a ribozómy, ktoré nie sú v axónoch.
10. Povrch dendritov je vo väčšine prípadov v kontakte so synoptickými plakmi a má aktívne zóny s postsynaptickou špecializáciou.
Štruktúra dendritov
Ak existuje pomerne rozsiahla literatúra o geometrii dendritov, dĺžke ich vetiev, orientácii, tak o vnútorná štruktúra, o štruktúre jednotlivých zložiek ich cytoplazmy existujú len samostatné rozptýlené informácie. Tieto informácie boli možné až po zavedení elektrónových mikroskopických štúdií do neurohistológie.
Hlavné charakteristické znaky dendritu, ktoré ho odlišujú na elektrónových mikroskopických rezoch:
1) nedostatok myelínového obalu,
prítomnosť správneho systému mikrotubulov,
3) prítomnosť aktívnych zón synapsií na nich s jasne vyjadrenou elektrónovou hustotou cytoplazmy dendritu,
4) odklon od spoločného kmeňa dendritov tŕňov,
5) špeciálne organizované zóny pobočkových uzlov,
6) zahrnutie ribozómov,
7) prítomnosť granulárneho a negranulárneho endoplazmatického retikula v proximálnych oblastiach.
Najpozoruhodnejším znakom dendritickej cytoplazmy je prítomnosť početných mikrotubulov. Sú dobre identifikovateľné v priečnych aj pozdĺžnych rezoch. Vychádzajúc z proximálnej časti dendritu prebiehajú mikrotubuly rovnobežne s dlhou osou dendritu až po jeho distálne vetvy. Mikrotubuly nasledujú v dendrite navzájom paralelne, bez toho, aby sa navzájom spájali alebo pretínali. Na prierezoch je vidieť, že vzdialenosti medzi jednotlivými tubulmi sú konštantné. Jednotlivé dendritické tubuly sa rozprestierajú na pomerne dlhé vzdialenosti, často sledujú krivky, ktoré môžu byť pozdĺž priebehu dendritov. Počet tubulov je relatívne konštantný na jednotku plochy prierezu dendritu a je približne 100 na 1 µm. Toto číslo je typické pre všetky dendrity odobraté rôzne oddelenia centrálny a periférny nervový systém, odlišné typy zvierat.
Funkciou mikrotubulov je transport látok pozdĺž procesov nervových buniek.
Keď sú mikrotubuly zničené, transport látok v dendrite môže byť narušený, a tým sú terminálne úseky procesov zbavené prítoku živín a energetické látky z bunkového tela. Dendrity, aby sa zachovala štruktúra synaptických kontaktov v extrémnych podmienkach a tým zabezpečila funkcia interneuronálnej interakcie, kompenzujú deficit živiny v dôsledku štruktúr, ktoré k nim priliehajú (synaptické plaky, viacvrstvové myelínové puzdro mäkkých vlákien, ako aj fragmenty gliových buniek).
Ak je pôsobenie patogénneho faktora eliminované včas, dendrity obnovujú štruktúru a správnu priestorovú organizáciu mikrotubulov, čím obnovujú systém transportu látok, ktorý je vlastný normálnemu mozgu. Ak je sila a trvanie patogénneho faktora významné, potom sa javy endocytózy namiesto ich adaptívnej funkcie môžu stať pre dendrity smrteľnými, pretože fagocytované fragmenty sa nedajú využiť a nahromadenie v cytoplazme dendritov povedie k ich nezvratnému poškodenie.
Porušenie v organizácii mikrotubulov vedie k prudkej zmene v správaní zvierat. U zvierat, u ktorých boli v experimente zničené mikrotubuly v dendritoch, bola pozorovaná dezorganizácia zložitých foriem správania, pričom sa zachovali jednoduché podmienené reflexy. U ľudí to môže viesť k vážnym poruchám vyššej nervovej aktivity.
Skutočnosť, že dendrity sú najcitlivejším miestom na pôsobenie patologického agens v duševná choroba, svedčia niektoré práce amerických vedcov. Ukázalo sa, že pri senilnej demencii (cyanotická demencia) a Alzheimerovej chorobe mozgové preparáty spracované Golgiho metódou neodhalia procesy nervových buniek. Kmene dendritov sa zdajú byť spálené a zuhoľnatené. Nedetekcia týchto procesov na histologických preparátoch mozgu je pravdepodobne spojená aj s porušením systému mikrotubulov a neurofilament v týchto procesoch.
Nájdené v dendritoch. Nasledujú rovnobežne s dlhou osou dendritu, môžu ležať oddelene alebo byť zhromaždené vo zväzkoch, ale nie sú striktne umiestnené v cytoplazme. Pravdepodobne spolu s mikrotubulmi môžu byť ekvivalentom neurofibríl.
Všetky dendrity CNS sú charakterizované zväčšením povrchu v dôsledku viacnásobného dichotomického delenia. V tomto prípade sa v deliacich zónach vytvárajú špeciálne expanzné miesta alebo uzly odbočiek.
Normálna analýza ukazuje, že v uzle vetvy, ku ktorému sa približujú dve dendritické vetvy, z ktorých každá nesie svoj vlastný signál, možno vykonať nasledujúce operácie. Cez uzol vetvy spoločný kmeň a ďalej do tela neurónu prechádzajú:
alebo signál z jednej pobočky,
alebo len z inej
alebo výsledok interakcie dvoch signálov,
alebo sa signály navzájom rušia.
Cytoplazma uzla vetvy obsahuje takmer všetky zložky, ktoré sú charakteristické pre telo nervová bunka a miesta sa svojou štruktúrou výrazne líšia od cytoplazmy spoločného dendritického kmeňa a vetiev získaných delením. Rozvetvené uzliny obsahujú zvýšený počet mitochondrií, viditeľné sú zrnité a hladké retikulum, zhluky jednotlivých ribozómov a ribozómy zostavené do roziet. Tieto zložky (granulárne a hladké retikulum, ribozómy) sa priamo podieľajú na syntéze bielkovín. Hromadenie mitochondrií v týchto miestach naznačuje intenzitu oxidačných procesov.
Funkcie dendritov
Chcel by som poznamenať, že hlavnými ťažkosťami, s ktorými sa výskumník stretáva pri štúdiu funkcie dendritov, je nedostatok informácií o vlastnostiach dendritovej membrány (na rozdiel od membrány tela neurónu) v dôsledku nemožnosti zavedenia mikroelektródy. do dendritu.
Pri hodnotení celkovej geometrie dendritov, rozmiestnenia synapsií a špeciálnej štruktúry cytoplazmy v miestach dendritického vetvenia možno hovoriť o špeciálnych neurónových lokusoch s vlastnou funkciou. Najjednoduchšia vec, ktorú možno pripísať dendritickým miestam na miestach vetvenia, je trofická funkcia.
Z vyššie uvedeného vyplýva, že cytoplazma dendritov obsahuje veľa ultra konštrukčné komponenty schopné plniť svoje dôležité funkcie. V dendrite sú určité lokusy, ktorých práca má svoje vlastné charakteristiky.
Hlavným účelom početných dendritických vetiev nervovej bunky je poskytnúť prepojenie s inými neurónmi. v mozgovej kôre cicavcov veľký podiel axodendrické spojenia spadajú na kontakty so špeciálnymi špecializovanými výrastkami dendritov - dendritických tŕňov. Dendritické tŕne sú fylogeneticky najmladšími útvarmi v nervovom systéme. V ontogenéze dozrievajú oveľa neskôr ako ostatné nervové štruktúry a predstavujú najplastickejší aparát nervovej bunky.
Dendritická chrbtica má spravidla charakteristický tvar v mozgovej kôre cicavcov. (obr. 2). Z hlavného dendritického kmeňa vystupuje pomerne úzka stopka, ktorá končí predĺžením - hlavou. Je pravdepodobné, že táto forma dendritického prívesku (prítomnosť hlavy) je spojená na jednej strane so zväčšením plochy synaptického kontaktu s axónovým zakončením a na druhej strane slúži na umiestňujú špecializované organely vo vnútri chrbtice, najmä ostnatý aparát, ktorý je prítomný iba v dendritických tŕňoch mozgovej kôry cicavcov. V tomto smere sa javí ako vhodná analógia s tvarom zakončenia synaptického axónu, keď tenké predterminálne vlákno tvorí predĺženie. Táto expanzia (synaptický plak) vytvára rozsiahly kontakt s inervovaným substrátom a obsahuje vo vnútri veľký súbor ultraštrukturálnych komponentov (synaptické vezikuly, mitochondrie, neurofilamenty, glykogénové granuly).
Existuje hypotéza (ktorú zdieľa a rozvíja najmä nositeľ Nobelovej ceny F. Crick), že geometria chrbtice sa môže meniť v závislosti od funkčného stavu mozgu. V tomto prípade sa úzky krčok chrbtice môže roztiahnuť a chrbtica samotná sa splošťuje, čo vedie k zvýšeniu účinnosti axo-chrbticového kontaktu.
Ak sa tvar a veľkosť dendritických tŕňov v mozgovej kôre cicavcov môže trochu líšiť, potom je v nich najstálejšia prítomnosť špecifického chrbticového aparátu. Ide o komplex vzájomne prepojených tubulov (cisterny) umiestnených spravidla v hlave chrbtice. Pravdepodobne je táto organela spojená s veľmi dôležitými funkciami, ktoré sú vlastné fylogeneticky najmladším mozgovým formáciám, pretože ostnatý aparát sa nachádza hlavne v mozgovej kôre a len u vyšších živočíchov.
Napriek všetkému je chrbtica derivátom dendritu, chýbajú jej neurofilamenty a dendritické tubuly, jej cytoplazma obsahuje hrubo alebo jemne zrnitú matricu. Ešte jeden vlastnosť chrbtice v mozgovej kôre je povinná prítomnosť synaptických kontaktov s zakončeniami axónov na nich. Cytoplazma chrbtice má špeciálne zložky, ktoré ju odlišujú od dendritických stoniek. V cytoplazme chrbtice je možné zaznamenať zvláštnu triádu: subsynaptická špecializácia aktívnych zón - ostnatý aparát - mitochondrie. Vzhľadom na rôznorodosť zložitých a dôležitých funkcií, ktoré mitochondrie vykonávajú, možno pri synaptickom prenose očakávať aj komplexné funkčné prejavy v „triádach“. Dá sa povedať, že cytoplazma dendritickej chrbtice a ostnatého aparátu môžu priamo súvisieť so synaptickou funkciou.
Dendritické tŕne a konce dendritov sú tiež veľmi citlivé na extrémne faktory. Pri akomkoľvek type otravy (napríklad alkoholická, hypoxická, ťažké kovy - olovo, ortuť atď.) sa mení počet tŕňov na dendritoch buniek mozgovej kôry. S najväčšou pravdepodobnosťou ostne nezmiznú, ale ich cytoplazmatické zložky sú narušené a horšie sú impregnované soľami. ťažké kovy. Keďže chrbtice sú jednou zo štrukturálnych zložiek interneuronálnych kontaktov, ich poruchy vedú k vážnemu narušeniu funkcie mozgu.
V niektorých prípadoch pri krátkodobom pôsobení extrémneho faktora môže na prvý pohľad nastať paradorzálna situácia, kedy sa počet ostňov nájdených na dendritoch mozgových buniek neznižuje, ale zvyšuje. Toto sa pozoruje počas experimentálnej cerebrálnej ischémie v jej počiatočnom období. Paralelne s nárastom počtu identifikovaných tŕňov, funkčný stav mozog. V tomto prípade je hypoxia faktorom, ktorý prispieva k zvýšenému metabolizmu v nervovom tkanive, lepšej implementácii rezerv, ktoré nie sú využité v normálne prostredie, a rýchle spaľovanie toxínov nahromadených v tele. Ultraštrukturálne sa to prejavuje intenzívnejším štúdiom cytoplazmy tŕňov, rastom a zväčšením cisterien chrbtového aparátu. Pravdepodobne je tento jav pozitívneho účinku hypoxie pozorovaný, keď človek zažíva veľké fyzické cvičenie v podmienkach hypoxie dobýva horské vrcholy. Tieto ťažkosti sú potom kompenzované intenzívnejšou produktívnou prácou, a to ako mozgu, tak aj iných orgánov.
Tvorba dendritov
Dendrity a ich interneuronálne spojenia vznikajú počas ontogenetického vývoja mozgu. Navyše, dendrity, najmä apikálne, u mladých jedincov zostávajú po určitú dobu voľné na vytváranie nových kontaktov. Časti dendritu umiestnené bližšie k telu bunky sú pravdepodobne spojené so silnejšími a jednoduchšími prirodzenými podmienenými reflexmi a konce sú ponechané na vytváranie nových spojení a asociácií.
V dospelosti už na dendritoch nie sú oblasti bez interneuronálnych kontaktov, ale starnutím sú to práve konce dendritov, ktoré ako prvé trpia a z hľadiska nasýtenia kontaktmi
u starých jedincov pripomínajú dendrity detstva. K tomu dochádza v dôsledku oslabenia procesov syntetizujúcich transportné proteíny v bunke a v dôsledku zhoršeného zásobovania mozgu krvou. Možno práve tu leží morfologický základ pre takéto známe v neurológii a in každodenný život skutočnosť, že keď sa starí ľudia ťažko učia niečo nové, často zabúdajú na aktuálne udalosti a veľmi dobre si pamätajú minulosť. To isté sa pozoruje v prípade otravy.
Ako už bolo uvedené, nárast a komplikácie dendritického stromu vo fylogenéze sú potrebné nielen pre vnímanie Vysoké číslo prichádzajúcich impulzov, ale aj na predspracovanie.
Dendrity neurónov centrálneho nervového systému majú v celom rozsahu synaptickú funkciu a koncové úseky nie sú v tomto nijako horšie ako stredné. Ak rozprávame sa o distálnych (terminálnych) úsekoch apikálnych dendritov pyramidálne neuróny mozgovej kôry, potom je ich podiel na realizácii interneuronálnych interakcií ešte výraznejší ako proximálnych. Tam na viac terminálne synaptické plaky na samotnom trupe a na vetvách apikálneho dendritu sú spojené viacerými kontaktmi na dendritických tŕňoch.
Štúdiom tohto problému pomocou elektrónovej mikroskopie sa vedci tiež presvedčili, že koncové časti dendritov sú husto pokryté synaptickými plakmi, a teda sú priamo zapojené do interneuronálnych interakcií. elektrónová mikroskopia tiež ukázali, že dendrity môžu vytvárať vzájomné kontakty. Tieto kontakty môžu byť buď paralelné, čomu väčšina autorov pripisuje elektrotonické vlastnosti, alebo typické asymetrické synapsie s dobre definovanými organelami, ktoré zabezpečujú chemický prenos. Takéto dendro-dendritické kontakty ešte len začínajú priťahovať pozornosť výskumníkov. Dendrit teda po celej svojej dĺžke plní synaptickú funkciu. Ako je povrch dendritu prispôsobený tak, aby poskytoval kontakty s koncami axónov?
Povrchová membrána dendritu je navrhnutá tak, aby bola maximálne využitá pre interneuronálne kontakty. Celý dendrit je jamkovaný s priehlbinami, záhybmi, vreckami, má rôzne nepravidelnosti typu mikrovýrastkov, hrotov, hríbovitých príveskov atď. Všetky tieto reliéfy dendritických kmeňov zodpovedajú tvaru a veľkosti vstupujúcich synaptických zakončení. A v rôzne oddelenia nervového systému a u rôznych zvierat má reliéf dendritického povrchu špecifické črty. Samozrejme, najpozoruhodnejším výrastkom dendritickej membrány je dendritická chrbtica.
Dendrity sú veľmi citlivé na pôsobenie rôznych extrémnych faktorov. Porušenie v nich vedie k mnohým chorobám, ako sú duševné poruchy.
Reflexný oblúk pozostáva z:
- receptory - vnímanie podráždenia.
- senzitívne (centripetálne, aferentné) nervové vlákno, ktoré prenáša vzruch do centra
- nervové centrum, kde dochádza k prepínaniu vzruchu zo senzorických neurónov na motorické neuróny
- motorické (odstredivé, eferentné) nervové vlákno, ktoré prenáša vzruchy z centrálneho nervového systému do pracovného orgánu
- efektor - pracovný orgán, ktorý vykonáva účinok, reakciu na podráždenie receptora.
Receptory a receptívne polia
Receptor- podráždenie vnímajúce bunky.
receptívne pole- to je anatomická oblasť, pri podráždení vzniká tento reflex.
Recepčné polia primárnych senzorických receptorov sú organizované najjednoduchším spôsobom. Napríklad taktilné alebo nociceptívne receptívne pole povrchu kože je rozvetvením jediného senzorického vlákna.
Receptory umiestnené v rôznych častiach receptívneho poľa majú rôznu citlivosť na adekvátnu stimuláciu. Vysoko citlivá zóna sa zvyčajne nachádza v strede receptívneho poľa a citlivosť klesá bližšie k periférii receptívneho poľa.
Recepčné polia sekundárnych senzorických receptorov sú organizované podobným spôsobom. Rozdiel je v tom, že vetvy aferentného vlákna nekončia voľne, ale majú synaptické kontakty s citlivými receptorovými bunkami. Chuťové, vestibulárne, akustické receptívne polia sú organizované týmto spôsobom.
prekrývajúcich sa receptívnych polí. Rovnaká oblasť citlivého povrchu (napríklad koža alebo sietnica) je inervovaná niekoľkými citlivými nervové vlákna, ktoré svojimi vetveniami prekrývajú receptívne polia jednotlivých aferentných nervov.
V dôsledku prekrytia receptívnych polí sa zväčšuje celková zmyslová plocha tela.
Klasifikácia reflexov.
Podľa typu vzdelania:
Podmienené (získané) - reagujte na meno, sliny zo psa do svetla.
Bezpodmienečné (vrodené) - blikajúce prehĺtanie, koleno.
Podľa polohy receptory:
Exteroceptívne (kožné, zrakové, sluchové, čuchové)
Interoceptívne (z receptorov vnútorných orgánov)
Proprioceptívne (z receptorov vo svaloch, šľachách, kĺboch)
Pre efektory:
Somatické alebo motorické (reflexy kostrového svalstva);
Vegetatívne vnútorné orgány – tráviace, kardiovaskulárne, vylučovacie, sekrečné atď.
Podľa biologického pôvodu:
Obranné alebo ochranné (reakcia na rozdelenie hmatovej bolesti)
Tráviace (dráždivé receptory v ústnej dutine.)
Sexuálne (hormóny v krvi)
Približné (otočenie hlavy, tela)
Motor
Posotonické (podporné držanie tela)
Podľa počtu synapsií:
Monosynaptické, ktorých oblúky pozostávajú z aferentných a eferentných neurónov (napríklad koleno).
Polysynaptické, ktorých oblúky obsahujú aj 1 alebo viac intermediárnych neurónov a majú 2 alebo viac synaptických spínačov. (somatické a vegetatívne odkazy).
Disynaptické (2 synapsie, 3 neuróny).
Podľa povahy odpovede:
Motor \ motor (svalové kontrakcie)
Sekrečné (sekrečné žľazy)
Vazomotorické (rozšírenie a zúženie krvných ciev)
Srdcové (zmena. Práca srdcového svalu.)
Podľa trvania:
fázový (rýchly) odber ruky
tonické (pomalé) udržiavanie polohy
Podľa umiestnenia nervového centra:
Miecha (zapojené neuróny SM) – odtiahnutie ruky od horúcich segmentov 2-4, trhnutie kolena.
reflexy v mozgu
Bulbar (medulla oblongata) - zatváranie očných viečok pri dotyku. k rohovke.
Mesencephalic (stredný m) - orientačný bod videnia.
Diencephalic (stredný mozog) - čuch
Kortikálna (kôra BP GM) - podmienená. ref.
Vlastnosti nervových centier.
1. Jednostranné šírenie vzruchu.
Vzruch sa prenáša z aferentného na eferentný neurón (dôvod: štruktúra synapsie).
Spomalenie prenosu vzruchu.
Kondicionér Prítomnosť mnohých synapsií závisí aj od sily dráždidla (suma) a od fyzického stavu. CNS (únava).
3.Zhrnutie súhrn účinkov, podprahové podnety.
Dočasné: ref. Z predch. Imp-sa ešte neprešla, ale stopa. Už prišiel.
Priestorové: miešanie viacerých. Stojatá voda Sú upravené sovou. Snímky. Ref.
Facilitačné a oklúzne centrum.
Objavil sa stred reliéfu - nastáva pri pôsobení optimálneho stimulu (maximálna odozva). Centrum pomoci.
Pri pôsobení min irr. (znížená otv. Rektsiya) došlo k upchatiu.
Asimilácia a transformácia rytmu excitácie.
Transformácia - zmena frekvencie nervového impulzu pri prechode nervovým centrom. Frekvencia sa môže zvýšiť alebo znížiť.
Asimilácia (tanec, denná rutina)
Dôsledok
Oneskorenie konca reakcie po ukončení stimulu. Súvisí s obehovým nervom. Imp. ZATVORENÉ Okruhy neurónov.
krátkodobé (zlomky sekundy)
dlhé (sekundy)
Rytmická činnosť nervových centier.
Zvýšenie alebo zníženie frekvencie nervových impulzov spojených s vlastnosťami synapsie a integračným trvaním neurónov.
8. Plasticita nervových centier.
Schopnosť prebudovať funkčnosť vlastnosti pre efektívnejšiu reguláciu funkcií, implementáciu nových reflexov, ktoré predtým neboli charakteristické pre toto centrum, alebo obnovenie func. V srdci vrstvy synps je zmena v mall-th str-ry.
Zmeny excitability pod vplyvom chemikálií.
Vysoká citlivosť na skutočný rozdiel.
Únava nervových centier.
Súvisí s vysokou únavou synapsií. Znížte pocity. Receptory.
Všeobecné princípy koordinačnej činnosti centrálneho nervového systému.
brzdenie-špeciálny ner. percent prejavujúce sa znížením alebo úplným vymiznutím resp. reakcie.
Princíp konvergencie
Konvergencia je konvergencia impulzov prichádzajúcich cez rôzne aferentné dráhy v ktoromkoľvek centrálnom neuróne alebo nervovom centre.
2 . S princípom úzko súvisí princíp konvergencie spoločná konečná cesta otvorte Sherrinkton. Mnoho rôznych stimulov môže vzbudiť rovnaký motorický neurón a rovnakú motorickú odpoveď. Tento princíp je spôsobený nerovnakým počtom aferentných a eferentných dráh.
Princíp divergencie
Toto je kontakt jedného neurónu s mnohými ďalšími.
Ožarovanie a koncentrácia vzruchu.
Šírenie procesu excitácie do iných nervových centier sa nazýva ožarovanie (volebné- v jednom smere , zovšeobecnené- rozsiahly).
Po určitom čase je ožarovanie nahradené fenoménom koncentrácie excitácie v rovnakom počiatočnom bode CNS.
Proces ožarovania zohráva pozitívnu (tvorba nových podmienených reflexov) a negatívnu (porušenie jemných vzťahov, ktoré sa vyvinuli medzi procesmi excitácie a inhibície, čo vedie k poruche motorickej aktivity).
Princíp reciprocity (spomaľuje)
Excitácia niektorých buniek spôsobuje inhibíciu iných prostredníctvom interkalárneho neurónu.
Dominantný princíp
Ukhtomsky sformuloval princíp dominancie ako pracovný princíp činnosti nervových centier. termín dominantný označuje dominantné ohnisko vzruchu centrálneho nervového systému, ktoré určuje aktuálnu činnosť organizmu.
Princípy dominantného zamerania :
Zvýšená excitabilita nervových centier;
Pretrvávanie excitácie excitácie v priebehu času;
Schopnosť sumarizovať vonkajšie podnety;
Zotrvačnosť (schopnosť udržať vzrušenie po dlhú dobu po ukončení pôsobenia podráždenia); schopnosť spôsobiť konjugované inhibície.

