5-10-2012, 19:14
Funzioni della corteccia
Lobo frontale. La funzione principale dei lobi frontali è il controllo dei movimenti volontari, il coordinamento dei meccanismi motori del linguaggio espressivo, nonché la fornitura di pensiero "creativo" o astratto.
Il lobo frontale può essere diviso in cinque regioni:
- il motore,
- premotore,
- visivo motorio frontale,
- Area motoria del discorso di Broca
- e l'area associativa frontale
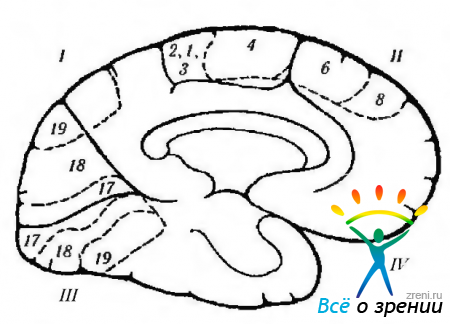
Riso. 4.1.12. Rappresentazione schematica delle funzioni della corteccia grande cervello, localizzati sulla superficie mediale e la loro violazione durante varie malattie. accompagnato da eccitazione o distruzione della corteccia (secondo Crosby et al., 1962): 1 - lobo parietale. Giro centrale (4). Sinistra regione occipitale: -alessia senza agrafia. Corteccia posteriore: -difficoltà a tradurre lo sguardo; - Sindrome di Balint (bilaterale), 11 - lobo frontale. Campo motorio aggiuntivo (6): + alzare la mano opposta, girare gli occhi e la testa verso la mano; + contrazione della pupilla; + ritardo nel parlato. Campo visivo motorio frontale (8). Giro della cintura: + espansione della pupilla; + lacrimazione. superficie orbitale. III - lobo occipitale. corteccia visiva primaria. Visione controlaterale: - emianopsia destra. Lobo IV-temporale: paralisi dei movimenti oculari; emiplegia ipsilaterale; idrocefalo; rigidità decerebrale; allucinosi peduncolare; emianopsia controlaterale; sindrome di Parnaud. Sotto la superficie: - riconoscimento facciale. Nota. I campi citoarchitettonici corrispondono alla classificazione di Brodmann. Più indica il risultato della stimolazione elettrica della corteccia e meno il risultato della distruzione del campo
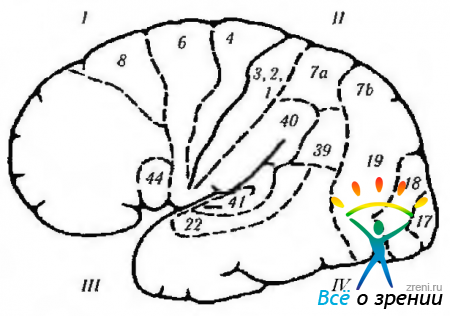
Riso. 4.1.13. Superficie laterale del cervello (le designazioni sono simili a quelle fornite nella figura precedente): 1 - lobo frontale. Campo motorio (4). Campo premotorio (6). Campo oculare motore (8): + saccade controlaterale; + chiusura dell'occhio opposto; + deviazione gomolaterale amichevole dell'occhio. Campo del linguaggio motorio (44): - afasia motoria (motoria). 11 - lobo parietale: - (- allucinosi semplice; - illusioni; - metamorfopsia: - perdita di attenzione; - agnosia; - aprassia; - indebolimento del nistagmo optocinetico; - emianopsia del quadrante inferiore. Giro sopramarginale (40): - amnesia; - gergo afasia Giro angolare (39): - agrafia - acalculia - perdita della capacità di distinguere tra destra e sinistra lato destro; agnosia delle dita. sensibilità somatica. III-lobo temporale: + allucinazioni visive; + dejà vu; + senso dell'olfatto compromesso; + convulsioni; + automatismi; - quadrianopsia superiore; -perdita di memoria; - Sindrome di Klüver-Bucy. corteccia uditiva primaria. Corteccia uditiva secondaria: -afasia uditiva. Planum temporale: - dislessia. IV-lobo occipitale (17, 18, 19): + sensazione di luce; -(-sensazione di colori; -(-sensazione di movimento; + sensazione di immagini lineari. Campi visivi associativi (18, 19): -(-movimenti oculari controlaterali come l'inseguimento regolare; + movimenti di vergenza; - sindrome di Anton
Area motoria primaria(area motoria; 4° campo secondo Brodman).
regione motoria b è davanti al solco centrale che separa il lobo frontale dal lobo parietale. È costituito da un gran numero di cellule piramidali giganti di Betz.
Rappresenta i movimenti, non i singoli muscoli. Le cellule che controllano i movimenti sono posizionate in modo tale che i movimenti della metà inferiore del corpo siano presentati nella parte superiore del giro e della metà superiore in quella inferiore. Gli assoni neuronali scendono attraverso la capsula interna e i peduncoli del cervello sotto forma di tratti corticobulbari e corticospinali.
Rimozione della corteccia motoria porta alla paralisi.
Zona premotoria(campo b) si trova direttamente anteriore al primario corteccia motoria, che occupa la parte posteriore dei tre giri frontali orientati orizzontalmente.
Forma le fibre dei tratti frontopontocerebellari e cede numerose fibre anche ai gangli della base.
Si suggerisce che la corteccia premotoria sia coinvolta nella pianificazione delle azioni motorie. Ciò è dimostrato dal fatto che solo quando si pensa all'azione prevista, i neuroni del sesto campo si eccitano. La stimolazione di questi neuroni provoca il sollevamento del braccio lato opposto, e la testa e gli occhi si girano nella direzione dell'arto sollevato. Le pupille si dilatano e il linguaggio è compromesso (vedi fig. 4.1.12, 4.1.13).
La distruzione di quest'area è accompagnata atassia pseudocerebellare del lato opposto. Anche la capacità di copiare i movimenti appresi (aprassia) è compromessa. In questo caso i movimenti individuali non vengono danneggiati, ma il paziente non è in grado di eseguirli nella sequenza desiderata.
Area visiva motoria frontale(campo 8 secondo Brodman).
Quest'area (campo 8 e parzialmente 6 e 9) occupa la parte posteriore del giro frontale mediale (gyrus frontalis medius).
Area visiva motoria frontale importante nei movimenti oculari volontari. Alcune aree definiscono convergenza, divergenza e accomodamento.
Va notato che durante la stimolazione elettrica è possibile rilevare il movimento degli occhi, della pupilla e delle palpebre in quasi tutte le aree degli emisferi cerebrali. La stimolazione elettrica dell'area motoria frontale dell'occhio porta a movimenti saccadici, in contrasto con la risposta alla stimolazione dei campi 18, 19 e 22 (movimenti di tracciamento graduali e di vergenza). La stimolazione dell'8o campo nelle scimmie è accompagnata da una concomitante deviazione controlaterale degli occhi. In questo caso, la pupilla si espande e la testa e il busto girano nella direzione opposta. Il danno al lobo frontale provoca una perdita reversibile della capacità di movimenti saccadici controlaterali.
Questa regione della corteccia è caratterizzata da numerose connessioni.
Fibre afferenti entrano nella corteccia visiva frontale dal tubercolo visivo, così come dalla corteccia temporale superiore, peristriatale e parietale (area 7). Qui sono stati trovati i neuroni che controllano i movimenti oculari.
Efferenti proiettata ai gangli della base, al talamo, alla regione pretettale (incluso il nucleo del tratto ottico), ai collicoli superiori dei quadrigemini e a parte formazione reticolare ponte.
A sua volta è da notare che le aree visive frontali vengono proiettate sulle aree frontali controlaterali e sulle aree corticali dallo stesso lato. Forniscono la percezione visiva (sul 7° campo corteccia parietale e corteccia peristriatale).
Quando proiettato su mesencefalo Le fibre della corteccia prefrontale attraversano la parte anteriore della capsula interna e si dividono in vie dorsali e ventrali nel diencefalo rostrale.
La via transtalamica dorsale attraversa il nucleo dorsomediale e intralamylare del talamo ottico e la parte mediana del cuscino, formando contemporaneamente non un gran numero di connessioni sinaptiche. Le sinapsi si trovano anche nei nuclei pretettali e nei tubercoli superiori dei quadrigemini.
Via peduncolo-tegmentale ventrale passa nella parte più ventrale del tronco cerebrale, fornendo fibre ai nuclei dell'ipotalamo e più in profondità - ai tubercoli superiori dei quadrigemini. Allo stesso tempo, le aree visive frontali hanno una proiezione topografica sui tubercoli superiori dei quadrigemini. Il percorso termina nella formazione reticolare del ponte.
Terzo legamento prefrontale si verifica nella regione della giunzione diencefalico-mesencefalica situata vicino al nucleo rosso. Questo percorso si proietta sul nucleo del fascicolo longitudinale mediale e sul nucleo intermedio di Cajal dallo stesso lato, nonché sul nucleo di Darkshevich e sulla parte rastrale del nucleo oculomotore, su entrambi i lati.
Varie aree della corteccia visiva frontale controllano i movimenti oculari saccadici. diversa ampiezza, così come la direzione delle saccadi. Forniscono la cosiddetta "attenzione visiva", indirizzando gli occhi verso l '"obiettivo".
Attualmente è oggetto di indagine il ruolo del lobo frontale nel funzionamento delle palpebre. La chiusura volontaria delle palpebre richiede il rilassamento dell'elevatore palpebrale e la contrazione del muscolo orbicolare dell'occhio. C'è controversia riguardo alla localizzazione delle aree del cervello che controllano i movimenti volitivi della palpebra. Questi siti possono essere localizzati nelle aree motorie premotorie o visive motorie frontali. La stimolazione di quest'area porta alla chiusura della palpebra controlaterale. Il danno unilaterale ai lobi frontali non dominanti può essere accompagnato dalla mancata chiusura bilaterale delle palpebre.
Area motoria del linguaggio di Broca(campi 44 e 45 secondo Brodman).
Quest'area si trova nella parte postero-inferiore del lobo frontale. Contribuisce all'inizio del discorso. Il danno all'area di Broca porta all'afasia, che consiste nella difficoltà di collegare una certa sequenza di parole per fornire un discorso correlato (afasia espressiva). I pazienti comprendono sia la lingua scritta che quella parlata, ma di solito non riescono a pronunciare nulla. Anche la capacità di scrivere è compromessa, sebbene la mano possa essere utilizzata per altre attività.
area associativa frontale. Questa importante area nella metà anteriore del lobo frontale è responsabile dell'avvio di tutti i tipi di funzioni comportamentali-intellettuali, viscerali ed emotive. Cambiamenti nella memoria, nell'intelligenza e nella personalità si verificano con irritazione o massiccia distruzione dei lobi frontali. Ciò è particolarmente pronunciato in questi casi. quando è interessato l’emisfero dominante.
Tratto olfattivo si trova sotto la superficie orbitale del lobo frontale ed è l'unica via sensoriale che raggiunge la corteccia senza la formazione di sinapsi nel talamo. Tumore unilaterale della superficie orbitaria del lobo frontale (meningioma del solco olfattivo o meningioma dell'ala minore) sfenoide) può portare alla compressione del tratto olfattivo e nervo ottico. Di conseguenza si sviluppano anosmia (mancanza di olfatto), atrofia del nervo ottico ipsilaterale e gonfiore della testa del nervo ottico sul lato opposto (sindrome di Kennedy Foster).
I danni alle strutture del sistema limbico all'interno del lobo frontale, in particolare al giro del cingolo, possono causare disturbi nell'aspetto emotivo della vita, così come segni viscerali, tra cui la lacrimazione senza causa e cambiamenti nella risposta pupillare agli stimoli luminosi.
Lobo parietale(vedi fig. 4.1.12, 4.1.13). In generale, la funzione dei lobi parietali è associata alla sensibilità, alla memoria legata alla parola e all'apprendimento, nonché all'orientamento del corpo nello spazio attraverso l'elaborazione delle informazioni visive.
Nel lobo parietale anteriore c'è un giro situato parallelo al giro precentrale o corteccia motoria. Questo è il giro postcentrale, o corteccia somatosensoriale primaria (campi 3, 1 e 2) (vedi Fig. 4.1.13). Il resto del lobo parietale può essere diviso in due lobuli: superiore e inferiore. I campi 5 e 7, situati sulla superficie del lobulo parietale superiore, sono associati alla correlazione delle informazioni somatosensoriali, che consentono di valutare consapevolmente il peso, le proprietà superficiali, le dimensioni e la forma di un oggetto.
Lobulo parietale inferiore(giro sopramarginale e angolare) è strettamente correlato ai meccanismi del linguaggio, poiché la distruzione di quest'area della corteccia nell'emisfero sinistro porta a una compromissione della percezione del linguaggio.
Il ruolo del lobo parietale nell'orientamento spaziale non è stato ancora studiato. Tuttavia, nei pazienti con sconfitta, si nota una violazione di questa funzione.
Le fibre afferenti passano attraverso il lobo parietale, collegando le aree 18 e 19 con i lobi frontali e il tronco encefalico. Questi percorsi includono i fasci longitudinali superiori e inferiori.
Si presume che la corteccia parietale (campo 7) fornisce la funzione di "attenzione" ai bersagli visivi.
Il settimo campo riceve afferenze da giro del cingolo(g. cinguli), dalla regione pretettale e dai tubercoli superiori dei quadrigemini attraverso i nuclei del talamo. Si trovano anche afferenze che vanno dall'area visiva frontale e dalla corteccia prestriata alla corteccia del lobulo parietale.
Le efferenze vengono proiettate sulla regione pretettale, sui tubercoli superiori dei quadrigemini, sulla materia grigia situata attorno all'acquedotto silviano e sui lobi parietali dei due emisferi del cervello.
Quando gli occhi si muovono, vengono attivati i neuroni del lobo parietale inferiore, il che indica il loro ruolo nel controllo dei movimenti oculari volontari. L'attività continua per tutta la durata dell'osservazione del target.
Le cellule del 7° campo non determinano i movimenti saccadici, poiché la loro attività non aumenta durante le saccadi.
Stimolazione del giro angolare(g. angularis) porta negli animali da esperimento ad una deviazione amichevole degli occhi. Questi sono i campi 38 e 39, che passano attorno al bordo posteriore del giro temporale medio (g. temporalis medius). Rimozione operativa questa zona della corteccia non porta a movimenti oculari alterati. Il danno bilaterale alla corteccia parietale può portare allo sviluppo dell'aprassia motoria dell'occhio.
La sconfitta del lobo parietale è accompagnata anche da una violazione dei meccanismi motori visivi che forniscono il nistagmo optocinetico, durante il quale gli occhi producono saccadi per cercare e fissare il bersaglio.
Altri segni motori di lesioni isolate del lobo parietale includono deviazione amichevole degli occhi nella direzione opposta rispetto al sito della lesione con chiusura forzata delle palpebre, scarsa mobilità del paziente e scarso controllo (incomprensione) degli arti del lato opposto. Le difficoltà nel comprendere la natura dell'azione degli arti portano all'aprassia costituzionale e oculomotoria. I pazienti con danni al lobo parietale possono avere difficoltà a spostare lo sguardo da un oggetto visivo a un altro nella metà controlaterale del campo visivo.
Un altro segno di disfunzione del lobo parietale è incapacità di fissare un oggetto visivo.
Oltre alle funzioni sensoriali, che sono localizzate nel labbro posteriore del solco centrale di Rolando, il lobo parietale è sede di un complesso processo di riconoscimento e comprensione. Il danno diffuso ai lobi parietali termina con agnosia, disattenzione visiva, metamorfopsia, fotopsia, allucinazioni, illusioni e alessia (vedi Fig. 4.1.12, 4.1.13).
Di più dettagli sulle conseguenze della disfunzione del lobo parietale sono riportate nella letteratura neurologica. Qui presentiamo solo alcune delle sindromi.
agnosia visiva, che spesso si sviluppa in violazione della circolazione sanguigna nel sistema nervoso centrale, è caratterizzata da una violazione del processo di riconoscimento. L'agnosia può essere completa o parziale. L'agnosia visiva viene diagnosticata nei casi in cui il paziente può identificare un oggetto solo tattilmente. Il riconoscimento e la comprensione delle immagini visive e il significato cromatico dell'immagine sono spesso compromessi.
Il danno bilaterale al lobo parietale (più comune nella demenza senile, rammollimento delle aree temporo-occipitale e parietale della corteccia, lesioni delle sezioni posteriori di entrambi gli emisferi, encefalite) sono accompagnati da agnosia visiva e atassia visiva (sindrome di Balint).
Agnosia visuo-spaziale caratterizzato da difficoltà nel valutare la dimensione dell'oggetto e la distanza da esso. Si nota anche l'emergere di una soluzione errata di problemi spaziali "sintetici" come la lettura di una mappa. L'agnosia simultanea si riduce al fatto che i pazienti mantengono la capacità di riconoscere i singoli oggetti, ma non hanno la capacità di percepire un gruppo di oggetti (immagini) nel suo insieme.
Asterognostia caratterizzato dal fatto che il paziente può analizzare gli oggetti dal loro aspetto, ma non è in grado di caratterizzarli tattilmente. Ciò si sviluppa a seguito dell'interruzione della complessa interazione tra informazioni tattili e visive nei lobi parietali della corteccia.
Lesioni diffuse dei lobi parietali possono portare a illusioni e metamorfopsie. Queste distorsioni includono micropsia, macropsia. I pazienti notano il movimento di bersagli fissi, stimano erroneamente la distanza dagli oggetti. A tipi speciali le illusioni includono la papinopsia (violazione della percezione visiva sotto forma di conservazione o ricomparsa di un'immagine visiva dopo la scomparsa di un oggetto dal campo visivo), poliopia allucinatoria, ecc.
Le illusioni si sviluppano a seguito di disturbi diffusi della corteccia e allucinazioni e fotopsie si sviluppano con lo sviluppo di un tumore o di un danno tossico alla corteccia.
Le caratteristiche della patologia del lobo parietale possono anche dipendere dal fatto che il focus patologico sia localizzato nell'emisfero dominante (di solito sinistro) o non dominante. Il lobo parietale dominante è responsabile dello sviluppo di disturbi del linguaggio come l'afasia sensoriale uditiva, agnosia visiva e agrafie.
Danno al giro angolare dominante(più spesso a causa di disturbi circolatori nel bacino dell'arteria angolare) sviluppano nistagmo, disturbi visivo-agnostici, agrafia, acalculia (incapacità di aggiungere o sottrarre numeri), agnosia digitale, incapacità di distinguere tra il lato destro e quello sinistro ( sindrome di Gerstmann; sindrome del giro angolare). Forse lo sviluppo dell'emianopsia omonima.
Se il giro angolare non dominante è danneggiato, l'interazione del paziente con l'ambiente viene interrotta. Di conseguenza, si sviluppano disorientamento visivo e agnosia topografica.
Anteriormente al giro angolare e attraverso il solco sopratemporale si trova giro sopramarginale. Nell'emisfero dominante, quest'area è il punto d'incontro delle componenti visiva e uditiva del linguaggio.
Il danno alle aree associative parietali posteriori spesso termina sviluppo di aprassia(difficoltà nell'eseguire movimenti specializzati "fini"). Ciò, apparentemente, è dovuto a un'incomprensione della sequenza di movimenti necessaria per eseguire una particolare funzione (vale a dire, non è chiaro al paziente cosa e in quale sequenza dovrebbe essere eseguito). Il paziente non ha la capacità di disegnare un diagramma semplice (aprassia costruttiva). Una forma meno grave di questa patologia è l'agnosia tattile (incapacità di riconoscere un oggetto alla palpazione). A volte questa malattia è chiamata asterognostia. Allo stesso tempo, non ci sono segni di perdita della sensibilità tattile o propriocettiva e l'integrazione delle informazioni visive e somatosensoriali è compromessa. L'aprassia e l'asterognostia si sviluppano spesso dopo un danno all'emisfero sinistro del cervello. L'asterognostia è solitamente limitata al lato opposto del corpo, mentre l'aprassia è solitamente bilaterale.
Le informazioni visive e uditive sono strettamente connesse con la funzione del linguaggio. Per questo motivo, il danno alle aree 39 e 40, così come a parti dell’area associativa uditiva, solitamente provoca afasia, nota come afasia Wernicke(Wernicke). Se il danno è limitato al giro angolare, si perde la capacità di comprendere la lingua scritta (alessia) e la capacità di scrivere (agrafia). Allo stesso tempo, i pazienti comprendono il discorso. Questa condizione di solito si sviluppa dopo un infortunio.
L'integrazione delle informazioni visive e somatosensoriali è importante per la formazione di una "immagine corporea", cioè per comprendere le caratteristiche della posizione del corpo nello spazio. Danni estesi ai campi 7, 39 e 40 di un emisfero possono portare allo sviluppo di "incomprensioni" o "negligenza" della metà opposta del corpo. I pazienti non sono in grado di fasciare o sciogliere i propri arti. Inoltre non riconoscono i propri arti.
Lobo occipitale(campi 17, 18 e 19). Lungo il solco dello sperone (fissura calcarina) si trova l'area visiva primaria (campo 17). Adiacente ad essa si trova l'area visiva "valutativa" (campo 18), che si ritiene converta i segnali ricevuti dal campo 17 in immagini visive.
L'area 17 della corteccia occipitale è zona di ricezione ed elaborazione delle informazioni visive e ha collegamenti importanti con le aree motorie visive della corteccia. È a questo livello della corteccia visiva che viene valutato il significato di ciò che si vede. L'irritazione di quest'area provoca allucinazioni e immagini oniriche.
Meritano attenzione anche le connessioni motorie del lobo occipitale perché sono coinvolte nel nistagmo optocinetico, nel riflesso di accomodazione e nella regolazione della visione binoculare.
I campi 18 e 19 sono interdipendenti e hanno più connessioni con il resto del cervello. Si ritiene che l'area 18 sia più interessata all'integrazione delle informazioni visive. ricevuto nel 17° campo, mentre il campo 19 traduce queste informazioni in attività mentale e motoria più complessa.
Anatomiche recenti e ricerca fisiologica ha rivelato che ci sono almeno, 10 sottoregioni dei campi 18 e 19, i cui neuroni hanno la capacità di rispondere agli stimoli visivi. Ciascuna di queste sottoregioni è associata al 17° campo. Tuttavia, non sono correlati tra loro. Il danno a queste sottoregioni non porta alla cecità. Alcuni sono violati funzioni visive, come l'analisi del colore, la percezione dell'integrità di un oggetto, ecc.
corteccia associativa visiva, localizzato in alcune aree del lobo temporale, è coinvolto anche nell'elaborazione delle informazioni visive. Ciò vale soprattutto per il campo 21 (nella corteccia temporale media). È questo campo che fornisce il riconoscimento dell'immagine e il suo danno termina con l'agnosia.
Le proprietà della corteccia visiva di cui sopra sono fornite anche da numerose connessioni orizzontali e verticali tra i neuroni in diversi strati della corteccia visiva. È grazie alla loro presenza che si nota l'eccezionale plasticità della corteccia visiva nell'elaborazione di informazioni visive di vario tipo.
L'organizzazione strutturale della corteccia visiva e le sue funzioni saranno descritte più in dettaglio nella sezione successiva.
Lobo temporale(campi 41, 22).
Questa parte del cervello associato alla percezione delle sensazioni uditive, partecipa alla funzione vocale attraverso il controllo uditivo della parola e svolge anche un ruolo nella valutazione dello spazio e della funzione della memoria. È stato stabilito da tempo che il giro temporale trasversale (campo 41) è il centro uditivo primario. Campo piccolo (22). che circonda questo giro, è considerato "mentale", o "valutativo", centro uditivo. L'irritazione della maggior parte del lobo temporale, specialmente lungo il giro temporale medio, provoca sensazioni che il paziente descrive come uditive. I pazienti con danni al "campo mentale" del lobo temporale talvolta perdono la capacità di valutare i toni sonori. Allo stesso tempo, il paziente non riconosce i motivi e i suoni musicali vengono percepiti da lui come rumori casuali.
Anche il campo 22 dell'emisfero sinistro è associato alla funzione vocale, poiché il danno a quest'area porta alla perdita della capacità di comprendere il significato delle parole. Esistono anche prove che il lobo temporale è associato alla sensibilità "vestibolare" (senso di equilibrio), poiché la stimolazione del giro temporale superiore posteriore in un paziente cosciente provoca vertigini e senso di rotazione.
Lesioni profonde nel lobo temporale spesso coinvolgono le fibre ottiche più basse provenienti dal corpo genicolato laterale. In cui deficit visivo. La più comune è l’emianopsia del quadrante superiore.
Con la sconfitta del lobo temporale o psicomotorio si sviluppa epilessia accompagnata da allucinazioni visive, la sensazione di deja vu e deja pense (“già visto” - fr.), in cui l'esperienza visiva e i pensieri hanno uno strano rapporto amichevole, compaiono automatismi che accompagnano l'amnesia, il comportamento compulsivo, la rabbia e l'ansia.
Nei pazienti con danno unilaterale al primario corteccia uditiva lieve perdita dell'udito. In violazione del riconoscimento uditivo (agnosia uditiva), i campi uditivi associativi sono interessati. I pazienti di solito riferiscono di poter sentire i suoni ma di non riuscire a identificarli.
Un'area coinvolta nel riconoscimento facciale si trova sulla superficie ventrale del lobo temporale. Danno a quest'area (più spesso in violazione della circolazione sanguigna nella regione dell'arteria basilare del cervello), che, di regola, si estende al lobo occipitale adiacente (campi 20, 21), entrambi gli emisferi, accompagnato dallo sviluppo della prosopagnosia(impossibilità di riconoscere i volti). In generale, questa condizione non è accompagnata da altri segni neurologici. Il paziente può leggere e nominare gli oggetti. Nei casi in cui il paziente sente la voce di una persona familiare, può immediatamente identificarla.
La restante parte del giro temporale superiore è occupata dall'area 22 (corteccia associativa uditiva), che riceve un gran numero di afferenze dai campi 41 e 42 per fornire fibre ai lobi parietali e occipitali. L'area 22 è coinvolta anche nelle funzioni linguistiche. I danni alla parte superiore del 22° campo portano a difficoltà nella comprensione della lingua.
I lobi temporali sono particolarmente importanti nella funzione della memoria.. Ad esempio, la stimolazione della corteccia associativa uditiva nei pazienti durante interventi neurochirurgici evoca ricordi complessi, sia uditivi che visivi. Con danni estesi al lobo temporale, si verifica un deterioramento della memoria.
lobo limbico(Fig. 4.1.14).
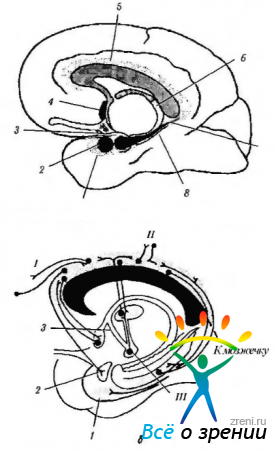
Riso. 4.1.14. Sistema limbico (secondo Brodsl, 1992): a - la posizione delle strutture del sistema limbico (1 - campo entoriale; 2 - amigdala; 3 - uncino; 4 - nucleo settale; 5 - giro cingolato; b - fornice; 7 - ippocampo; 8 - giro paraippocampale); b - connessioni del giro cingolato (I - corteccia associativa frontale; II - corteccia associativa parietale e temporale; III - corpo mastoideo). Il giro cingolato è collegato ai campi associativi della corteccia cerebrale e del sistema limbico, fornendo interazione tra loro.
Il sistema limbico è un complesso di formazioni del terminale, del diencefalo e del mesencefalo. In realtà " sistema limbico" coincide con il concetto precedentemente utilizzato: "cervello olfattivo".
Se la nuova corteccia (neocorteccia) controlla le relazioni spazio-temporali dell’organismo con l’ambiente, ed è anche responsabile del pensiero logico-formale e delle capacità stereognostiche, allora il sistema limbico determina le funzioni emotive e la motivazione all’azione così come i processi di apprendimento e di memoria. Controlla anche l'omeostasi. La sua funzione importante è l'organizzazione delle reazioni comportamentali dell'individuo in risposta all'influenza dell'ambiente esterno e ai cambiamenti nell'ambiente interno del corpo. Queste reazioni hanno lo scopo di preservare l'individuo e sono mediate dai centri inferiori del diencefalo.
Il lobo limbico ha qualcosa a che fare anche con la funzione dell'olfatto, poiché l'irritazione o il danno alla regione dell'ippocampo sono accompagnati dall'olfatto.
Si forma il lobo limbico:
- area subcausale (area subcallosa),
- giro del cingolo (gyrus cinguli),
- istmo del giro cingolato
- giro paraippocampale (giro parahippocampalis),
- ippocampo (corno di ammonio, giro dentato e base dell'ippocampo o subiculum)
- e il legamento diagonale di Broca.
Le strutture sottocorticali del sistema limbico comprendono le tonsille (corpo amigdaloideo), i nuclei del setto e il nucleo talamico anteriore.
Le connessioni afferenti ed efferenti delle strutture del sistema limbico, sia tra loro che con altre parti del cervello, sono estremamente diverse. Le sue connessioni reciproche con l'ipotalamo sono più pronunciate.
Ipotalamo e corpi mammillari collegato all'ippocampo e alla regione del setto attraverso il fornice, all'amigdala attraverso la stria terminale e il fascio amigdalofuga, e al cervello olfattivo attraverso il fascio mediale prosencefalo. Il sistema limbico è collegato al mesencefalo attraverso l'ipotalamo e i corpi mammillari.
Il sistema limbico comunica con la neocorteccia nella parte frontale e Lobo temporale. Regioni temporali sono i principali responsabili della trasmissione delle informazioni dalla corteccia visiva, uditiva e somatica all'amigdala e all'ippocampo. Le regioni frontali fungono da divisione principale della neocorteccia, regolando le zone limbiche.
Gangli della base rappresentano un accumulo sottocorticale di neuroni sotto forma di un gruppo di nuclei (Fig. 4.1.15; 4.1.16).
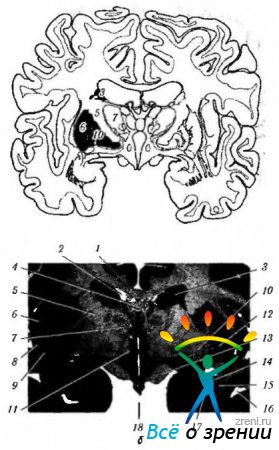
Riso. 4.1.15. Gangli della base: a - una rappresentazione schematica della posizione delle strutture legate ai gangli della base; b - sezione frontale del cervello che passa attraverso le strutture relative ai gangli della base (4 - corpo calloso; 2 - corno ventricolo laterale; 3- corpo del nucleo caudato; 4 volte; 5 capsule interne; 6 - isolotto; 7 - tubercolo visivo; 8- conchiglia; 9 - recinzione; 10 - palla pallida; 11 - regione ipotuberosa; 12 - sostanza senza nome: 13 - tratto visivo; 14 - parte inferiore posteriore del nucleo caudato; 15 - tonsilla: 16 - corno temporale del ventricolo laterale; 17 - corno frontale del ventricolo laterale; 18 - terzo ventricolo)
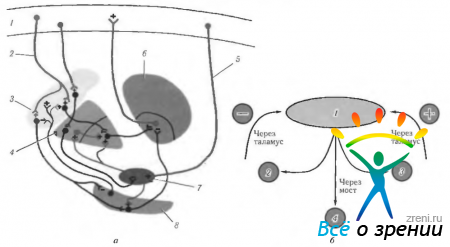
Riso. 4.1.16. Le più importanti connessioni afferenti, efferenti e interne dei gangli della base e la loro connessione con il cervelletto (spiegazione nel testo): a - (corteccia 1-motoria; 2 - fibre efferenti dalla corteccia allo striato; 3 - striato; 4 - palla pallida; 5 - fibre afferenti; 6 - tubercolo visivo; 7 - nucleo subtalamico; 8 - substantia nigra); b - (1 - corteccia cerebrale (motoria); 2 - gangli della base; 3 - cervelletto; 4 neuroni motori e intercalari)
Controllano il movimento del corpo. Questa funzione dei gangli della base è stata stabilita sulla base di osservazioni cliniche. Quando sono danneggiati, c'è una violazione attività motoria sia sotto forma di incapacità di avviare i movimenti necessari, sia di incapacità di sopprimere i movimenti.
Nella malattia di Parkinson, che si verifica quando i gangli della base sono danneggiati, i pazienti hanno un volto "inespressivo". Ciò è associato ad una ridotta attività motoria dei muscoli facciali e al controllo dei movimenti oculari, in particolare ai disturbi dei movimenti saccadici. È per questo motivo che esamineremo più da vicino la neuroanatomia di quest'area.
Ce ne sono tre principali accumuli di nuclei sottocorticali, che sono chiamati striato ( corpo striato), recinto (claustrum) e amigdala (corpus amigdaloideum).
- Il corpo striato è costituito da due parti: il nucleo caudato (nucleo caudatus) e il nucleo lentiforme (nucleo lentiformis).
Nucleo caudato si trova sopra e medialmente al nucleo lenticolare, separato da quest'ultimo da uno strato di sostanza bianca chiamato capsula interna (capsula interna). La parte anteriore ispessita del nucleo caudato, la sua testa (caput nuclei caudati) formano la parete laterale corno anteriore ventricolo laterale, mentre la sezione sottile posteriore del nucleo caudato (corpus et cauda nuclei caudati) si estende indietro lungo il fondo della parte centrale del ventricolo laterale. Sul lato mediale, il nucleo caudato è adiacente alla collinetta visiva, separato da essa da una striscia di sostanza bianca (stria terminalis). Anteriormente e inferiormente, la testa del nucleo caudato raggiunge la sostanza perforata anteriore (substantia perforata anterior). A questo punto la testa è collegata al nucleo lenticolare (con una parte chiamata putamen). Oltre a questa ampia connessione di entrambi i nuclei sul lato ventrale, sono presenti anche sottili strisce di sostanza grigia intervallate da ciuffi bianchi della capsula interna. Hanno dato origine al nome "corpo striato" (corpus striatum).
Nucleo lenticolare(nucleo lentiformis) si trova lateralmente al nucleo caudato e alla collinetta visiva, separato da essi da una capsula interna. Su una sezione orizzontale dell'emisfero, la superficie mediale del nucleo lentiforme, rivolta verso la capsula interna, ha la forma di una fragilità con l'apice rivolto verso il centro. Il lato anteriore dell'angolo è parallelo al nucleo caudato e il lato posteriore è parallelo al talamo. La superficie laterale è leggermente convessa e si affaccia sul lato laterale dell'emisfero nella regione dell'insula. Anteriormente e ventralmente, come già indicato, il nucleo lentiforme si fonde con la testa del nucleo caudato.
Nella sezione frontale il nucleo lentiforme ha la forma di un cuneo, il cui apice è rivolto verso il lato mediale, e la base è lateralmente. Il nucleo lenticolare è diviso in tre segmenti da due strati bianchi paralleli (lamine midollari). L'area grigio scuro situata lateralmente è chiamata conchiglia (putamen), e le due mediali, più chiare, insieme sono chiamate palla pallida (globus pallidus).
Differente già nel suo aspetto macroscopico, palla pallida(globus pallidus) ha anche una struttura istologica, che differisce significativamente dalla struttura di altre parti dello striato.
Alla luce di tutte queste caratteristiche, il globo pallido viene distinto in una speciale unità morfologica chiamata pallido, mentre la designazione striato è lasciata solo per il putamen e il nucleo caudatus. Di conseguenza il termine "nucleo lenticolare" perde il suo significato originario e può essere utilizzato solo in senso puramente topografico. Allo stesso tempo, invece del vecchio nome corpo striato, i nuclei caudato e lenticolare sono chiamati sistema striopallidar.
Il sistema striopallidario è la parte principale del sistema extrapiramidale e inoltre è il centro regolatore più alto funzioni autonome in relazione alla termoregolazione e al metabolismo dei carboidrati, dominando su centri vegetativi simili nell'ipotalamo.
- Recinzione(claustrum) è una sottile lamina di materia grigia, adagiata nella regione dell'isola, tra questa e il putamen. È separata da quest'ultima da uno strato di sostanza bianca chiamato capsula esterna (capsula esterna).
- amigdala(corpus amygdaloideum) si trova sotto il putamen all'estremità anteriore del lobo temporale. L'amigdala sembra appartenere ai centri olfattivi sottocorticali e al sistema limbico. Termina con il fascio di fibre provenienti dal lobo olfattivo e termina con la sostanza perforata anteriore (substantia perforata anterior), notata nella descrizione della collinetta visiva chiamata stria.
Il ruolo dell'amigdala negli esseri umani non è ben compreso. Fondamentalmente, il ruolo dell'amigdala è stato rivelato sulla base di uno studio su pazienti neurologici. Grazie a questi studi è stato stabilito che la stimolazione meccanica o elettrica dell'amigdala nell'uomo porta spesso alla paura o ad altre reazioni emotive. Quindi, l'amigdala umana partecipa ai processi di percezione emotiva. È stato anche dimostrato che con un danno bilaterale alle tonsille si verifica una perdita di memoria "emotiva", la comparsa di un'espressione facciale "spaventata" e una violazione dell'integrazione delle informazioni olfattive e visive.
È noto che una persona ha la capacità di determinare l'umore e lo stato emotivo di un altro individuo. IN Ultimamente coinvolgimento dell'amigdala in questo complesso processo fisiologico. Nel determinare lo stato emotivo di una persona, il nostro cervello cattura e analizza i sottili cambiamenti nell'espressione facciale e, prima di tutto, nella direzione dello sguardo. I meccanismi neurali alla base di questi processi non sono esattamente chiari. Tuttavia, è accertato che l’impostazione “accurata” dello sguardo, valutata psicofisiologicamente come presenza di “attenzione”, determinato superiore corteccia temporale e amigdala. Nello studio di pazienti con autismo, è stato rivelato che spesso presentano un'anomalia dell'amigdala o un suo danno. I meccanismi immediati alla base del giudizio sociale di altre persone basato sull’orientamento dello sguardo rimangono in gran parte sconosciuti.
Recentemente, anche la substantia nigra e i nuclei subtalamici sono stati riferiti ai gangli della base. Inoltre, nella substantia nigra si distinguono 2 parti: reticolare (pars reticulata) e compatta (pars compacta).
I metodi morfologici ed elettrofisiologici lo hanno dimostrato la maggior parte i segnali afferenti che arrivano ai gangli della base entrano nello striato (vedi Fig. 4.1.16). Questi segnali provengono da molte fonti, le principali sono:
- tutte le aree della corteccia emisferi;
- nuclei intralamellari del talamo
- substantia nigra (lungo la via dopaminergica).
Dall'interno del globo pallido ha origine il più importante di tutti vie efferenti gangli della base. Questo percorso termina principalmente nel tubercolo visivo e nel tetto del mesencefalo.
Pertanto, i gangli della base svolgono il ruolo di anello intermedio nella catena che collega le aree motorie della corteccia con tutte le altre aree. La loro funzione principale è “pianificazione” dell’attività fisica.
IN l'anno scorso hanno scoperto che anche i gangli della base insieme ad alcune aree corticali ne determinano alcune funzioni cognitive, compresa l'attenzione, la memoria. Il concetto di "attenzione", nel senso più ampio del termine, è un "processo di scelta". Considerando che il processo di orientamento nello spazio e la scelta dell'obiettivo di interesse, a cui prendono parte i movimenti oculari saccadici, è associato ai gangli della base, diventa chiaro un ruolo significativo del complesso basale nei processi di orientamento attraverso le saccadi. In questo caso, un posto speciale nel controllo delle saccadi dell'occhio è svolto dalla substantia nigra, in particolare dalla sua parte reticolare. Sia anatomicamente che elettrofisiologicamente è stata dimostrata la presenza della sua connessione con i tubercoli superiori dei quadrigemini. La funzione principale dei neuroni della substantia nigra è quella di inibire l'attività dei neuroni del collicolo superiore dei quadrigemini coinvolti nella generazione delle saccadi.
Nell'attività motoria degli occhi vi prende parte anche il nucleo caudato, che ha connessioni con la substantia nigra ed i tubercoli esterni dei quadrigemini. L'eccitazione del nucleo caudato porta a movimenti saccadici dell'occhio nella direzione opposta al nucleo eccitato.
Anche il nucleo subtalamico è coinvolto nel movimento oculare. Su questo nucleo viene proiettata una palla pallida, la corteccia della regione frontale. Le fibre partono dal nucleo subtalamico fino alla substantia nigra, i segmenti esterno ed interno del globo pallido.
La funzione del globo pallido nell'attività oculomotoria è meno chiara. Ciò è dovuto al fatto che il globo pallido presenta un gran numero di connessioni tra i vari nuclei dei gangli della base e poche connessioni con centri nervosi situato all'esterno dei gangli della base. I neuroni oculomotori sono stati trovati solo nella parte dorsale del globo pallido, cioè nella parte che riceve input dal nucleo caudato.
Il ruolo dei gangli della base nel movimento oculare confermato da numerose osservazioni cliniche. Nella malattia di Parkinson, accompagnata dalla degenerazione dei gangli della base e della materia grigia del mesencefalo e dall'interruzione delle vie frontomesencefaliche, si sviluppa una paralisi sopranucleare progressiva. In tali pazienti, la velocità dei movimenti oculari saccadici è ridotta. Le saccadi sono solitamente ipometriche, che sono associate a una violazione della funzione di "memoria saccadica". Spesso il movimento oculare non può essere completato nella sua interezza. I movimenti oculari verticali sono influenzati in modo più significativo rispetto a quelli orizzontali. I pazienti mostrano anche rigidità assiale e demenza. Cambiamenti simili sono stati riscontrati anche nella malattia di Huntington.
corpo calloso(corpo calloso). Il corpo calloso è un grande fascio di fibre (costituito da circa 250 milioni di fibre) che collega entrambi gli emisferi (Fig. 4.1.17).
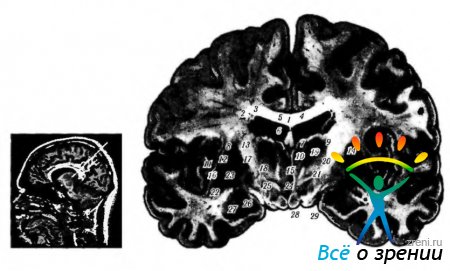
Riso. 4.1.17. Localizzazione dei nuclei del talamo: a - risonanza paramagnetica nucleare; b - sezione frontale del cervello (1 - corpo del corpo calloso; 2 - corpo del nucleo caudato; 3 - parte centrale del ventricolo laterale; 4 - setto trasparente; 5 - plesso coroideo ventricolo laterale; 6 - volta; 7 - gruppo anteriore dei nuclei del talamo; 8- capsula esterna; 9 - gruppo laterale dei nuclei del talamo; 10- nuclei mediali del talamo: - conchiglia; 12- placca midollare laterale; 13 - capsula interna; 14 - nucleo reticolare del talamo; 15 - picco intertalamico; 16 - parte laterale della palla pallida; 17 - placca midollare mediale; 18 - fascio mammillotalamico; 19 - Campo trote HI; 20 - gopa incerta; 21 - Campo trote H2; 22 - recinzione; 23-parte mediale della palla pallida; 24 - terzo ventricolo; 25 - nuclei dell'ipotalamo; 26-tratto visivo; 27 - amigdala; Corpo a 28 capezzoli; 29-base delle gambe del cervello)
La funzione principale di questo raggio è la trasmissione di informazioni da un emisfero all'altro.
Le principali funzioni del corpo calloso sono:
- Correlazione delle immagini nelle metà destra e sinistra del campo visivo.
- Integrazione delle sensazioni provenienti dagli arti accoppiati, necessaria per l'apprendimento della coordinazione motoria.
- Integrazione dei processi di attenzione e attivazione negli emisferi.
Nei pazienti in cui è stato reciso il corpo calloso per scopi terapeutici, si è verificata una disfunzione cerebrale Vita di ogni giorno non è stato osservato. Sono stati rivelati solo attraverso prove speciali. Altri pazienti talvolta sviluppano disturbi comportamentali piuttosto significativi. Ciò si manifesta molto spesso con "cecità verbale", "sordità verbale" e perdita di coordinazione dei movimenti degli arti destro e sinistro.
Continua nel prossimo articolo: Anatomia del cervello? Parte 3
E agli occhi di tutta la capitale
Il galletto svolazzava dal ferro da calza,
volò sul carro
E si sedette sulla corona del re,
Sorpreso, diede un becco alla corona
E salì alle stelle... e allo stesso tempo
Dadon cadde dal carro -
Ha sussultato una volta ed è morto.
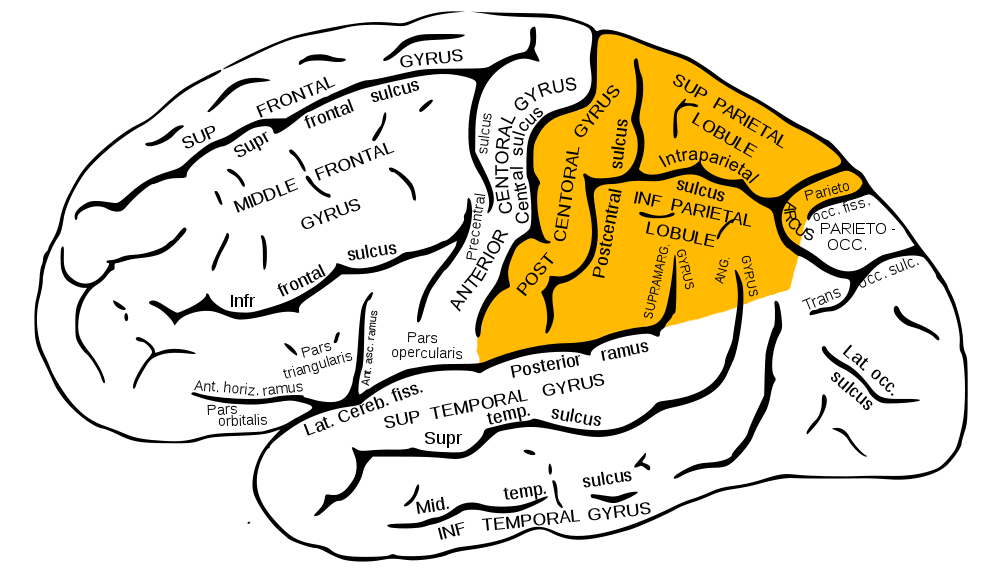
Sembra che tu abbia già capito cosa verrà discusso nel prossimo materiale del ciclo "Come funziona il cervello". Abbiamo già parlato dei lobi frontali, temporali e parietali, ora passiamo ai lobi parietali. Sono anche Lobus parietalis in latino, familiari ai medici.
Lobi parietali mostrati in giallo
Il lobo parietale si trova appena sopra l'occipite ed è "costituito" da tre circonvoluzioni: una verticale- posteriore centrale (la sezione più vecchia) e due orizzontali: parietale superiore (nuovo) e parietale inferiore (più recente).
Come la struttura dei lobi frontali, parti del corpo umano sono “proiettate” sulla circonvoluzione anteriore centrale del parietale: il terzo inferiore è il viso, il terzo medio è il braccio e il busto, quello superiore è la gamba. Non dimenticare che la quota è "raddoppiata", quindi metà di essa è responsabile di una metà (opposta) del corpo.
La struttura dei lobi parietali
Inoltre, nel giro parietale superiore ci sono centri responsabili tipi complessi sensibilità profonda: senso muscolo-articolare, bidimensionale-spaziale (definizione di numeri, lettere, figure disegnate con matita o altro oggetto contundente sulla pelle umana), senso di peso e volume di movimento, senso di riconoscere gli oggetti al tatto.
Nel lobo parietale inferiore ci sono centri di prassi, cioè movimenti che sono diventati "automatici" nel processo di ripetizione ed esercizio, che si sviluppano nel processo di apprendimento e pratica costante, ad esempio camminare, mangiare, vestirsi e Presto.
Il lobo parietale è coinvolto nell'elaborazione e nella percezione cosciente dei segnali somatosensoriali (dai recettori nei muscoli, nella pelle, nelle articolazioni e organi interni) informazioni che influiscono sui movimenti volontari.
Le lesioni del lobulo parietale superiore sono accompagnate dallo sviluppo di una violazione della capacità di riconoscere gli oggetti toccandoli con gli occhi chiusi. I pazienti descrivono le qualità individuali di un oggetto, ma non riescono a sintetizzarne l'immagine.
Con la sconfitta del lobulo parietale inferiore, la sensazione dello schema corporeo è disturbata. Una persona non è in grado di capire dove si trova il lato destro e dove il lato sinistro, non riconosce le proprie dita. Un altro tipo di disturbo è l'inconsapevolezza del proprio difetto (il paziente afferma di muovere gli arti paralizzati). Questi pazienti possono sviluppare pseudopolimelia.— sensazione di arti o parti del corpo in più. Tali pazienti possono rimuovere autonomamente l'arto “interferente” o contribuire alla sua amputazione.
Con un danno alla corteccia del giro angolare, il paziente perde il senso della percezione spaziale del mondo che lo circonda, la posizione del proprio corpo e le interconnessioni delle sue parti. Ciò è accompagnato da una varietà di sintomi psicopatologici: depersonalizzazione, derealizzazione. Possono essere osservati a condizione di piena conservazione della coscienza e del pensiero critico.
Con lesioni del lobo parietale dell'emisfero dominante, una persona sviluppa la dislessia - l'incapacità di leggere, distinguere tra destra e sinistra, così come la discalculia - l'incapacità di fare aritmetica. È interessante notare che spesso la discalculia è una malattia indipendente e non una conseguenza di disturbi neurologici o problemi psicologici. Oltre a questi problemi, è possibile anche l'aprassia: una violazione o l'incapacità di eseguire un'azione intenzionale (beh, ad esempio, prendere un bicchiere e bere) pur mantenendo le componenti elementari dell'azione.
Anastasia Sheshukova
Il lobo occipitale occupa le sezioni posteriori degli emisferi. Sulla superficie convessa dell'emisfero, il lobo occipitale non ha confini netti che lo separano dai lobi parietale e temporale, ad eccezione della parte superiore del solco parietale-occipitale, che, situato sulla superficie interna dell'emisfero, separa il lobo parietale dal lobo occipitale. I solchi e le convoluzioni della superficie laterale superiore del lobo occipitale sono instabili e hanno una struttura variabile. Sulla superficie interna del lobo occipitale è presente un solco sperone che separa il cuneo (un lobulo triangolare del lobo occipitale) dalla circonvoluzione linguale e dalla circonvoluzione occipitotemporale.
La funzione del lobo occipitale è associata alla percezione e all'elaborazione delle informazioni visive, all'organizzazione di complessi processi di percezione visiva. In questo caso, la metà superiore della retina è proiettata nella regione del cuneo, che percepisce la luce dai campi visivi inferiori; nella regione del giro lingulare si trova la metà inferiore della retina, che percepisce la luce dai campi visivi superiori.
Isola
L'isolotto, o il cosiddetto lobulo chiuso, si trova in profondità nel solco laterale. L'isolotto è separato dalle sezioni adiacenti adiacenti da una scanalatura circolare. La superficie dell'isolotto è divisa dalla sua scanalatura centrale longitudinale nelle parti anteriore e posteriore. Nell'isoletta viene proiettato un analizzatore del gusto.
corteccia limbica
Sulla superficie interna degli emisferi sopra il corpo calloso si trova il giro del cingolo. Questo giro, con un istmo dietro il corpo calloso, passa nel giro vicino al cavalluccio marino: il giro paraippocampale. Il giro cingolato insieme al giro paragitpocampale costituiscono il giro a volta.
Le superfici interna ed inferiore degli emisferi sono combinate nella cosiddetta corteccia limbica (marginale), insieme all'amigdala del gruppo dei nuclei sottocorticali, al tratto olfattivo e al bulbo, aree dei lobi frontale, temporale e parietale del corteccia cerebrale, nonché con la regione ipotuberosa e la formazione reticolare del tronco. La corteccia limbica è combinata in un unico sistema funzionale: il complesso limbico-reticolare. La funzione principale di queste parti del cervello non è tanto quella di fornire comunicazione con il mondo esterno, ma di regolare il tono della corteccia, le pulsioni e la vita affettiva. Regolano le funzioni complesse e sfaccettate degli organi interni e le risposte comportamentali. Il complesso limbico-reticolare è il più importante sistema integrativo del corpo. Anche il sistema limbico è importante nella formazione delle motivazioni. La motivazione (o motivazione interna) comprende le reazioni istintive ed emotive più complesse (alimentari, difensive, sessuali). Il sistema limbico è coinvolto anche nella regolazione del sonno e della veglia.
La corteccia limbica svolge anche un'importante funzione olfattiva. L'olfatto è la percezione delle sostanze chimiche presenti nell'aria. Cervello olfattivo una persona fornisce il senso dell'olfatto, nonché l'organizzazione di forme complesse di reazioni emotive e comportamentali. Il cervello olfattivo fa parte del sistema limbico.
Il cervello olfattivo è costituito da due sezioni: periferica e centrale. La sezione periferica è rappresentata dal nervo olfattivo, bulbi olfattivi, centri olfattivi primari. La sezione centrale comprende il giro del cavalluccio marino: l'ippocampo, il giro dentato e quello a volta.
L'apparato recettore olfattivo è situato nella mucosa nasale. Attraverso il sistema dei conduttori nervosi, le informazioni dai recettori vengono trasmesse alla sezione corticale dell'analizzatore olfattivo (Fig. 8).
Riso. 8. Analizzatore olfattivo (schema):
1 - epitelio olfattivo, cellule olfattive bipolari; 2 - bulbo olfattivo; 3 - tratto olfattivo; 4 - centri olfattivi primari; 5 - tubercolo visivo; 6 - centro olfattivo corticale; 7 - corpo calloso
La regione corticale dell'analizzatore olfattivo si trova nel giro del cingolo, nel giro del cavalluccio marino e nell'uncino del cavalluccio marino, che insieme formano una regione anulare chiusa. La parte periferica dell'analizzatore olfattivo è collegata alle regioni corticali di entrambi gli emisferi.
Il meccanismo fisiologico della percezione degli odori da parte dell'analizzatore olfattivo non è del tutto chiaro. Esistono due ipotesi principali che spiegano la natura di questo processo da posizioni diverse. Secondo una delle ipotesi, l'interazione tra le molecole odoranti e i chemocettori avviene come una chiave e una serratura, cioè il tipo di molecola corrisponde a un recettore speciale. Un'altra ipotesi si basa sul presupposto che le molecole di una sostanza odorosa possiedano una certa onda di oscillazione, sulla quale i recettori olfattivi sono “sintonizzati”. Le molecole che hanno vibrazioni simili dovrebbero avere un'onda comune e, di conseguenza, dare odori simili.
Il termine “cervello olfattivo” in relazione alla fisiologia umana è alquanto arbitrario e non rivela pienamente la sua funzione multiforme e universale. Il “collocamento” del collegamento centrale del cervello olfattivo negli emisferi cerebrali non è casuale ed è il risultato dell’enorme ruolo “informativo” che l’olfatto ha svolto nel processo di evoluzione nell’adattamento all’ambiente esterno e nella regolazione del complesso reazioni comportamentali. Procurarsi il cibo, scegliere un individuo del sesso opposto, prendersi cura della prole, l'integrità del territorio, organizzare comunità di gruppo all'interno di una specie: tutte queste funzioni quotidiane in molti animali vengono eseguite con la partecipazione diretta di un sistema finemente costruito di ricezione olfattiva e , in base a ciò, la capacità di un numero di animali di inviare specifiche sottili e differenziate sostanze odorose- segnali-informatori.
Le forme universali di reazioni comportamentali negli animali, manifestate nella cura quotidiana dell'habitat, della prole, creano l'impressione che siano dotati di ragione. L'intelligenza apparente è semplicemente il risultato della reazione agli stimoli esterni. Tuttavia, questi stessi stimoli e le risposte ad essi corrispondono perfettamente ai bisogni biologici degli animali.
Nella vita delle persone, l'olfatto ha perso il valore di informazione biologica che aveva negli animali. Il sistema olfattivo umano è progettato sia per svolgere una funzione ristretta, “propria”, sia per una sorta di “carica” delle emozioni. La forza dell'impatto degli odori sulla sfera emotiva, che essi costituiscono il più importante “substrato alimentare delle emozioni”, è ben nota fin dall'antichità nella storia dell'umanità.
Il senso dell'olfatto di una persona può variare. Di norma, queste variazioni sono insignificanti, ma in alcuni casi l'acutezza dell'olfatto può essere molto elevata (degustatori nell'industria dei profumi).
Poiché l'analizzatore olfattivo svolge un ruolo importante nella regolazione delle emozioni, la sua sezione centrale è riferita al sistema limbico, chiamato figurativamente il “denominatore comune” di molte reazioni emotive e viscerosomatiche del corpo.
Grandi emisferi del cervello
sono la parte più grande del cervello. Coprono il cervelletto e il tronco encefalico. Gli emisferi cerebrali costituiscono circa il 78% della massa totale del cervello.
Nel processo di sviluppo ontogenetico dell'organismo, gli emisferi cerebrali si sviluppano dalla vescica cerebrale terminale del tubo neurale, quindi questa parte del cervello è anche chiamata telencefalo.
Gli emisferi cerebrali sono divisi in linea mediana profonda fessura verticale a destra e emisfero sinistro. Nella profondità della parte centrale, entrambi gli emisferi sono interconnessi da una grande adesione: il corpo calloso. Ogni emisfero ha lobi: frontale, parietale, temporale, occipitale.
I lobi degli emisferi cerebrali sono separati tra loro da solchi profondi. Tre solchi profondi sono molto importanti: quello centrale (di Roland), che separa il lobo frontale dal parietale; laterale (silviano), che separa il lobo temporale dal parietale, e parietale-occipitale, che separa il lobo parietale dall'occipitale sulla superficie interna dell'emisfero.
Ogni emisfero ha una superficie superiore-laterale (convessa) - convessa, inferiore - basale e interna - mediale. Ogni lobo dell'emisfero ha circonvoluzioni cerebrali, separate l'una dall'altra da solchi. Dall'alto, l'emisfero è ricoperto da una corteccia, un sottile strato di materia grigia, che consiste in cellule nervose.
Corteccia- la formazione più giovane del sistema nervoso centrale in termini evolutivi. Nell'uomo raggiunge il suo massimo sviluppo. La corteccia cerebrale è di grande importanza nella regolazione dell'attività vitale del corpo, nell'attuazione di forme complesse di comportamento e nella formazione delle funzioni neuropsichiche.
Sotto la corteccia c'è materia bianca emisferi, consiste di processi di cellule nervose - conduttori. A causa della formazione delle circonvoluzioni cerebrali, la superficie totale della corteccia cerebrale aumenta in modo significativo. L'area totale della corteccia emisferica è di 1200 cm2, di cui 2/3 della sua superficie si trovano nella profondità dei solchi e 1/3 sulla superficie visibile degli emisferi. Ogni lobo del cervello ha un significato funzionale diverso.
Corteccia è costituito da 4 lobi, separati tra loro da solchi. I solchi principali che separano i lobi frontale, parietale e temporale sono Rolandov e Sylviev.
Lobi della corteccia cerebrale :
frontale (frontale ),
parietale (parietale ),
occipitale (occipitale ),
temporale (temporale ),
In relazione alle azioni si parla di sistemi locali del cervello.
SISTEMI LOCALI DEL CERVELLO:
Regioni occipitali del cervello svolgere le funzioni di organizzazione della percezione visiva. Zone primarie della corteccia occipitale - funzioni elementari visione.
Sezioni secondarie della corteccia occipitale - funzioni optognostiche.
Regioni temporali del cervello"responsabile" dell'organizzazione della percezione uditiva. Le zone primarie della corteccia temporale sono le funzioni elementari dell'udito.
Zone secondarie della corteccia temporale - funzioni acustico-gnostiche.
Parti sensomotorie e premotorie del cervello- organizzazione dei movimenti. 1. Parti postcentrali del cervello - organizzazione afferente dei movimenti. 2. Zone della corteccia premotoria: organizzazione efferente (programmazione) del movimento.
3. Zone motorie della corteccia: un analizzatore motorio, l'implementazione di un programma motorio. Lobi frontali del cervello (regioni prefrontali) effettuare la regolazione dell'attività mentale, cioè la regolazione degli stati di attività, dei movimenti e delle azioni volontarie, dei processi cognitivi e della sfera emotiva e personale, sono generalmente responsabili del pensiero, dell'attività intellettuale. Regioni parietali del cervello sono rappresentati da zone primarie (tutti i tipi di sensibilità cutanea-cinestetica), zone secondarie (rappresentazioni visuo-spaziali, idee sullo schema corporeo, somatognosi, stereognosi) e zone terziarie.
Lobo frontale occupa le porzioni anteriori degli emisferi. È separato dal lobo parietale dal solco centrale e dal lobo temporale dal solco laterale. Ci sono quattro giri nel lobo frontale: uno verticale - precentrale e tre orizzontali - giro frontale superiore, medio e inferiore.
Le circonvoluzioni sono separate l'una dall'altra da solchi. Sulla superficie inferiore dei lobi frontali si distinguono il giro diretto e quello orbitale. Il giro diretto si trova tra il bordo interno dell'emisfero, il solco olfattivo e il bordo esterno dell'emisfero.
Nelle profondità del solco olfattivo si trovano il bulbo olfattivo e il tratto olfattivo. Il lobo frontale di una persona costituisce il 25-28% della corteccia, la massa media del lobo frontale è di 450 g.
La funzione dei lobi frontali è associata all'organizzazione dei movimenti volontari, ai meccanismi motori della parola, alla regolazione di forme complesse di comportamento e ai processi di pensiero. Diversi centri funzionalmente importanti sono concentrati nelle circonvoluzioni del lobo frontale. Il giro centrale anteriore è una "rappresentazione" della zona motoria primaria con una proiezione rigorosamente definita delle parti del corpo. Il viso è "situato" nel terzo inferiore del giro, la mano nel terzo medio, la gamba terzo superiore. Il tronco è rappresentato nelle sezioni posteriori del giro frontale superiore. Pertanto, una persona viene proiettata sottosopra e in basso nel giro centrale anteriore.
Giro centrale anteriore insieme alle sezioni posteriori adiacenti delle circonvoluzioni frontali, svolge un ruolo funzionalmente molto importante. È il centro dei movimenti volontari. Nelle profondità della corteccia del giro centrale dal cosiddetto cellule piramidali -motoneurone centrale- inizia la via motoria principale, la via piramidale o corticospinale. I processi periferici dei motoneuroni emergono dalla corteccia, si riuniscono in un unico potente fascio, attraversano la sostanza bianca centrale degli emisferi ed entrano nel tronco cerebrale attraverso la capsula interna; all'estremità del tronco encefalico si incrociano parzialmente (passando da un lato all'altro) e poi scendono nell'encefalo midollo spinale. Questi rami terminano con materia grigia midollo spinale. Lì entrano in contatto con il motoneurone periferico e gli trasmettono impulsi dal motoneurone centrale. Gli impulsi di movimento volontario vengono trasmessi lungo il percorso piramidale.
Nelle sezioni posteriori del giro frontale superiore è presente anche un centro extrapiramidale della corteccia, che è strettamente connesso anatomicamente e funzionalmente con le formazioni del cosiddetto sistema extramiramidale. Sistema extrapiramidale- un sistema motorio che aiuta l'attuazione del movimento arbitrario. Questo è un sistema per "fornire" movimenti arbitrari. Essendo filogeneticamente più antico del sistema piramidale, il sistema extrapiramidale umano fornisce la regolazione automatica degli atti motori “appresi”, il mantenimento del tono muscolare generale, la “prontezza” dell'apparato motorio periferico al movimento, la ridistribuzione del tono muscolare durante i movimenti. Inoltre, è coinvolto nel mantenimento di una postura normale.
Nella parte posteriore del giro frontale medio si trova il centro oculomotore frontale, che controlla la rotazione amichevole e simultanea della testa e degli occhi (il centro di rotazione della testa e degli occhi nella direzione opposta). L'irritazione di questo centro fa girare la testa e gli occhi nella direzione opposta. La funzione di questo centro è di grande importanza nell'attuazione dei cosiddetti riflessi di orientamento, molto importanti per la preservazione della vita degli animali.
Nella parte posteriore del giro frontale inferiore si trova centro motorio della parola(Centro di Brock).
Anche la corteccia frontale degli emisferi cerebrali riceve Partecipazione attiva nella formazione del pensiero, nell'organizzazione di attività mirate, nella pianificazione a lungo termine.
Lobo temporale occupa la superficie laterale inferiore degli emisferi. Il lobo temporale è separato dai lobi frontale e parietale da un solco laterale.
Sulla superficie laterale superiore del lobo temporale ci sono tre circonvoluzioni: superiore, media E inferiore. Il giro temporale superiore è situato tra il solco silviano e quello temporale superiore, il giro medio è tra i solchi temporali superiore e inferiore, e il giro inferiore è tra il solco temporale inferiore e la fessura cerebrale trasversa. Sulla superficie inferiore del lobo temporale si distinguono il giro temporale inferiore, il giro occipitotemporale laterale e il giro dell'ippocampo (zampe del cavalluccio marino).
Funzione del lobo temporale associato alla percezione delle sensazioni uditive, gustative, olfattive, all'analisi e alla sintesi dei suoni del linguaggio, ai meccanismi di memoria. Il principale centro funzionale della superficie laterale superiore del lobo temporale è situato nel giro temporale superiore. Qui è il centro uditivo, o gnostico, del discorso (centro di Wernicke).
Nel giro temporale superiore e sulla superficie interna del lobo temporale si trova l'area di proiezione uditiva della corteccia. L'area di proiezione olfattiva si trova nel giro dell'ippocampo, soprattutto nella sua sezione anteriore (il cosiddetto uncino). Accanto alle zone di proiezione olfattiva si trovano anche quelle gustative.
I lobi temporali svolgono un ruolo importante nell'organizzazione del complesso processo mentale, in particolare la memoria.
Lobo parietale occupa le superfici laterali superiori dell'emisfero. Dal lobo parietale frontale, anteriormente e lateralmente, è limitato dal solco centrale, dal temporale dal basso - dal solco laterale, dall'occipitale - da una linea immaginaria che passa dal bordo superiore del solco parietale-occipitale al bordo inferiore dell'emisfero.
Sulla superficie laterale superiore del lobo parietale ci sono tre convoluzioni: una verticale - centrale posteriore e due orizzontali - parietale superiore e parietale inferiore. La parte del giro parietale inferiore, che avvolge la parte posteriore del solco laterale, è chiamata sopramarginale (sopramarginale), e la parte che circonda il solco superiore giro temporale, un dominio nodale (angolare).
Il lobo parietale, come il lobo frontale, ne costituisce una parte significativa emisferi cerebrali. In termini filogenetici, si distingue una vecchia sezione - il giro centrale posteriore, una nuova - il giro parietale superiore e una più recente - il giro parietale inferiore. La funzione del lobo parietale è associata alla percezione e all'analisi degli stimoli sensibili, all'orientamento spaziale. Diversi centri funzionali sono concentrati nelle circonvoluzioni del lobo parietale.
Nel giro centrale posteriore, i centri di sensibilità vengono proiettati con una proiezione corporea simile a quella del giro centrale anteriore. Nel terzo inferiore del giro, viene proiettato il viso, nel terzo medio - il braccio, il busto, nel terzo superiore - la gamba. Nel giro parietale superiore ci sono centri responsabili di tipi complessi di sensibilità profonda: sensazione muscolo-articolare, bidimensionale-spaziale, senso di peso e volume di movimento, senso di riconoscere gli oggetti al tatto.
Pertanto, la sezione corticale dell'analizzatore sensibile è localizzata nel lobo parietale.
I centri della prassi si trovano nel lobo parietale inferiore. La prassi è intesa come movimenti mirati che sono stati automatizzati nel processo di ripetizioni ed esercizi, che si sviluppano nel processo di apprendimento e pratica costante durante la vita individuale.
Camminare, mangiare, vestirsi, scrivere meccanicamente, diversi tipi l'attività lavorativa (ad esempio, i movimenti del conducente per guidare un'auto, falciare, ecc.) sono prassi.
Prassi- la più alta manifestazione della funzione motoria umana. Viene effettuato come risultato delle attività combinate di vari territori. corteccia cerebrale.
Lobo occipitale occupa le regioni posteriori degli emisferi. Sulla superficie convessa dell'emisfero, il lobo occipitale non ha confini netti che lo separano dai lobi parietale e temporale, ad eccezione della parte superiore del solco parietale-occipitale, che, situato sulla superficie interna dell'emisfero, separa il lobo parietale dal lobo occipitale.
I solchi e le convoluzioni della superficie laterale superiore del lobo occipitale sono instabili e hanno una struttura variabile.
Sulla superficie interna del lobo occipitale è presente un solco sperone che separa il cuneo (un lobulo triangolare del lobo occipitale) dalla circonvoluzione linguale e dalla circonvoluzione occipitotemporale.
La funzione del lobo occipitale è associata alla percezione e all'elaborazione informazioni visive, organizzazione di processi complessi della percezione visiva. In questo caso, la metà superiore della retina è proiettata nella regione del cuneo, che percepisce la luce dai campi visivi inferiori; nella regione del giro lingulare si trova la metà inferiore della retina, che percepisce la luce dai campi visivi superiori.
Isola, o il cosiddetto lobulo chiuso, si trova nella profondità del solco laterale. L'isolotto è separato dalle sezioni adiacenti adiacenti da una scanalatura circolare.
La superficie dell'isolotto è divisa dalla sua scanalatura centrale longitudinale nelle parti anteriore e posteriore. Nell'isola è proiettato analizzatore del gusto.
Analizzatore olfattivo
Cellule nervose, percependo irritazioni olfattive, si trovano nella mucosa delle parti superiori della cavità nasale. Da qui gli assoni di queste cellule entrano nella cavità cranica ed entrano nei bulbi olfattivi. Da loro vengono inviate le fibre nervose lobo temporale (superficie interna) dove sono le cellule nervose? analizzatore olfattivo.
Analizzatore del gusto
Questo analizzatore inizia nelle terminazioni nervose delle papille gustative della lingua, che in esse sono rappresentate dalle papille gustative. Le fibre nervose che escono dalle papille gustative vanno al cervello e terminano, come i nervi dell'analizzatore olfattivo, a superficie interna Lobo temporale.
corpo calloso- lamina sottile arcuata, filogeneticamente giovane, raccorda le superfici mediane entrambi gli emisferi. La parte centrale allungata del corpo calloso passa dietro in un ispessimento, e davanti si curva e si curva in modo arcuato.
Il corpo calloso collega le parti filogeneticamente più giovani degli emisferi e svolge un ruolo importante nello scambio di informazioni tra di loro.
STELO CERVELLO O STELO CERVELLO -
un sistema tradizionalmente distinto di regioni cerebrali, che è una formazione estesa che continua il midollo spinale.
Il tronco cerebrale comprende sempre il midollo allungato, il ponte e il mesencefalo. Spesso comprende il cervelletto, talvolta il diencefalo.
Midollo -
dipartimento del cervello. Esiste anche il nome tradizionale bulbus (bulbo, per la forma di questo reparto).
Il midollo allungato entra nel tronco cerebrale.
All'esterno, sul lato ventrale (facciale), ci sono le piramidi (contengono il tratto corticospinale - il percorso dalla corteccia ai motoneuroni del midollo spinale) e le olive (contengono i nuclei dell'oliva inferiore associati al mantenimento dell'equilibrio). . Sul lato dorsale: fasci sottili e a forma di cuneo che terminano con tubercoli di nuclei sottili e a forma di cuneo (cambiano le informazioni sulla sensibilità profonda della metà inferiore e superiore del corpo, rispettivamente), la metà inferiore della fossa romboidale, che è il fondo del quarto ventricolo, e i corpi di corda che lo separano, o le gambe inferiori del cervelletto.
All'interno si trovano anche i nuclei dall'VIII al XII (e uno dei nuclei VII) dei nervi cranici, parte della formazione reticolare, l'ansa mediale e altre vie ascendenti e discendenti.
Ha la forma di un tronco di cono.
Grazie alla ricerca di scienziati come R. Magnus e I. F. Klein, è stato stabilito che esiste un complesso sistema di centri riflessi nel midollo allungato che forniscono una certa posizione nel corpo a causa di riflessi statici e statico-cinetici. Questi riflessi, infatti, sono meccanismi di ridistribuzione del tono muscolare in modo tale che venga mantenuta una postura comoda per l'animale (riflessi tonico-posturali) o il ritorno ad una determinata da una postura scomoda (riflessi rettificativi), e l'equilibrio viene mantenuto anche durante l'accelerazione (riflessi stato-cinetici). L'implementazione di questi riflessi avviene con la partecipazione di formazioni del tronco come la formazione reticolare, il nucleo rosso e i nuclei vestibolari.
Formazione reticolare - è una formazione che va dal midollo spinale al talamo in direzione rostrale (corteccia). Oltre a partecipare all'elaborazione delle informazioni sensoriali, la formazione reticolare ha un effetto attivante sulla corteccia cerebrale, controllando così l'attività del midollo spinale. Per la prima volta viene scoperto il meccanismo d'azione della formazione reticolare tono muscolareè stato stabilito da R. Granit: ha dimostrato che la formazione reticolare è in grado di modificare l'attività dei motoneuroni γ, in conseguenza dei quali gli ihaxons (γ-efferenti) causano la contrazione dei fusi muscolari e, di conseguenza, un aumento negli impulsi afferenti provenienti dai recettori muscolari. Questi impulsi, entrando nel midollo spinale, causano l'eccitazione dei motoneuroni α, che è la causa del tono muscolare.
È stato stabilito che due gruppi di neuroni partecipano allo svolgimento di questa funzione della formazione reticolare: neuroni della formazione reticolare del ponte e neuroni della formazione reticolare del midollo allungato. Comportamento dei neuroni della formazione reticolare midollo allungato simile al comportamento dei neuroni nella formazione reticolare del ponte: provocano l'attivazione dei motoneuroni α dei muscoli flessori e, quindi, inibiscono l'attività dei motoneuroni α dei muscoli estensori. I neuroni della formazione reticolare del ponte agiscono esattamente al contrario, eccitano i motoneuroni α dei muscoli estensori e inibiscono l'attività dei motoneuroni α dei muscoli flessori. La formazione reticolare ha una connessione con il cervelletto (parte delle informazioni da esso va ai neuroni del midollo allungato (dai nuclei del cervelletto sugheroso e sferico), e dalla tenda ai neuroni del ponte) e con il corteccia cerebrale, da cui riceve informazioni. Ciò suggerisce che la formazione reticolare sia un collettore di flusso sensoriale aspecifico, possibilmente coinvolto nella regolazione dell'attività muscolare.
importante significato funzionale reticolare, o una formazione di rete del tronco cerebrale, che si sviluppa in connessione con l'emergere di un sistema di vago, vestibolare e nervi trigeminali.
La formazione reticolare è costituita da cellule nervose di varie dimensioni e forme, nonché da una fitta rete fibre nervose, andando in direzioni diverse e localizzati principalmente vicino al sistema ventricolare. Nei rapporti cortico-sottocorticali viene data primaria importanza alla formazione reticolare. Si trova ai piani intermedi. midollo allungato,ipotalamo, materia grigia del tegmento, ponte.
Numerosi collaterali provenienti da tutti i sistemi afferenti (sensoriali) si avvicinano alla formazione reticolare. Attraverso questi collaterali, eventuali irritazioni provenienti dalla periferia, dirigendosi verso alcune aree della corteccia lungo specifiche vie del sistema nervoso, raggiungono anche la formazione reticolare. I sistemi ascendenti non specifici (cioè i percorsi dalla formazione reticolare) forniscono l'eccitazione della corteccia cerebrale, l'attivazione della sua attività.
Insieme ai sistemi ascendenti non specifici, attraverso il tronco encefalico passano i sistemi discendenti non specifici, che influenzano i meccanismi riflessi spinali.
La formazione reticolare è strettamente connessa al sistema limbico e alla corteccia cerebrale. A causa di ciò, si forma una connessione funzionale tra le parti superiori del sistema nervoso centrale e il tronco. cervello. Questo sistema è chiamato complesso limbico-reticolare o asse limbico-reticolare. Questo complesso complesso strutturale e funzionale garantisce l'integrazione delle funzioni più importanti, alla cui attuazione partecipano varie parti del cervello.
È noto che lo stato di veglia della corteccia è fornito da sistemi specifici e non specifici. La reazione di attivazione è mantenuta da una fornitura costante di impulsi provenienti dai recettori uditivo, visivo, olfattivo, gustativo e analizzatori cutanei cinestetici. Questi stimoli vengono trasmessi lungo specifiche vie afferenti a varie parti della corteccia. Da tutti i partecipanti talamo, e poi numerosi collaterali alla formazione reticolare partono nella corteccia degli emisferi cerebrali delle vie afferenti, che ne assicurano l'attività attivante ascendente.
A sua volta, la formazione reticolare riceve impulsi da cervelletto, nuclei sottocorticali, sistema limbico che forniscono risposte comportamentali emotivamente adattive, forme di comportamento motivazionali. Tuttavia, il grado di fornitura di reazioni riflesse incondizionate adattive da parte di un sistema non specifico nell'uomo e negli animali è diverso. Se negli animali le formazioni sottocorticali e il sistema limbico svolgono un ruolo di primo piano nel soddisfare i bisogni vitali dell'organismo per la sua sopravvivenza in ambiente, quindi nell'uomo, a causa del predominio della corteccia, l'attività delle strutture profonde del cervello (formazioni sottocorticali, sistema limbico, formazione reticolare), in misura maggiore che in un animale, è subordinata a corteccia cerebrale. La formazione reticolare gioca un ruolo importante nella regolazione del tono muscolare. Il tono muscolare è regolato da due tipi di tratti reticolospinali. Il tratto reticolospinale a conduzione veloce regola i movimenti veloci; Via reticolospinale a conduzione lenta - movimenti tonici lenti.
La formazione reticolare del midollo allungato è coinvolta nella comparsa della rigidità decerebrata. Quando il tronco cerebrale viene tagliato sopra il midollo allungato, l'attività dei neuroni che hanno un effetto inibitorio sui motoneuroni del midollo spinale diminuisce, il che porta ad un forte aumento del tono dei muscoli scheletrici.
Funzioni del midollo allungato
Riflessi protettivi (ad esempio tosse, starnuti).
Riflessi vitali (ad esempio la respirazione).
regolazione del tono vascolare.
Centri riflessi del midollo allungato:
protettivi (tosse, starnuti, ecc.)
centri di controllo del tono muscolo scheletrico per mantenere la postura di una persona.
accorciamento o allungamento del tempo del riflesso spinale
digestione
attività cardiaca
Ponte
Il ponte Varolii (a nome di Costanzo Varolia), o ponte - una parte del cervello, è, insieme al cervelletto, parte del rombencefalo. appartiene al cervello,
Alla base del ponte sono presenti vie discendenti: tratto piramidale corticospinale, tratti corticobulbari, tratti corticoponte.
mesencefalo(lat. Mesencefalo) - parte del cervello, l'antico centro visivo. Incluso nel tronco cerebrale.
funzioni del mesencefalo
1. motore,
2. sensoriale (vista, udito),
3. Regolamentare gli atti di masticazione e deglutizione,

