Tuki- ja liikuntaelimistön tehtävänä on pitää kehoa tietyssä asennossa ja liikkua avaruudessa. Tuki- ja liikuntaelimistöön kuuluu luusto (luuranko), nivelsiteet, nivelet ja luurankolihas. Luut, nivelsiteet ja nivelet ovat liikkuvien elinten passiivisia elementtejä. Lihakset ovat moottorijärjestelmän aktiivinen osa.
Somatomotorisen hermoston evoluutio
Seuraavassa käsittelemme vain somatomotorista hermostoa, joka hermottaa luurankolihaksia. Animaatio: lihasten supistumisen molekyylimekanismi. Somatomotorinen hermosto muuttuu ja kehittyy vähitellen evoluution aikana. Kehitys tapahtuu rinnakkain luurankolihasten muodostumisen ja liikemekanismin kanssa, koska rakenteet ovat riippuvaisia toisistaan. Evoluution aikana voimme puhua eläimistä vain, jos ne toimivat lihas näkyy ihmiskehossa. Alkukantaiset, yksinkertaisesti rakennetut eläimet, joilla on verkkohermosto, reagoivat törmäykseen ympäristöön kokonsa voimakkaalla liikkeellä lihasmassa, jonka tarkoituksena oli välttää haitallisia vaikutuksia tai lähestyä ruokaansa.
Luuranko toimii tukena ja suojana koko keholle ja yksittäisiä elimiä, ja monet luut ovat myös voimakkaita vipuja, joiden avulla suoritetaan erilaisia kehon ja sen osien liikkeitä avaruudessa. Lihakset saavat liikkeelle koko luun vipujärjestelmän. Luuranko muodostaa kehon perustan ja määrittää sen koon ja muodon. luurangon osat, kuten kallo, rintakehä ja lantio selkäranka, toimivat elintärkeiden elinten - aivojen, keuhkojen, sydämen, suoliston jne. - säilytys- ja suojapaikkana. Luuranko osallistuu aineenvaihduntaan, erityisesti tietyllä tasolla pysymiseen mineraalikoostumus verta. Lisäksi useat luut muodostavat aineet (kalsium, fosfori, sitruunahappo jne.) pääsevät tarvittaessa helposti aineenvaihduntaprosesseihin. Luuaineen orgaaninen perusta koostuu pääasiassa proteiineista, kun taas mineraali koostuu kalsium- ja fosforisuoloista.
Heidän yksinkertainen hermostonsa ja lihaksisto erilaista liikettä ei sallittu. Evoluution aikana kehittyi yhä täydellisempiä liikkeitä. Ne tulivat mahdollisiksi kehityksen, erilaistumisen ansiosta hermosto ja liikemekanismit ja niiden harmonisointi.
Ilman kiinteää luurankoa elävät nilviäiset käyvät lihasrenkaidensa aaltoilevan peristalttisen supistuksen läpi. Lihasrenkaiden segmenttejä ohjaavat gangliosolut, jotka ovat pitkittäisesti yhteydessä toisiinsa hermosäikeitä liikkeen synkronoimiseksi.
Luiden kovilla ja sileillä pinnoilla on suurennettuna huokoinen rakenne. Olla olemassa erilaisia tyyppejä luukudosta, jotka ovat useimmiten eri osat yksi luu: tiivis kerros ja sienimäinen aine. Luut, kuten nikamat, reisiluun kaula, epifyysi säde, koostuvat pääasiassa sienimäisestä aineesta. Sienimäisessä aineessa luupalkit on järjestetty kaareviksi levyiksi, jotka on yhdistetty poikittais- tai vinosti juoksevilla poikkipalkeilla. Raajojen pitkät putkiluut koostuvat pääasiassa aineesta, jossa luulevyt sijaitsevat erittäin tiukasti.
Aluksi hajanainen paikallinen lihaskuituja muodostavat myöhemmin vuorovaikutuksessa olevia lihaskimppuja; lihaksia. Kiinteän luurangon evoluutio mahdollisti nopean kehityksen, joka ratkesi eri tavalla niveljalkaisilla ulkoisena luurankona, selkärankaisissa - sisäisenä luurankona. Vaikka molemmat ovat tehokkaita kehon liikkeen auttamisessa, jälkimmäinen ratkaisu osoittautui hyödyllisemmäksi, koska se mahdollisti tasaisen kasvun, mikä mahdollisti suurempien, monimutkaisempien elävien olentojen luomisen.
Niveljalkaisten hermosto on segmentoitu. Raajan aktiivisuutta mediaalisessa segmentissä säätelevät segmentaaliset hermosolmut. Niveljalkaisten gangliot voivat toimia itsenäisesti. Vaikka pään ganglio on suurempi kuin toinen, se ei hallitse muiden ganglioiden toimintaa. Sen koko riittää lähinnä silmiin, antenneihin ja suuhun, jotka säilytetään päähän. Kun niveljalkaiselta leikataan pää, eläin voi myös liikkua useita päiviä, kunnes se näkee nälkää ilman suuta. Interganglioniset yhteydet koordinoivat raajojen liikkeitä.
Luut kuten muutkin sisäelimet koostuvat soluista. On olemassa erityisiä soluja, jotka jatkuvasti tuhoavat luuainetta (osteoklastit); solut, jotka uudistavat, palauttavat luuta (osteoblastit) ja solut, jotka vastaavat luurungon muodostumisesta ja luukudoksen mineralisaatiosta (osteosyytit).
Ihmisen elämän aikana luissa tapahtuu jatkuvasti luukudoksen uudelleenjärjestelyprosesseja: jollain erikseen otetulla pienellä luukudoksen alueella luukudos tuhoutuu, sitten poistetun vanhan luun tilalle tulee täsmälleen sama määrä uutta luuta. . Terveillä ihmisillä luukudoksen tuhoutumisprosessit ja uuden luun muodostuminen ovat kvantitatiivisesti samat. Näiden solujen toimintaa säätelevät monet biologisesti aktiiviset aineet, kuten kilpirauhasen ja lisäkilpirauhasen hormonit, lisämunuaiskuoren hormonit, D3-vitamiini ja lopuksi sukupuolihormonit (estrogeenit ja progesteroni). Luukudoksen kasvu ja kehitys tapahtuu 16-25 vuoden iässä. Luumassan maksimitason saavuttamisen jälkeen, 30-40-vuotiaana, alkaa sen vähäinen menetys, joka on 0,2-0,5 % vuodessa.
verrattuna hänen monimutkainen toiminto, hermosoluja löytyy yllättävän vähän gangliosta. Koko raajan lihaksessa on yleensä neljä neuronia. Hermosolujen väliset yhteydet tarjoavat molemminpuolista hermotusta koukistus- ja ojentajalihasten välillä ja koordinoivat liikettä muiden raajojen kanssa.
Ensimmäiset selkärankaiset ovat vedessä eläviä eläimiä; Kalastaa. Aluksi heidän luurankolihaksensa koostuivat vain aksiaalisista lihaksista, jotka nikamahappo absorboivat ja jotka olivat peräisin myotomista. Lihakset tarjoavat vain kehon taivutuksen ja kääntymisen. Ensimmäinen homogeeninen aksiaalinen lihasmassa segmentoidaan myöhemmin; siten yksittäiset lihakset kehittyivät. Sitten lihaksia venytettiin myös kehon seinämässä. Evoluution seuraavana tuloksena evät, ja sitten, kun jotkut uskalsivat mantereelle, käyttöön raajat.
30-40 vuoden iässä luukato on 0,5 % vuodessa. Ja vaihdevuosien alkamisen jälkeen naisilla menetetään 3-5% luumassasta vuodessa.
Ihmiskehossa on myös eristetty pehmeä luuranko (luuranko), joka osallistuu elinten pitämiseen luiden lähellä. Pehmeä luuranko sisältää fasciaa, nivelsiteitä, elinten sidekudoskapseleita ja muita rakenteita. Suurin osa lihaksista on kiinnittynyt luihin. Lihakset sisältävät luuston luut liikkeessä ja tekevät työn. Monet kehon onteloita ympäröivät lihakset suojaavat sisäelimiä.
Lihakset, jotka tarjosivat nämä rakenteet, puhkesivat ulos kehon seinästä. Raajojen kehitys oli harppaus selkärankaisten liikkumisjärjestelmän kehityksessä. Raajat mahdollistavat elävien olentojen liikkumisen tehokkaammin, joten niiden elinolot, sopeutuminen ympäristöön ovat parantuneet merkittävästi. Evoluution aikana niiden rakennetta ja kokoa on parannettu jatkuvasti. Kehon kokoa huomattavasti suuremmat raajat sallivat kaksijalkaisen elämän. Tämän seurauksena he pystyivät paremmin navigoimaan korkeammat aistielimet päässään, ja heidän liikkumiskuormasta vapautettuja käsiään voitiin käyttää muihin tarkoituksiin.
Luiden kunto riippuu niiden kantamasta kuormasta. Hyvin kehittynyt lihaskudos vahvistaa niveliä ja edistää luiden normaalia kehitystä ja toimintaa. Sekä lihakset että luut menettävät massaansa, jos niille kohdistuva kuormitus on liian pieni. Siksi tuki- ja liikuntaelimistön pitämiseksi terveessä tilassa pitkään on välttämätöntä harjoitella jatkuvasti, suorittaa erilaisia fyysinen harjoitus. Jokainen yli 30 vuoden ikäinen ihminen on tuomittu liikuntakasvatukseen.
Nämä ja vastaavat käsien pienten nivelten uudelleenjärjestelyt rohkaisivat työkalujen käyttöön, mikä nopeuttai myös aivojen, jotka olivat tärkein ihmisperäisen sopeutumiskyvyn elin, kehitystä. Kahdenvälisyys korosti erityisesti motorisen järjestelmän kehitystä ja samalla vaikutti osaltaan, sillä kehon pystyasento vaatii paljon enemmän motorista koordinaatiota kuin neljän jalan korkeudella seisominen.
Raajojen laajentuminen muutti myös lihaksia. Paljon suuremmassa käytettävissä olevassa tilassa sekä kertominen että raajan pidentäminen ovat sallittuja. Ne auttavat lisäämään sekä lihasvoimaa että pienempien liikkeiden suorituskykyä. Liikkumismekanismin kehittyminen on antanut uusia mahdollisuuksia selkärankaisille. Niiden liikkeestä tulee täydellisempää, nopeampaa, tarkempaa ja sopivampaa. Välttämättä kehitetyt hermorakenteet, jotka ohjaavat luurankolihasten toimintaa. Liikkeen liikkumista helpotti samanaikaisesti kehittyneet aistielimet.
Tämä on erityisen tärkeää naisille, joiden luut ovat vähemmän tiheitä kuin miesten. Lisäksi naiset ovat alttiimpia menettämään lihasmassaa vanhuudessa.
Luun massaa, sen muodostumisastetta ja sen häviämistä voidaan hallita hyvää ravintoa. Kalsium ja D-vitamiini vahvistavat luita ja estävät niitä halkeilemasta, murtumasta ja muista vammoista. Tarvittava kalsiumin vuorokausiannos aikuisille on noin 1200-1500 mg. Naiset, kuten miehet, kasvattavat 50 % luumassastaan 20 ikävuoteen mennessä. Säännölliseen liikuntaan yhdistettynä kalsium edistää tervettä, vahvaa luumassaa. 20–30 vuoden iässä luumassa kasvaa hieman, ja 30 vuoden kuluttua alkaa sen häviämisprosessi.
Heidän avullaan ihminen voi saada ympäristöstään entistä kattavampaa ja tarkempaa tietoa. Näitä mahdollisuuksia hyödyntääkseen liikekoordinaatiota tukevat neuronit lisääntyivät aivoissa ja muodostivat yhä monimutkaisempia ja intensiivisempiä yhteyksiä aistielimiin pitääkseen ne aktiivisena. Aksiaalisia lihaksia ohjasivat alun perin yksinomaan selkärangan motoriset neuronit. Yksittäisten lihasten erottamisen jälkeen yksittäisten lihasten hallinnan ottivat selkärankaisilla kehittyvät aivot, joilla on keskeinen ohjausrooli.
Kehosi biologisen toiminnan yhteydessä erityistä huomiota terveellinen ruokavalio pitäisi antaa naisille. Raskaus ja imetys tarvitsevat lisää kalsiumia. Jos naisen veri ei sisällä vaadittava määrä Tästä mineraalista luista tulee lisäkalsiumin lähde, mikä lopulta johtaa niiden heikkenemiseen. tupakointi ja liikakäyttö Alkoholijuomat edistää myös luun menetystä. Estrogeenihormoni säätelee kalsiumin huuhtoutumista luista ja luo näin normaalit olosuhteet luun pitkäaikaiselle kasvulle. Vaihdevuosien ja vaihdevuosien jälkeisenä aikana naisen keho kuitenkin lakkaa tuottamasta estrogeenia, joka on pääasiallinen syy kiihtyneeseen luukatoon. Säännöllinen liikunta ja suurempi päivittäinen kalsiumin saanti auttavat estämään tätä prosessia kuukautisten jälkeisenä aikana. Sen lisäksi, että kalsiumia otetaan ravintolisänä, on suositeltavaa sisällyttää tätä alkuainetta sisältävä ruoka päivittäiseen ruokavalioon.
Ensimmäinen selkärankaisen autokeskus oli lokomotorinen alue, joka sijaitsee mesencephalonissa. Vaikka evoluutio on kehittynyt uusille aivojen alueille, jotka ovat erikoistuneet motorisen ohjauksen hallintaan, retikulospastinen kanava säilyy myös ihmisillä. Lihasjännityksen hallinnan lisäksi hän osallistuu pääasiassa rytmisen liikkeiden suorittamiseen.
Evoluution aikana selkärankaisten aivot hallitsevat alun perin osaa nykyaikaisesta aivorungosta. Kun aivojen toiminnoista tuli monimutkaisempia, tarvittiin lisää hermosoluja niiden hallitsemiseen. Koska aivoissa ei tuolloin ollut tarpeeksi tilaa, uudet neuronit muodostivat uuden pesäkkeen tähän asti sijainneiden aivojen eteen. Seuraavassa vaiheessa muodostuu kolme näköstä vastaavaa ydintä: kaksi väliaivojen taakse ja yksi yläpuolelle. Aikaisemmilla selkärankaisilla on kolme silmää. Kahden lateraalisen silmän tiedot käsiteltiin kahdella basalokatiivisella lateraalisella genikulaattikorpuskkelilla, jotka yllä olevat parietaalisilmät risteytyvät ylemmän ytimen kautta.
Ohjelmaan fyysiset aktiviteetit painoharjoittelun pitäisi olla mukana. Tärkeä tekijä, joka määrittää tuki- ja liikuntaelimistön terveyden, on optimaalisen painon ylläpitäminen.
Erityistä huomiota tulee kiinnittää jalkoihin. Jaloilla on erittäin tärkeä rooli tuki- ja liikuntaelinten sekä koko kehon terveydelle. Asianmukainen hoito jalkojen taakse kuuluu muiden hygieniatoimenpiteiden lisäksi huolellinen kenkien valinta. Mukavat ja oikein valitut kengät ovat erinomainen ehkäisy monille jalkasairauksille.
Sivusilmät ja niiden tumat säilyivät ihmisellä myös näköeliminä, kun taas parietaalinen silmä ja sen tuma vähitellen väistyivät, menettivät näkönsä ja vajosivat syvyyksiin kuin epitalamus. Seuraava toiminto päiväkefalonissa oli somatosensorinen. Matelijoilla somatosensorisen toiminnan hoito kehittyi edelleen; dorsaaliset ja anterioriset talamuksen tumat kehittyivät. Samanaikaisesti anteroventraalinen tuma oli ensimmäinen koordinaatiokeskus mesencephalonille.
Talamuksen viimeinen kehittynyt alue oli Pulvinar Talami, joka hoiti aistielinten välistä koordinaatiota. Ei-nisäkkäillä se otti koko aivojen kolmen sensorisen lohkon roolin. Sammakkoeläimistä on jo löydetty tyviganglioita, jotka koordinoivat liikejärjestelmää. Ne kehittyivät nopeasti, mutta vain nisäkkäissä.
"Tuki- ja liikuntaelimistö" ja muut artikkelit osiosta Tuki- ja liikuntaelinten sairaudet
Vapaaehtoisen liikkeen toiminnallinen rakenne. Edellä olevasta seuraa, että kaikenlaisen liikkeen varmistamisessa on mukana eri komponentteja, joten yksi pääkysymyksistä on, miten varmistetaan toimeenpanolaitteen vastaanottama kertakäyttöinen komento. Tietyn liikkeen strategiasta ja taktiikoista riippumatta ohjelman tarjoavan järjestelmän päätehtävä on koordinoida kaikkia ryhmän komponentteja.
Dieenisolun sivuseinistä ulottuvia uraanin telenkefalon-kertymiä on jo muodostumassa sammakkoeläimiin. Ensimmäiset telencefaaliset rakenteet vastaavat tyviganglioiden ja nisäkkään limbisen järjestelmän kasveja. Heidän päätehtävänään olivat hajuaisti ja tiettyjen, erityisesti seksuaalisten käytösten koordinointi. Aivokuori on nisäkkäiden "keksintö". Edellä esitettyjen perusteiden mukaan löytyy sellaisia uusia toimintoja, joiden hermosolut eivät ole löytäneet riittävästi tilaa välipäässä.
Kun päiväkefaloni ei enää voinut kasvaa, uusia hermosoluja syntyi sen seinämästä kasvaneen neuroepiteeliputken seinämästä. Sivukammiot kehittyivät vähitellen ja enkefalonin lohkot niiden seinistä. Ensimmäiset nisäkkäät olivat pieniä, sauvan muotoisia jyrsijöitä, jotka aluksi heittelivät ympäriinsä ja muuttuivat sitten pussimaisiksi. Dinosaurusten aikana he turvautuivat metron luoliin. Elämäntapansa mukaisesti he navigoivat pimeissä luolissa ensisijaisesti hajunsa perusteella.
CNS sillä on useita geneettisesti kiinteitä ohjelmia (esimerkiksi toimintaan perustuva liikkuva kävelyohjelma selkäranga - liittyy selkärankaan.");" onmouseout="nd();" href="javascript:void(0);">selkäydin generaattori). Tällaiset yksinkertaiset ohjelmat yhdistetään monimutkaisempiin järjestelmiin, kuten pystyasennon ylläpitämiseen. Tämä assosiaatio tapahtuu oppimisen seurauksena, joka saadaan aikaan anteriorisen aivokuoren osallistumisen kautta. pallonpuoliskot.
Monimutkaisin ja fylogeneettisesti nuorin on kyky muodostaa liikesarja ja ennakoida sen toteutumista. Ratkaisu tähän ongelmaan liittyy frontaaliseen assosiatiiviseen järjestelmään, joka muistaa ja tallentaa muistiin tällaiset liikesarjat. Tämän koodauksen korkein heijastus ihmisessä on liikkeen peruskäsitteiden verbalisointi tai sanallinen säestys.
Liikeohjausjärjestelmän yleinen säännöllisyys on takaisinkytkennän käyttö. Tämä ei sisällä vain proprioseptiivistä palautetta alkaneesta liikkeestä, vaan myös Aktivointi - viritys tai lisääntynyt aktiivisuus, siirtyminen lepotilasta aktiiviseen tilaan. onmouseout="nd();" href="javascript:void(0);">aktivointi palkitsemis- tai rangaistusjärjestelmät. Lisäksi mukana on myös sisäinen palaute, ts. tiedot moottorijärjestelmän taustalla olevien tasojen toiminnasta tai efferentti kopio itse moottorikäskystä. Tämäntyyppinen palaute on tarpeen uusien motoristen koordinaatioiden kehittämiseksi. Vaihtelevan monimutkaisuuden ja nopeuden liikkeissä takaisinkytkentä voidaan sulkea eri tasoilla. Siksi molemmat ohjaustyypit - ohjelmointi ja seuranta - voivat esiintyä rinnakkain samassa liikkeenohjausjärjestelmässä.
Lopuksi on suositeltavaa lainata erinomaisen fysiologin N.A. Bernsteinin mukaan liikkeet "...eivät ole spatiaalisia, vaan semanttisia ja toimintatason ketjujen motoriset komponentit sanelevat ja valitaan kohteen semanttisen olemuksen ja sen mukaan, mitä sillä pitäisi tehdä."
Selviytymisen kannalta kaksi tärkeintä toimintoa ovat haju ja lisääntyminen. Siten näitä kahta toimintoa on käsitelty kahdella uudella kehitysalueella, joka on peräisin dienecephalonista. heidän jäännöksensä, limbinen järjestelmä ja hajukuori, joka suorittaa samoja tehtäviä, löytyy myös ihmisistä mediaalisesta keskuskuoresta. Nämä kaksi toimintoa liittyvät läheisesti morfologisesti ja toiminnallisesti jokaiseen selkärankaiseen, jopa ihmisiin, joiden haju on huomattavasti vähentynyt. Hajuvesiteollisuus perustuu tähän asenteeseen.
Corpus callosumin puolipallot puuttuvat vanhimpien elossa olevien nisäkkäiden, kloaakkojen ja lintujen aivoista. Seuraava askel evoluutiossa oli somatosensorisen järjestelmän merkittävä kehitys. Nämä yölliset jyrsijät, jotka ovat samanlaisia kuin nykypäivän rotat ja hiiret, asuivat edelleen pimeässä maan alla. He pystyivät nopeasti ja luotettavasti navigoimaan pimeillä maanalaisilla kanavillaan pitkät hiukset kasvoille, koska he voisivat käyttää niitä tutkan tavoin, yhdistettyjen erittäin herkkien painereseptorien kautta. Tätä tarkkaa mekaanista "kuva-analyysiä" varten he tarvitsivat uuden neuronipopulaation.
10.4 Liikkeen organisoinnin elektrofysiologiset korrelaatit
Eri näkökohtien tutkimiseen käytetään sähköfysiologisia menetelmiä motorista toimintaa ja erityisesti ne, jotka eivät ole suoran havainnoinnin ulottuvilla. Arvokasta tietoa liikeorganisaation fysiologisista mekanismeista antavat menetelmät aivokuoren vyöhykkeiden vuorovaikutuksen arviointiin, paikallisen EEG:n ja liikkeeseen liittyvien potentiaalien analysointiin sekä hermosolujen toiminnan tallentamiseen.
Aivojen biopotentiaalien vyöhykkeiden välisten yhteyksien tutkiminen mahdollistaa yksittäisten aivokuoren vyöhykkeiden vuorovaikutuksen dynamiikan jäljittämisen liikkeen suorittamisen eri vaiheissa uusia motorisia taitoja opetettaessa, tunnistaa vyöhykkeiden välisen vuorovaikutuksen erityispiirteet aikana. erilaisia tyyppejä liikkeet.
Nämä hermosolut siirtyivät vasta muodostuneeseen teleaivokehään ja muodostivat ensimmäisen todellisen lohkon - parietaalinen lohko. Tämä läppä pysyi myös ihmisen somatosensorisena keskuksena. Opossumissa ja vastaavissa lajeissa aivokuorta käytetään pääasiassa ihon parantamiseen. Vanhoilla nisäkkäillä signaalit kulkivat lihaskarista vasta kehittyneeseen pikkuaivoon ja sieltä pikkuaivojen ytimien kautta talamukseen.
Seuraavana evoluution vaiheena jo maan pinnalle matkaavien kanimaisten jyrsijöiden aivot kohtasivat uuden haasteen: heidän oli kuunneltava kaukaa ja löydettävä mahdollinen hyökkäävä vihollinen, jotta he pääsisivät pakoon. aika. Uusi akustinen keskus on siirtynyt alemmasta colliculuksesta kohti ajallinen lohko, joka kehittyy neuroepiteeliputken seinämästä, joka kasvaa ulos dienosiphalonista ja heilahtaa alas tilanpuutteen vuoksi. Osat sivukammio, joka sisältää kaksi plexuksen verisuoniseinämää, soluväliaineen ja alemman maissin sekä parietaali- ja ajallinen lohko, jotka kehittyvät sen seinämästä, ovat aivokuoren vanhimmat osat.
Tilasynkronointi (PS), eli Aivokuoren eri kohdista tallennettujen sähköisten värähtelyjen synkroninen dynamiikka heijastaa aivojen rakenteiden tilaa, mikä helpottaa virityksen leviämistä ja luo olosuhteet vyöhykkeiden väliselle vuorovaikutukselle. PS-rekisteröintimenetelmän on kehittänyt erinomainen venäläinen fysiologi M.N. Livanov.
Tutkimukset yksittäisten vyöhykkeiden EEG:n rytmisistä komponenteista ja niiden tila-aikasuhteista ihmisillä vapaaehtoisten liikkeiden suorittamisen aikana todellinen mahdollisuus lähestyä motorisen toiminnan aikana systeemitasolla kehittyvien toiminnallisten vuorovaikutusten keskeisten mekanismien analysointia. Rytmisten liikkeiden suorittamisen aikana tallennetun EEG:n korrelaatioanalyysi osoitti, että ihmisillä ei vain motorisen aivokuoren keskukset, vaan myös etu- ja alaparietaalivyöhykkeet osallistuvat aivokuoren liikkeiden järjestämiseen.
Vapaaehtoisten liikkeiden oppiminen ja niiden harjoittelu aiheuttavat kortikaalisten biopotentiaalien keskusten välisten korrelaatioiden uudelleenjakautumista. Harjoittelun alussa kokonaismäärä yhteistoimintaan osallistuvat keskukset lisääntyvät jyrkästi, ja motoristen vyöhykkeiden EEG:n rytmisen komponenttien synkroninen suhde etu- ja takaosan kanssa Aivokuoren assosiatiiviset vyöhykkeet - vyöhykkeet, jotka vastaanottavat tietoa reseptoreista, jotka havaitsevat erilaisten modaliteettien ärsytystä, ja kaikilta projektioalueilta. ");" onmouseout="nd();" href="javascript:void(0);">assosiatiivinen alueilla. Kun liike hallitaan, PS:n kokonaistaso laskee merkittävästi, ja päinvastoin, yhteydet motoristen vyöhykkeiden ja alempien parietaalisten vyöhykkeiden välillä lisääntyvät. 
On tärkeää huomata, että oppimisprosessissa eri aivokuoren vyöhykkeiden biopotentiaalien rytminen koostumus järjestetään uudelleen: EEG:hen aletaan tallentaa hitaita rytmejä, jotka osuvat taajuudellaan liikkeiden suorittamisen rytmiin. Näitä ihmisen EEG:n rytmejä kutsutaan "merkittyiksi". Samat merkittyjä värähtelyjä havaittiin lapsilla esikouluikäinen kun he tekevät rytmisiä liikkeitä ergografissa.
Ihmisen EEG:n systemaattiset tutkimukset syklisen (ajoittain toistuvan) ja asyklisen motorisen toiminnan aikana ovat paljastaneet merkittäviä muutoksia aivokuoren sähköisen aktiivisuuden dynamiikassa. EEG:ssä lisääntyy sekä paikallinen että etäinen biopotentiaalien synkronointi, mikä ilmaistaan jaksollisten komponenttien tehon kasvuna, auto- ja ristikorrelogrammien taajuusspektrin muutoksina, tietynlaisena linjauksena. taajuusspektrien ja funktioiden maksimit Koherenssi - EEG-taajuusindikaattoreiden synkronointiaste aivokuoren eri osien välillä.");" onmouseout="nd();" href="javascript:void(0);">yhdenmukaisuus samalla taajuudella.
PS ja reaktioaika. Reaktioaika on yksi yksinkertaisimmista moottorin ilmaisimista. Siksi erityisen kiinnostavaa on se tosiasia, että jopa yksinkertaisella motorisella reaktiolla voi olla erilaisia fysiologisia korrelaatioita riippuen sen keston kasvusta tai vähenemisestä. Näin ollen, kun vertaillaan kuvaa aivojen EEG:n spektrikomponenttien keskusten välisistä korrelaatiosuhteista yksinkertaisen motorisen reaktion aikaan, kävi ilmi, että assosiatiivisten vyöhykkeiden EEG:n spatiotemporaalisten suhteiden uudelleenjärjestely liittyy vasteeseen. aikaa tiettyyn ärsykkeeseen. Nopeilla reaktioilla terveellä ihmisellä syntyi useimmiten korkeita biopotentiaalien korrelaatioita molemmilla alemmilla parietaalialueilla (hieman enemmän vasemmalla aivopuoliskolla). Jos reaktioaika piteni, siihen liittyi biopotentiaalien synkronointi etuosat aivokuori ja vasemman pallonpuoliskon alempi parietaalinen alue jätettiin vuorovaikutuksen ulkopuolelle. Lisäksi havaittiin suhde vaihesiirtojen suuruusluokkien välillä Alfarytmi - elektroenkefalogrammin päärytmi suhteellisen lepotilassa, taajuudella välillä 8 - 14 Hz ja keskimääräisellä amplitudilla 30 - 70 μV. ");" onmouseout="nd();" href="javascript:void(0);">rytmi alfa rekisteröity etu-, precentraali- ja takaraivoalueet aivot ja yksinkertaisen motorisen reaktion nopeus.
On tärkeää huomata, että biopotentiaalien synkronoinnin lisääntyminen tapahtuu ihmisellä, joka on jo työskentelyä edeltävässä tilassa, keskittymisprosessissa ennen motorista toimintaa sekä henkisen liikkeiden suorittamisen aikana.
PS ja liikkeen erityispiirteet. Biopotentiaalien PS:n epäspesifisen kasvun lisäksi havaittiin sen selvä selektiivinen nousu aivokuoren vyöhykkeiden välillä, jotka osallistuivat suoraan tietyn motorisen toiminnan järjestämiseen. Esimerkiksi suurin samankaltaisuus sähköisessä aktiivisuudessa havaitaan: käsiä liikutettaessa - etuosan ja lihasten motorisen esityksen välillä Yläraajat; liikuttaessa jalkoja - etuosan ja lihasten motorisen esityksen välillä alaraajoissa. Tarkoilla toimilla, jotka vaativat hienoa avaruudellista suuntausta ja visuaalista ohjausta (ammunta, miekkailu, koripallo), visuaalisen ja motorisen alueen vuorovaikutus tehostuu.
Urheilijoiden aivojen eri osien biopotentiaalien PS:n monimutkainen dynamiikka paljastettiin suorituksessa erilaisia harjoituksia ja EEG:n rytmisten komponenttien vuorovaikutuksen lisääntymisen riippuvuus motorisen toiminnan tavasta, urheilijoiden pätevyydestä, henkilön kyvystä ratkaista taktisia ongelmia, tilanteen monimutkaisuudesta on osoitettu. Siten korkeasti pätevillä urheilijoilla keskusten välinen vuorovaikutus on paljon intensiivisempää ja selvemmin paikallistunutta. Kävi myös ilmi, että monimutkaisemmat motoriset tehtävät vaativat enemmän korkeatasoinen EEG-rytmien spatiaalinen synkronointi ja taktisten tehtävien ratkaisemiseen kuluva aika korreloi keskusten välisten vuorovaikutusten lisääntymisnopeuden kanssa. Tässä tapauksessa motorinen vaste seuraa sen jälkeen, kun aivokuoren biopotentiaalien enimmäissynkronointi on saavutettu.
Yhdessä ihmisten aivojen biopotentiaalien PS-tutkimukset ovat mahdollistaneet sen, että suoritettaessa yksinkertaisia ja monimutkaisia motorisia toimintoja aivojen eri keskukset joutuvat vuorovaikutukseen muodostaen monimutkaisia toisiinsa yhdistetyistä vyöhykkeistä koostuvia järjestelmiä, joiden aktiivisuuspisteet eivät vain projektiossa, mutta myös assosiatiivisilla alueilla, erityisesti etu- ja alaparietaalissa. Nämä keskusten väliset vuorovaikutukset ovat dynaamisia ja muuttuvat ajassa ja tilassa, kun moottoritoimi suoritetaan.
10.5. Liikkeisiin liittyvien aivojen potentiaalien kompleksi
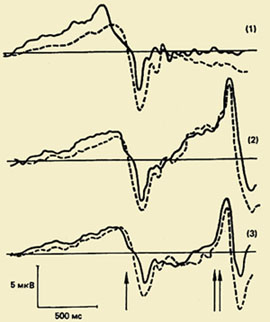 Yksi tärkeimmistä suunnasta motorisen toiminnan psykofysiologian tutkimuksessa on liikkeisiin liittyvien aivopotentiaalien värähtelykompleksin (PMSD) tutkimus. Tämän ilmiön merkitys ymmärtämiselle fysiologiset mekanismit liikkeen organisointi on erittäin laaja, koska PMSD:n tutkimus mahdollistaa aivokuoressa tapahtuvien prosessien piilotettujen sekvenssien paljastamisen liikkeen valmistelun ja suorittamisen aikana, ja tiedonkäsittelyprosessien kronometria on joukko menetelmiä aivokuoressa tapahtuvien prosessien mittaamiseen. mittaukseen perustuvan tietojenkäsittelyn yksittäisten vaiheiden kesto fysiologiset indikaattorit, erityisesti herätettyjen ja tapahtumaan liittyvien potentiaalien komponenttien piilevät jaksot.");" onmouseout="nd();" href="javascript:void(0);"> kello nämä prosessit, ts. asettaa aikarajansa.
Yksi tärkeimmistä suunnasta motorisen toiminnan psykofysiologian tutkimuksessa on liikkeisiin liittyvien aivopotentiaalien värähtelykompleksin (PMSD) tutkimus. Tämän ilmiön merkitys ymmärtämiselle fysiologiset mekanismit liikkeen organisointi on erittäin laaja, koska PMSD:n tutkimus mahdollistaa aivokuoressa tapahtuvien prosessien piilotettujen sekvenssien paljastamisen liikkeen valmistelun ja suorittamisen aikana, ja tiedonkäsittelyprosessien kronometria on joukko menetelmiä aivokuoressa tapahtuvien prosessien mittaamiseen. mittaukseen perustuvan tietojenkäsittelyn yksittäisten vaiheiden kesto fysiologiset indikaattorit, erityisesti herätettyjen ja tapahtumaan liittyvien potentiaalien komponenttien piilevät jaksot.");" onmouseout="nd();" href="javascript:void(0);"> kello nämä prosessit, ts. asettaa aikarajansa.
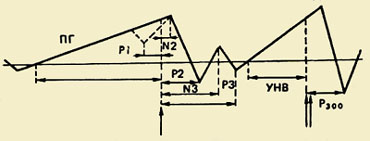
PMSD:n komponenttien koostumus. Tämä liikkeen valmistelu-, toteutus- ja arviointiprosesseja heijastava kompleksi rekisteröitiin ensimmäistä kertaa 60-luvulla. Kävi ilmi, että liikettä edeltää hidas negatiivinen värähtely - valmiuspotentiaali (RP). Se alkaa kehittyä 1,5 - 0,5 s ennen liikkeen alkamista. Tämä komponentti on rekisteröity pääasiassa molempien pallonpuoliskojen keski- ja fronto-keskijohtoihin. 500-300 ms ennen liikkeen alkua PG muuttuu epäsymmetriseksi - sen suurin amplitudi havaitaan precentral-alueella, Kontralateraalinen - vartalon vastakkaiselle puolelle.");" onmouseout="nd();" href="javascript:void(0);"> vastakkainen liikettä. Noin puolet aikuisista, tämän hitaan negatiivisen värähtelyn taustalla, vähän ennen liikkeen alkamista, tallennetaan pieni amplitudipositiivinen komponentti. Sitä kutsutaan "premotoriseksi positiiviseksi" (PMP). Seuraava nopeasti kasvava negatiivinen amplitudivärähtely, niin kutsuttu moottoripotentiaali (MP), alkaa kehittyä 150 ms ennen liikkeen alkamista ja saavuttaa maksimiamplitudinsa liikkuvan raajan motorisen esityksen alueella aivokuoressa. Tämä potentiaalikompleksi päättyy positiiviseen komponenttiin noin 200 ms liikkeen alkamisen jälkeen.
Komponenttien toiminnallinen merkitys. On yleisesti hyväksyttyä, että valmiuspotentiaali (RP) syntyy motorisessa aivokuoressa ja liittyy liikkeen suunnittelu- ja valmisteluprosesseihin. Se kuuluu aivopotentiaalin hitaiden negatiivisten värähtelyjen luokkaan, jonka esiintyminen selittyy aivokuoren vastaavien alueiden hermosolujen aktivaatiolla.
Hypoteesit koskien toiminnallinen arvo PMP:t ovat erilaisia.
Tätä värähtelyä pidetään sekä heijastuksena keskuskomennon toimituksesta aivokuoresta lihaksiin että aivokuoren rentoutumisen seurauksena tietyn liikkeen organisointivaiheen jälkeen ja heijastuksena prosesseista. toisen raajan siihen liittyvien liikkeiden tukahduttamiseen ja palautteena lihasafferenteilta. Tällä hetkellä jotkut kirjoittajat uskovat, että PMP:t ovat vain heijastus motorisen potentiaalin alkamisesta.
Kun MP rekisteröitiin apinoissa, MP:ssä tunnistettiin kaksi alakomponenttia. Ensimmäinen osakomponentti korreloi motorisen aivokuoren aktivoitumisen kanssa, joka liittyy liikkeen alkamiseen (synaptinen aktiivisuus pyramidaaliset neuronit), ja toinen - Brodmann-kenttien kenttien 2.3 ja 4 aktivoinnin myötä - erilliset aivokuoren alueet, jotka eroavat solurakenteestaan (sytoarkkitehtoniikka) ja toiminnoistaan. Esimerkiksi kentät 17, 18, 19 ovat aivokuoren visuaalisia alueita, joilla on erilaiset rakenteet ja toiminnot visuaalisen havainnon tarjoamisessa.");" onmouseout="nd();" href="javascript:void(0);">kirjoittaja Brodman . MP:n rekisteröinti epilepsiaa sairastavalla henkilöllä mahdollisti kolmen komponentin erottamisen siinä. Ensimmäistä komponenttia kutsuttiin initiaatiopotentiaaliksi. Sillä on korkea amplitudi ja se tapahtuu liikkeen alkamisen jälkeen precentraalisessa kontralateraalisessa aivokuoressa. Toinen, joka tapahtuu myogrammin alun jälkeen ja on enemmän lokalisoitunut kontralateraaliseen somatosensoriseen kenttään, voidaan yhdistää sekä liikkeen aloitukseen että sensoriseen palautteeseen. Kolmas komponentti heijastaa impulssia, joka tulee lihasafferenteista aivokuoreen.
MP:tä seuraavaa positiivista potentiaalia pidetään heijastuksena perifeerisistä reseptoreista tulevasta käänteisestä afferentaatiosta, motorisista keskuksista nousevasta aktiivisuudesta, motorisen ohjelman ja sen suorituksen neuronaalisen kuvan vertailusta tai aivokuoren prosesseista. rentoutuminen liikkeen jälkeen.
Odotuksen aalto. PMSD:n lisäksi on kuvattu toinenkin sähköfysiologinen ilmiö, joka on oleellisesti lähellä valmiuspotentiaalia. Se on noin potentiaalin negatiivisesta vaihtelusta, joka on tallennettu aivokuoren etuosiin varoituksen ja laukaisusignaalin (reaktion vaativan) välisenä aikana. Tällä vaihtelulla on useita nimiä: odotusaalto, E-aalto, ehdollinen negatiivinen poikkeama (CNV). E-aalto esiintyy 500 ms varoitussignaalin jälkeen, sen kesto pitenee ensimmäisen ja toisen ärsykkeen välisen aikavälin pidentyessä. E-aallon amplitudi kasvaa suoraan verrannollisesti motorisen vasteen nopeuteen käynnistysärsykkeelle. Se lisääntyy huomion jännityksen ja tahdonvoiman lisääntymisen myötä, mikä osoittaa tämän sähköfysiologisen ilmiön yhteyden motorisen toiminnan ja käyttäytymisen vapaaehtoisen säätelyn mekanismeihin yleensä.
10.6. hermotoimintaa
Toiminnalliset aivokuoren pylväät. Ihmisen aivokuoren motorisella vyöhykkeellä on niin sanottuja jättimäisiä pyramideja Beza-solut - aivokuoren pyramidisolut. onmouseout="nd();" href="javascript:void(0);">Betz-solut, jotka on järjestetty erillisiin sarakkeisiin. Samanlaisia tehtäviä suorittavat pyramidisolut sijaitsevat vierekkäin, muuten olisi vaikea selittää tarkkaa aivokuoren somatotooppista organisaatiota. Tällaiset moottoripylväät pystyvät herättämään tai inhiboimaan funktionaalisesti homogeenisten ryhmien Motorinen neuroni (motorinen neuroni) - hermosolu, jonka aksoni hermottaa lihaskuituja. onmouseout="nd();" href="javascript:void(0);">motoriset neuronit.
Yksittäisten pyramidisolujen aktiivisuuden rekisteröinti käyttämällä istutettuja mikroelektrodeja eläimissä, jotka suorittavat erilaisia liikkeitä, mahdollisti perustavan tärkeän tosiasian toteamisen. Aivokuoren neuronit, jotka säätelevät minkään lihaksen toimintaa, eivät ole keskittyneet vain yhteen sarakkeeseen. Moottoripylväs on suurelta osin neuronien toiminnallinen yhdistelmä, joka säätelee useiden tiettyyn niveleen vaikuttavien lihasten toimintaa. Siten motorisen aivokuoren pyramidaalisten neuronien sarakkeissa liikkeet eivät ole niinkään edustettuina kuin lihakset.
Motoristen ohjelmien hermokoodit. Tietojen koodaus neuronissa tapahtuu sen purkautumistiheyden mukaan. Hermosolujen impulssiaktiivisuuden analyysi erilaisten motoristen ohjelmien kehittämisen aikana eläimillä osoitti, että neuronit ovat mukana niiden rakentamisessa. eri osastoja moottorijärjestelmää suorittaessaan tiettyjä toimintoja. Joidenkin ideoiden mukaan motoristen ohjelmien sisällyttäminen tapahtuu ns. komentohermosolujen aktivoitumisen vuoksi. Komentohermosolut ovat puolestaan korkeampien aivokuoren keskusten hallinnassa. Esto on hermostuneisuusprosessi, joka on vastakkainen herättämiseen; ilmenee tietylle kehon järjestelmälle ominaisen toiminnan heikkenemisenä tai lopettamisena. onmouseout="nd();" href="javascript:void(0);">Hidasta komentohermosolu johtaa sen ohjaaman ohjelman pysähtymiseen, herättämiseen, päinvastoin hermopiirin aktivoitumiseen ja motorisen ohjelman toteutumiseen.
Komentohermosolujen osallistuminen aivojen kiinteään toimintaan määräytyy nykyisen motivaation ja tämän motivaation tyydyttämiseen tähtäävän erityisen motorisen ohjelman mukaan. Moottoriohjelma, ollakseen luonteeltaan mukautuva, tulee ottaa huomioon kaikki signaalin kannalta merkittävät ulkoisen ympäristön komponentit, joihin nähden tarkoituksenmukaista liikettä tehdään, ts. perustuu monisensorisen periaatteeseen Konvergenssi on hermosolujen ryhmän aksonien yhdistyminen, joka johtuu synapsien muodostumisesta samaan postsynaptiseen neuroniin. onmouseout="nd();" href="javascript:void(0);">konvergenssi.
Termien sanasto
- liikkuminen
- motorinen neuroni
- artikulaatio
- aktivointi
- johdonmukaisuutta
- Brodmannin kentät
- tietojenkäsittelyprosessien kronometria
Kysymyksiä itsetutkiskelua varten
- Mitä eroa on staattisen ja dynaamisen kehon kuvan välillä?
- Mitkä aivojen rakenteet ovat ratkaisevassa roolissa vapaaehtoisen liikkeen järjestämisessä?
- Miten pyramidaalisen ja ekstrapyramidaalisen järjestelmän toiminnot eroavat toisistaan?
- Mitä liikkeeseen liittyvät aivojen mahdollisuudet antavat ymmärtää liikkeen organisoinnin aivojen mekanismeja?
Bibliografia
- Batuev A.S. Moottorianalysaattorin toiminnot. L.: Nauka, 1970.
- Batuev A.S. Korkeampi hermostunut toiminta. Moskova: Korkeakoulu, 1991.
- Bernstein N.A. Esseitä liikkeiden fysiologiasta ja toiminnan fysiologiasta. Moskova: Nauka, 1966.
- Bloom F., Lizerson A., Hofstadter L. Aivot, mieli ja käyttäytyminen. M.: Mir, 1988.
- Dudel J., Ruegg I., Schmidt R., Janig V. Human Physiology. T. 1 / Ed. R. Schmidt ja G. Thevs. M.: Mir, 1985.
- Ihmisen ja eläimen fysiologian yleinen kurssi / Toim. A.A. Nozdrachev. Moskova: Korkeakoulu, 1991.
- Sologub E.B. Ihmisaivojen sähköinen toiminta motorisen toiminnan prosessissa. L.: Nauka, 1973.
- Khrizman T.P. Lapsen liike ja aivojen sähköinen toiminta. Moskova: Pedagogiikka, 1973.
- Evarts E. Aivomekanismit, jotka ohjaavat liikettä // Brain. M.: Mir, 1982.
Kurssityön ja esseiden aiheet
- N.A:n opetukset Bernstein liikkeen rakenteesta.
- Liikkeiden psykofysiologiset typologiat.
- Ihmisen käsien liikkeet ja niiden neurofysiologiset mekanismit.
- Vapaaehtoiset liikkeet ja niiden neurofysiologiset mekanismit.
- Liikkeisiin liittyvien aivopotentiaalien tutkimuksen historialliset vaiheet.
- Hermotoiminnan rooli liikkeiden rakentamisessa.

