Emisferi grandi grandi emisferi
cervello, formazioni accoppiate unite dal corpo calloso nel cosiddetto telencefalo. Superficie emisferi cerebrali rappresentato da numerose circonvoluzioni profonde grandi o piccole. Ci sono lobi: frontale, parietale, temporale, insulare, occipitale. Materia grigia del cervello, composta da cellule nervose- neuroni, forma la corteccia cerebrale e i gangli sottocorticali (nodi). materia bianca formato dai processi dei neuroni che compongono le vie cerebrali.
GRANDI EMISFERIGRANDI EMISFERI del cervello, formazioni appaiate unite dal corpo calloso (cm. Corpo calloso) nel cosiddetto telencefalo. La superficie degli emisferi cerebrali è rappresentata da numerose circonvoluzioni profonde grandi o piccole. Ci sono lobi: frontale, parietale, temporale, insulare, occipitale. La materia grigia del cervello, costituita da cellule nervose - neuroni, forma la corteccia cerebrale e i gangli sottocorticali (cm. GANGLIO)(nodi). La materia bianca è formata dai processi dei neuroni che compongono i percorsi del cervello.
Dizionario enciclopedico. 2009 .
Scopri cosa sono i "grandi emisferi" in altri dizionari:
Il cervello ha formazioni accoppiate, unite dal corpo calloso nel cosiddetto. telencefalo. La superficie degli emisferi cerebrali è rappresentata da numerose circonvoluzioni profonde grandi o piccole. Ci sono lobi: frontale, parietale, temporale, insulare... Grande dizionario enciclopedico
Il cervello, formazioni accoppiate unite dal corpo calloso nel cosiddetto. telencefalo. La superficie del B.p. è rappresentata da numerosi. B. o M. circonvoluzioni profonde. Ci sono lobi: frontale, parietale, temporale, insulare, occipitale. Grigio in... ... Scienze naturali. Dizionario enciclopedico
GRANDI EMISFERI DEL CERVELLO- parti superiori del cervello, costituite dallo strato superficiale della corteccia cerebrale e dalle parti profonde della sottocorteccia; ricoprono il cervelletto e il tronco encefalico. B. p. g. m. diviso per linea mediana agli emisferi destro e sinistro, che sono profondi... Psicomotoria: dizionario-libro di consultazione
L'Oceano Artico, a differenza dell'Oceano Australe, è completamente di natura mediterranea. Ha confini naturali per una distanza considerevole e solo in tre punti si fonde direttamente con le acque dell'Atlantico e del Pacifico... ...
L'Oceano Artico, a differenza di quello meridionale, è completamente di natura mediterranea. Ha confini naturali per una distanza considerevole e solo in tre punti si fonde direttamente con le acque dell'Atlantico e del Pacifico... ... Dizionario Enciclopedico F.A. Brockhaus e I.A. Efron
Chiamata anche morfologia comparata, è lo studio dei modelli di struttura e sviluppo degli organi mediante confronto vari tipi Creature viventi. I dati dell'anatomia comparata sono la base tradizionale della classificazione biologica. Sotto morfologia... Enciclopedia di Collier
Vista in sezione del cervello maschile adulto. Il cervello umano (lat. encefalo) riguarda ... Wikipedia
La scienza che studia la struttura del corpo singoli organi, i tessuti e le loro relazioni nel corpo. Tutti gli esseri viventi sono caratterizzati da quattro caratteristiche: crescita, metabolismo, irritabilità e capacità di riprodursi. La combinazione di queste caratteristiche... ... Enciclopedia di Collier
Animali (Mammalia), la classe dei vertebrati, la più famoso gruppo animali, tra cui più di 4.600 specie della fauna mondiale. Comprende gatti, cani, mucche, elefanti, topi, balene, persone, ecc. Nel corso dell'evoluzione, i mammiferi hanno realizzato le più ampie... ... Enciclopedia di Collier
I Medicina Sistema di medicina conoscenza scientifica e attività pratiche, i cui obiettivi sono rafforzare e preservare la salute, prolungare la vita delle persone, prevenire e curare le malattie umane. Per portare a termine questi compiti, M. studia la struttura e... ... Enciclopedia medica
Il cervello umano è un sistema estremamente complesso. Grazie a questo organo, le persone hanno raggiunto il livello di sviluppo osservato ora. Come è lui?
Sviluppo evolutivo
Il corso di biologia scolastica moderna copre argomenti da semplici a complessi. All'inizio stiamo parlando su cellule, protozoi, batteri, piante, funghi. Successivamente c'è una transizione verso gli animali e l'uomo. In una certa misura, ciò riflette l’ipotetico corso dell’evoluzione. Osservando la struttura, ad esempio, dei vermi, è facile notare che è molto più semplice di quella degli esseri umani o degli animali superiori. Ma questi organismi hanno qualcosa di importante: un ganglio nervoso che svolge le funzioni del cervello.
Prosencefalo
Se chiedi a qualcuno di disegnare il contenuto di un teschio umano, molto probabilmente disegnerà un diagramma degli emisferi. Questa è davvero una delle parti più evidenti e più grandi. Ma il prosencefalo contiene anche il midollo allungato. In generale, la loro struttura è piuttosto complessa. E se prendiamo in considerazione una divisione più dettagliata, possiamo anche nominare tutti i dipartimenti prosencefalo:
- ippocampo;
- gangli della base;
- grande cervello.
Naturalmente esiste una divisione ancora più dettagliata, ma, di regola, interessa solo gli specialisti. Bene, per coloro che stanno semplicemente espandendo i propri orizzonti, sarà molto più interessante scoprire cosa stanno facendo tutti questi dipartimenti. Allora quali sono le funzioni del prosencefalo? E perché ci sono differenze tra il modo di pensare dei destrimani e quello dei mancini?
Funzioni
Il proencefalo comprende le parti sviluppate più recentemente. E questo significa che è grazie a loro che una persona ha le qualità che possiede. E se il diencefalo si occupa principalmente della regolazione del metabolismo, dei riflessi e dei bisogni primitivi, oltre che semplici attività fisica, quindi gli emisferi sono il luogo stesso in cui sorgono i pensieri coscienti, dove avviene l'apprendimento e la memorizzazione delle informazioni e dove viene creato qualcosa di nuovo.
Gli emisferi sono anche convenzionalmente divisi in diverse zone-parti: parietale, frontale, posteriore e temporale. E qui ci sono cellule coinvolte, tra le altre cose, nell'analisi delle informazioni provenienti dall'esterno: centri visivi, uditivi, olfattivi, gustativi e tattili.
La cosa più interessante è che da un punto di vista funzionale gli emisferi sinistro e destro sono diversi. Naturalmente, ci sono casi in cui, quando una parte del cervello è stata danneggiata, un'altra ha assunto i suoi compiti, cioè c'è una certa intercambiabilità, ma in un caso normale la situazione può essere così: emisfero sinistro si occupa dell'analisi dell'intonazione del discorso di un'altra persona, e quello di destra si occupa dell'interpretazione del significato di ciò che è stato detto. Questo è il motivo per cui i mancini e i destrimani, che hanno parti diverse più sviluppate, la pensano in modo leggermente diverso.
Inoltre, le funzioni del cervello anteriore includono la memoria, varie reazioni agli stimoli esterni, la pianificazione e la costruzione di scenari e situazioni future. Qui si trova anche il centro vocale. Qui si svolge tutta l'attività nervosa superiore: creatività, riflessione, idee.
È anche piuttosto interessante che il prosencefalo si sviluppi attivamente non solo nel periodo prenatale, ma anche nei primi anni di vita. Ogni nuova abilità e abilità, parola appresa, qualsiasi Informazioni importanti- tutto ciò crea nuovo connessioni neurali. E questo tipo di mappa è unica per ogni persona.

- Le capacità di pensiero non dipendono dalla massa del cervello, ma sono correlate a un valore come il numero di circonvoluzioni.
- La velocità dei segnali tra i neuroni raggiunge i 288 chilometri orari. Man mano che si invecchia, questa cifra diminuisce.
- Il cervello consuma la maggior quantità di energia tra organi umani- circa 20%. Questo è un indicatore enorme, considerando che la sua massa rispetto al corpo è solo del 2%. Inoltre, per il suo normale funzionamento, è necessaria una quantità sufficiente di liquido nel corpo.
- L’affermazione secondo cui il cervello utilizza solo il 10% delle sue risorse è un mito. In effetti, non possono funzionare molti centri contemporaneamente, ma in un modo o nell'altro sono tutti coinvolti.
campi_testo
campi_testo
freccia_verso l'alto
Il diencefalo, insieme al tronco cerebrale, è coperto dall'alto e dai lati grandi emisferi - telencefalo. Gli emisferi sono costituiti da gangli sottocorticali (gangli della base) e hanno cavità -. L'esterno degli emisferi è coperto (con un mantello).
Gangli basali o gangli sottocorticali
campi_testo
campi_testo
freccia_verso l'alto
Gangli della base O nodi sottocorticali (nuclei basali)– le formazioni sono filogeneticamente più antiche della corteccia. I gangli della base prendono il nome dal fatto che si trovano alla base degli emisferi cerebrali, nella loro parte basale. Questi includono i nuclei caudato e lenticolare, uniti nello striato, nel recinto e nell'amigdala.
Nucleo caudato
campi_testo
campi_testo
freccia_verso l'alto
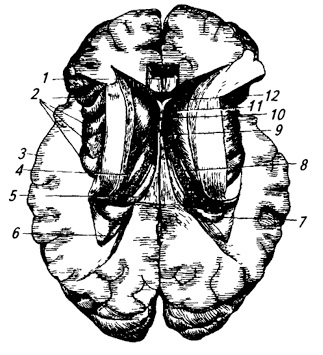
Nucleo caudato (nucleo caudato) allungato nel piano sagittale e fortemente curvo (Fig. 3.22; 3.32; 3.33). La sua parte anteriore e ispessita lo è Testa– posto davanti al talamo ottico, nella parete laterale del corno anteriore ventricolo laterale, dietro si restringe gradualmente e si trasforma in coda. Il nucleo caudato ricopre il talamo visivo davanti, sopra e lateralmente.
Riso. 3.22.
1 – nucleo caudato;
2 – colonne della volta;
3 – ghiandola pineale;
4 – superiore e
5 – collicolo inferiore;
6 – fibre del peduncolo cerebellare medio;
7 – via del peduncolo cerebellare superiore (preparata);
8 – nucleo della tenda;
9 – verme;
10 – sferico,
11 – sughero e
13 – nucleo dentato;
12 – corteccia cerebellare;
14 – peduncolo cerebellare superiore;
15 – triangolo del guinzaglio;
16 – cuscino talamico;
17 – talamo visivo;
18 – commissura posteriore;
19 – terzo ventricolo;
20 – nucleo anteriore del talamo visivo
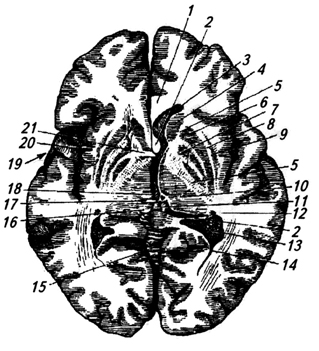
 Riso. 3.32.
Riso. 3.32. Riso. 3.32. Cervello - sezione orizzontale dei ventricoli laterali:
1 – corpo calloso;
2 – isola;
3 – corteccia;
4 – coda del nucleo caudato;
5 – volta;
6 – corno posteriore del ventricolo laterale;
7 – ippocampo;
8 – plesso coroideo;
9 – foro interventricolare;
10 – partizione trasparente;
11 – testa del nucleo caudato;
12 – corno anteriore ventricolo laterale
Nucleo a forma di lenticchia
campi_testo
campi_testo
freccia_verso l'alto
Nucleo a forma di lenticchia (nucleo lentiforme) situato all'esterno del talamo ottico, a livello dell'insula. La forma del nucleo è simile a una piramide triangolare, con la base rivolta verso l'esterno. Il nucleo è nettamente diviso da strati di sostanza bianca in una parte laterale di colore più scuro - conchiglia e mediale - palla pallida, costituito da due segmenti: interno ed esterno (Fig. 3.33; 3.34).
 Riso. 3.33.
Riso. 3.33. Riso. 3.33. Sezione orizzontale degli emisferi cerebrali a livello dei gangli della base:
1 - corpo calloso;
2 – volta;
3 – corno anteriore del ventricolo laterale;
4 – testa del nucleo caudato;
5 – capsula interna;
6 – conchiglia;
7 – globo pallido;
8 – capsula esterna;
9 – recinzione;
10 – talamo;
11 – ghiandola pineale;
12 – coda del nucleo caudato;
13 – plesso corioideo del ventricolo laterale;
14 – corno posteriore del ventricolo laterale;
15 – verme cerebellare;
16 – quadrigemino;
17 – commissura posteriore;
18 – cavità del terzo ventricolo;
19 – fossa del solco laterale;
20 – isola;
21 – commissura anteriore
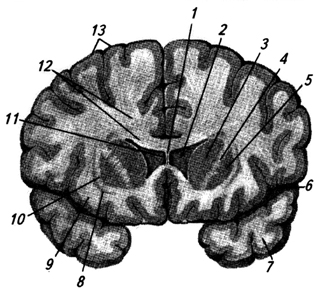 Riso. 3.34.
Riso. 3.34. Riso. 3.34. Sezione frontale attraverso gli emisferi cerebrali a livello dei gangli della base:
1 - corpo calloso;
2 – ventricolo laterale;
3 – nucleo caudato (testa);
4 – capsula interna;
5 - nucleo a forma lenticolare;
6 – scanalatura laterale;
7 - lobo temporale;
8 – recinzione;
9 – isola;
10 – capsula esterna;
11 – partizione trasparente;
12 – splendore del corpo calloso;
13 – corteccia cerebrale
Conchiglia
campi_testo
campi_testo
freccia_verso l'alto
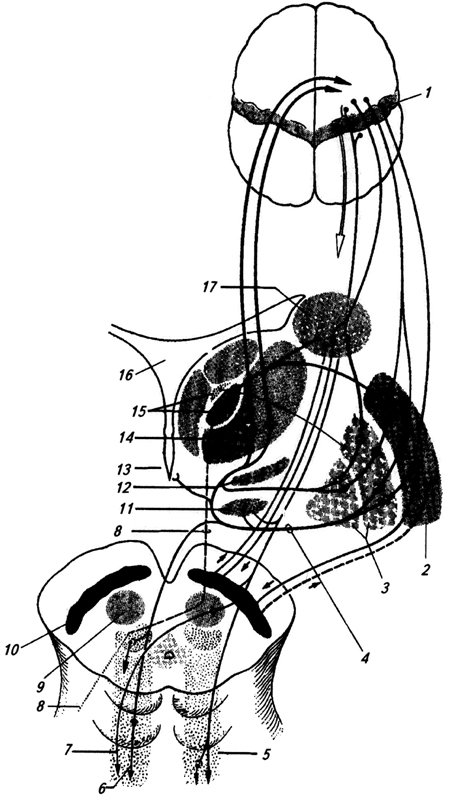 Riso. 3.35.
Riso. 3.35. Conchiglia (putamen) per caratteristiche genetiche, strutturali e funzionali è prossimo al nucleo caudato.
Entrambe queste formazioni hanno una struttura più complessa del globo pallido. Le fibre si avvicinano a loro principalmente dalla corteccia cerebrale e dal talamo (Fig. 3.35).
Riso. 3.35. Connessioni afferenti ed efferenti dei gangli della base:
1 - giro precentrale;
2 – conchiglia;
3 – segmenti esterni ed interni del globo pallido;
4 – ansa lenticolare;
5 - formazione reticolare;
6 – tratto reticolospinale,
7 - tratto rubrospinale;
8 – tratto cerebellotalamico (dal nucleo dentato del cervelletto);
9 – nucleo rosso;
10 – sostanza nera;
11 – nucleo subtalamico;
12 – Zona incerta;
13 – ipotalamo;
14 – ventrolaterale,
15 – nuclei intralaminari e centromediani del talamo;
16 – III ventricolo;
17 – nucleo caudato
Palla pallida
campi_testo
campi_testo
freccia_verso l'alto
Il globo pallido (globus pallidus) è principalmente associato alla conduzione di impulsi lungo numerosi percorsi discendenti nelle strutture sottostanti del cervello: il nucleo rosso, la substantia nigra, ecc. Le fibre dei neuroni del globo pallido vanno agli stessi nuclei di il talamo che sono collegati al cervelletto. Da questi nuclei partono numerosi percorsi che portano alla corteccia cerebrale.
Il globo pallido riceve impulsi dal nucleo caudato e dal putamen.
Striato ( corpo striato) (striato), che unisce i nuclei caudato e lenticolare, appartiene all'efferente sistema extrapiramidale. I dendriti dei neuroni striatali sono ricoperti da numerose spine. Su di essi terminano le fibre dei neuroni della corteccia, del talamo e della substantia nigra (Fig. 3.35). A loro volta, i neuroni striatali inviano assoni ai nuclei intralaminare, anteriore e laterale del talamo. Da loro, le fibre vanno alla corteccia e quindi il circuito di feedback tra i neuroni corticali e lo striato è chiuso.
Durante il processo di filogenesi, questi nuclei furono costruiti sopra i nuclei del mesencefalo. Ricevendo impulsi dal talamo, lo striato prende parte a movimenti automatici complessi come camminare, arrampicarsi e correre. Nei nuclei dello striato gli archi degli incondizionati più complessi sono chiusi, cioè riflessi congeniti. Il sistema extrapiramidale è filogeneticamente più antico del sistema piramidale. Nel neonato, quest'ultimo non è ancora sufficientemente sviluppato e gli impulsi ai muscoli vengono trasmessi dai gangli sottocorticali attraverso il sistema extrapiramidale. Di conseguenza, i movimenti del bambino nei primi mesi di vita sono caratterizzati da generalizzazione e indifferenziazione. Man mano che la corteccia cerebrale si sviluppa, gli assoni delle sue cellule crescono fino ai gangli della base e l'attività di questi ultimi inizia ad essere regolata dalla corteccia. I gangli sottocorticali sono associati non solo alle reazioni motorie, ma anche a funzioni vegetative- questi sono i più alti centri sottocorticali sistema nervoso autonomo.
Amigdala
campi_testo
campi_testo
freccia_verso l'alto
Amigdala (corpus atugdaloideum) (amigdala) – insieme di cellule della sostanza bianca del lobo temporale. Con aiuto commissura anteriore si collega con il corpo con lo stesso nome dall'altra parte. L'amigdala riceve impulsi da una varietà di sistemi afferenti, compreso il sistema olfattivo, ed è correlata alle reazioni emotive (Fig. 3.36).
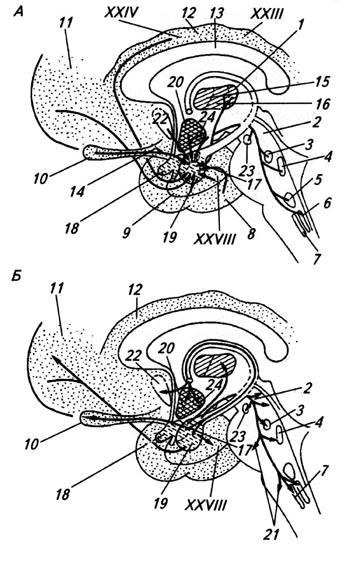 Riso. 3.36.
Riso. 3.36. Riso. 3.36. Strutture cerebrali associate all'amigdala: connessioni afferenti (A) ed efferenti (B) dell'amigdala:
1 - nuclei talamici;
2 – materia grigia periacqueduttale;
3 – nucleo parabrachiale;
4 – punto blu;
5 - nuclei di sutura;
6 – nucleo del tratto solitario;
7 - nucleo dosale del nervo X;
8 – corteccia temporale;
9 – corteccia olfattiva;
10 – bulbo olfattivo;
11 - corteccia frontale;
12 – giro del cingolo;
13 – corpo calloso;
14 – nucleo olfattivo;
15 - anteroventrale e
16 – nucleo dorsomediale del talamo;
17 – centrale,
18 – corticale e
19 – nucleo basolaterale dell'amigdala;
20 – ipotalamo;
21 – formazione reticolare;
22 – partizione;
23 – substantia nigra;
24 – nucleo ventromediale dell'ipotalamo; XXIII, XXIV, XXVIII – campi corticali
Gli emisferi cerebrali sono le aree più grandi del cervello. Emisferi umani grande cervello ha ricevuto il massimo sviluppo rispetto ad altre parti, che distingue in modo significativo il cervello umano e quello animale. Sinistra e emisfero destro Il cervello è separato gli uni dagli altri da una fessura longitudinale che corre lungo la linea mediana. Se guardi la superficie del cervello dall'alto e di lato, puoi vedere una depressione a forma di fessura che inizia 1 cm posteriormente al punto medio tra i poli anteriore e posteriore del cervello e va in profondità. Questo è il solco centrale (rolandico). Sotto di essa, lungo la superficie laterale del cervello corre la seconda grande fessura fessura laterale (Silviana). Funzioni emisfero cerebrale il prosencefalo è l'argomento dell'articolo.
1 106654
Galleria fotografica: Funzioni dell'emisfero cerebrale del prosencefalo
Lobi del cervello
I grandi emisferi sono divisi in lobi, i cui nomi sono dati dalle ossa che li ricoprono: i lobi frontali si trovano davanti alla fessura Rolandica e sopra la fessura Silviana.
Il lobo parietale si trova dietro la porzione centrale e sopra la porzione posteriore del solco laterale; si estende indietro fino al solco parieto-occipitale, lo spazio che lo separa Lobo parietale dall'occipitale, che forma la parte posteriore del cervello.
Il lobo temporale è l'area situata sotto la fessura silviana e delimitata posteriormente da Lobo occipitale sì.
Poiché il cervello cresce rapidamente anche prima della nascita, la corteccia cerebrale inizia ad aumentare la sua superficie, formando pieghe, che portano alla formazione di un caratteristico aspetto cervello somigliante Noce. Queste pieghe sono conosciute come giri e le depressioni che le separano sono chiamate fessure. Alcuni solchi si trovano nello stesso posto in tutte le persone, quindi vengono utilizzati come punti di riferimento per dividere il cervello in quattro lobi.
Sviluppo di giri e solchi
Solchi e convoluzioni iniziano ad apparire al 3-4° mese di sviluppo fetale. Fino a questo punto, la superficie del cervello rimane liscia, come il cervello degli uccelli o degli anfibi. La formazione di una struttura piegata fornisce un aumento della superficie della corteccia cerebrale in condizioni di volume limitato del cranio. Diverse aree della corteccia cerebrale svolgono funzioni specifiche e altamente specializzate. La corteccia cerebrale può essere divisa nelle seguenti aree:
Aree motorie: avviano e controllano i movimenti del corpo. L'area motoria primaria controlla i movimenti volontari del lato opposto del corpo. Direttamente davanti alla corteccia motoria si trova la cosiddetta corteccia premotoria, mentre la terza area, l'area motoria supplementare, si trova sulla superficie interna del lobo frontale.
Le aree sensoriali della corteccia cerebrale percepiscono e sintetizzano le informazioni dai recettori sensoriali in tutto il corpo. L'area somatosensoriale primaria riceve informazioni dal lato opposto del corpo sotto forma di impulsi dai recettori sensoriali per il tatto, il dolore, la temperatura e la posizione delle articolazioni e dei muscoli (recettori propriocettivi).
La superficie del corpo umano ha le sue “rappresentazioni” nelle aree sensoriali e motorie della corteccia cerebrale, che sono organizzate in un certo modo. Il neurochirurgo canadese Wilder Penfield, che esercitò la professione negli anni '50, creò una mappa unica aree sensoriali corteccia cerebrale, che percepiscono le informazioni provenienti da varie parti del corpo. Come parte della sua ricerca, ha condotto esperimenti in cui ha offerto una persona sotto anestesia locale descrivi i tuoi sentimenti nel momento in cui ti ha stimolato certe aree superficie del cervello. Penfield scoprì che la stimolazione del giro postcentrale produceva sensazioni tattili in aree specifiche sul lato opposto del corpo. Altri studi hanno dimostrato che il volume della corteccia motoria, responsabile di varie aree del corpo umano, dipende più dal livello di complessità e precisione dei movimenti eseguiti che dalla forza e dal volume massa muscolare. La corteccia cerebrale è costituita da due strati principali: la materia grigia - un sottile strato di cellule nervose e gliali spesso circa 2-A mm e la materia bianca, che è formata da fibre nervose (assoni) e cellule gliali.
La superficie degli emisferi cerebrali è ricoperta da uno strato materia grigia, il cui spessore in diverse parti del cervello varia da 2 a 4 mm. La materia grigia è formata dai corpi delle cellule nervose (neuroni) e delle cellule gliali che svolgono una funzione di supporto. Nella maggior parte della corteccia cerebrale, al microscopio si possono osservare sei distinti strati di cellule.
Neuroni della corteccia cerebrale
- Le cellule piramidali prendono il nome dalla forma del corpo del neurone, che ricorda una piramide; i loro assoni (fibre nervose) emergono dalla corteccia cerebrale e trasportano informazioni ad altre parti del cervello.
- Le cellule non piramidali (tutte le altre) sono progettate per percepire ed elaborare informazioni provenienti da altre fonti.
Lo spessore dei sei strati di cellule che compongono la corteccia cerebrale varia notevolmente a seconda della regione del cervello. Il neurologo tedesco Korbinian Brodmann (1868-191) esaminò queste differenze colorando le cellule nervose e osservandole al microscopio. Il risultato della ricerca scientifica di Brodmann fu la divisione della corteccia cerebrale in 50 aree separate sulla base di determinati criteri anatomici. Studi successivi hanno dimostrato che i “campi di Brodmann” così individuati svolgono un ruolo specifico ruolo fisiologico e hanno modi unici di interagire.
Cervello (continua)
Il cervello più grande e complesso nei mammiferi è il prosencefalo, o telencefalo, costituito da due emisferi cerebrali (emisferi cerebri). Apparentemente gli emisferi sono sorti principalmente (e forse esclusivamente) in relazione alla ricezione olfattiva. Gli odori non significano molto nella vita dei primati superiori, compresi gli esseri umani. Tuttavia, di più fasi iniziali Nell'evoluzione, fino agli antenati dei vertebrati, l'olfatto è stato il canale principale attraverso il quale gli animali ricevevano informazioni sul mondo che li circondava. Pertanto, è del tutto naturale che i centri olfattivi del cervello fungessero da base su cui costruire il complesso meccanismi nervosi. Già nelle prime fasi dell'evoluzione dei tetrapodi, gli emisferi si trasformano in grandi e importanti centri per la correlazione dei segnali sensoriali. Quando apparvero i mammiferi, la superficie notevolmente espansa degli emisferi divenne il centro associativo dominante, il luogo dell'attività mentale superiore. In diversi rappresentanti della classe, il rapporto tra la massa degli emisferi del prosencefalo e la massa dell'intero cervello varia: nel riccio ( Erinaceus europaeus) è del 48%, per gli scoiattoli ( Sciurus vulgaris) - 53%, nel lupo ( Canis lupus) - 70%, nel lato bianco ( Delphinus delphis) - 75%, nella maggior parte dei primati - 75-80%, nell'uomo - circa 85%. Negli uccelli, gli emisferi cerebrali hanno una massa approssimativamente uguale al resto del cervello o inferiore ad esso, a volte più volte. Infine, l'estrema importanza degli emisferi cerebrali è testimoniata dal fatto che la loro distruzione porta al completo fallimento funzionale del mammifero.
I bulbi olfattivi (bulbi olfactorii) sono adiacenti alla parte anteriore degli emisferi. Queste formazioni sono maggiormente sviluppate negli animali dotati di buon olfatto e sono molto ridotte nelle forme prettamente acquatiche. In base alle differenze nel loro sviluppo, si distinguono diversi tipi di struttura cerebrale. Nei marsupiali, insettivori, edentati, carnivori, roditori e alcuni altri, i bulbi olfattivi sono grandi e sporgono bene quando si guarda il cervello dall'alto. Questo tipo di cervello con un perfetto sviluppo dei lobi olfattivi è detto macrosmatico. Nei pinnipedi, nei sireni e in molti primati i bulbi sono poco sviluppati; Questi animali sono caratterizzati da un cervello microsmatico. Infine, per i cetacei i cosiddetti cervello anosmatico con bulbi olfattivi ridotti. In precedenza si pensava che la capacità di distinguere tra segnali chimici fosse completamente persa nelle balene e nei delfini, ma si scopre che questo non è del tutto vero.
Gli strati superficiali degli emisferi del prosencefalo dei mammiferi formano il pallio, o volta midollare (pallio). Lo strato superiore, costituito dai corpi dei neuroni e senza polpa fibre nervose, chiamata corteccia (cortex cerebri) ed è la materia grigia del fornice. I corpi dei neuroni si trovano nella corteccia a strati, formando particolari strutture schermo. Questa organizzazione del cervello consente di rappresentare spazialmente il mondo esterno sulla base delle informazioni provenienti dai sensi. Le strutture dello schermo sono caratteristiche dei centri cerebrali più importanti dei mammiferi, mentre negli altri vertebrati sono meno comuni, soprattutto nei centri visivi. Sotto la corteccia c'è uno strato di processi neuronali mielinizzati: la sostanza bianca del fornice. Le fibre nervose della sostanza bianca formano fasci conduttivi che saldano gli emisferi: le commissure cerebrali. Sebbene la maggior parte della materia grigia sia concentrata nella corteccia, è presente anche nello strato sottocorticale, dove è rappresentata da cluster relativamente piccoli: i nuclei. Questi includono i corpi striatali (corpora striata), che si trovano sotto la cavità del ventricolo laterale e hanno ricevuto il loro nome a causa delle fibre nervose che li attraversano. Sotto il controllo della corteccia, lo striato svolge la funzione di regolare le reazioni stereotipate e automatizzate - riflessi incondizionati.
Schemi che riflettono il progressivo sviluppo degli emisferi del prosencefalo. Vista laterale dell'emisfero con il bulbo olfattivo. Vari dipartimenti, diversi citologicamente tra loro, si colorano diversamente.
R: l'emisfero non è altro che il lobo olfattivo.
B - si differenziano la sezione dorsale - archipallio (= ippocampo) e la sezione ventrale - nucleo basale (striato).
B - i gangli della base si sono spostati nella regione interna dell'emisfero.
D - appare una piccola area, che rappresenta un neopallio.
D - il paleopallio è spinto sulla superficie mediale dell'emisfero, ma il neopallio è ancora di dimensioni modeste e sotto il solco olfattivo sono conservate sezioni olfattive notevolmente sviluppate.
E - Il reparto olfattivo primitivo è conservato solo nella regione ventrale, ed il neopallio raggiunge uno sviluppo fortissimo. (Dopo Romer e Parsons, 1992.) La volta cerebrale cominciò a formarsi nei pesci. A causa del progressivo sviluppo dell'olfatto sistema sensoriale sviluppano un paleopallium, o arco antico (paleopallium), che ricopre completamente i piccoli emisferi. Allo stadio di pesce con pinne lobate, l'archipallium, o arco vecchio (archipallium), appare nella parte dorsale degli emisferi più vicino all'asse del corpo. Negli anfibi e nei rettili primitivi si sviluppa ulteriormente, a seguito della quale il paleopallio viene messo da parte e si conserva solo lungo la superficie laterale degli emisferi. Allo stesso tempo, il paleopallio continua ad essere prevalentemente di natura olfattiva e negli stadi evolutivi più elevati forma i lobi olfattivi della corteccia cerebrale. L'archipallio è in una certa misura, anche se piccola, un centro correlativo, che riceve fibre ascendenti dal diencefalo, nonché fibre dal bulbo olfattivo e dal lobo olfattivo; sembra anche essere associato al comportamento emotivo. Il percorso neurale da questa regione all'ipotalamo costituisce l'elemento principale di un fascio di fibre chiamato fornice nei mammiferi.
Negli anfibi compare per la prima volta il rudimento del neopallium, o nuovo arco (neopallium). Nei rettili il neopallio forma già una piccola area tra la volta antica e quella vecchia. Fin dall'inizio della sua evoluzione, quest'area è stata un centro associativo, come i gangli della base, ricevendo fibre che trasmettono segnali sensoriali dal tronco cerebrale ad esso e, al contrario, trasmettono comandi direttamente alle colonne motorie.
Nei monotremi il neopallium è ancora incuneato tra il paleopallium all'esterno e l'archipallium all'interno. Nei marsupiali cresce oltre il tetto e le pareti laterali degli emisferi. In questo caso, l'archipallio è spostato sulla superficie mediale e il paleopallio è limitato alla parte ventrolaterale dell'emisfero, situata sotto il solco nasale (fissura rhinalis), un solco che rappresenta il confine tra le aree olfattive e non olfattive. della corteccia. Nella placenta, a causa di un'ulteriore complicazione ed espansione del neopallio, gli emisferi raggiungono dimensioni tali da superare in volume il resto delle parti cerebrali messe insieme. Gli emisferi crescono all'indietro e ai lati, ricoprendo successivamente il diencefalo, mesencefalo e parte del cervelletto. I ventricoli accoppiati e le antiche strutture responsabili principalmente dell'olfatto (bulbi olfattivi, vecchi strati della volta midollare e relativi fasci e nuclei nervosi) vengono respinti e deformati nel cervello placentare. Pertanto, il paleopallio è conservato sulla superficie ventrale dell'emisfero sotto forma di una piccola area olfattiva chiamata lobo piriforme (lobus piriformis), e l'archipallio è ripiegato in profondità nella piega del lobo temporale in un rotolo chiamato ippocampo ( ippocampo). La superiorità degli emisferi cerebrali rispetto alle altre parti del cervello è evidente in tutti i mammiferi, ma è particolarmente pronunciata nelle forme avanzate, come l'uomo. Gli emisferi dominano anche dal punto di vista funzionale. A differenza dei mammiferi, la complicazione del prosencefalo negli uccelli si esprime principalmente nella crescita dei nuclei basali (nuclei basales), piuttosto che nel rimanente fornice sottile. 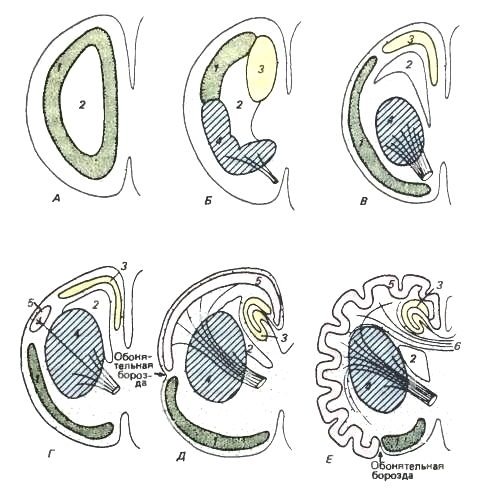
Sezioni schematiche dell'emisfero proencefalo sinistro. Le designazioni dei colori sono le stesse della figura precedente.
1 - paleopallio; 2 - ventricolo laterale; 3 - archipallio; 4 - nuclei basali; 5 - neopallio; 6 - corpo calloso.
A - stadio primitivo. L'emisfero, in sostanza, è il lobo olfattivo. La materia grigia scarsamente differenziata si trova all'interno del cervello.
B - stadio osservato negli anfibi moderni. La materia grigia si trova ancora lontano dalla superficie esterna, ma è già divisa in paleopallio (= lobo olfattivo), archipallio (= ippocampo) e gangli della base (= striato). Quest'ultimo acquisisce il significato di un centro associativo che ha connessioni afferenti ed efferenti con il talamo (rappresentato da linee che simboleggiano fasci di fibre tagliate).
B - uno stadio più progressivo, in cui i gangli della base sono immersi all'interno dell'emisfero, mentre le sezioni della corteccia si sono spostate leggermente verso l'esterno.
G è lo stadio in cui si trovano i rettili avanzati. Appare il neopallio.
D - stadio di un mammifero primitivo. Neopallio aumentato. Ha estese connessioni con il tronco encefalico. Archipallium su superficie mediale l'emisfero si avvolge come l'ippocampo. Il paleopallio è ancora molto sviluppato.
E - stadio di un mammifero altamente organizzato. Il Neopallium cresce estremamente e si raccoglie in pieghe. Il paleopallio occupa una regione ventrale limitata, che rappresenta un lobo a forma di pera. Si sviluppa il corpo calloso, un potente ponte che collega le aree del neopallio dei due emisferi. (Dopo Romer e Parsons, 1992.) La corteccia del neopallio è chiamata neocorteccia o neocorteccia. Nei mammiferi funge da centro del riflesso superiore (riflesso condizionato) attività nervosa, coordinando il lavoro di altre parti del cervello. Da qui gli impulsi vengono inviati a vari organi e tessuti del corpo e qui viene effettuata la regolazione processi fisiologici in base alle condizioni ambientali. È la nuova corteccia che accumula tracce di singole eccitazioni e delle loro combinazioni, a seguito delle quali la memoria operativa si arricchisce, fornendo la capacità di scegliere soluzioni ottimali in nuove situazioni. Nella maggior parte dei casi, queste decisioni rappresentano nuove combinazioni di elementi comportamentali precedentemente noti, ma vengono anche sviluppate e consolidate nuove opzioni di azione. Man mano che si sviluppa, la nuova corteccia non solo assume le funzioni di un centro correlativo e associativo di nuovi tipi emergenti di attività nervosa superiore, ma inizia anche a svolgere molte funzioni che in precedenza appartenevano ai centri del tronco cerebrale e dei gangli della base. In questo caso gli antichi centri che controllano gli atti istintivi non vengono eliminati, ma sono solo soggetti ad un controllo superiore.
In connessione con lo sviluppo della nuova corteccia, il tetto del mesencefalo perde il suo antico significato, rimanendo solo un centro di riflesso e di trasmissione. Gli impulsi sensoriali uditivi e altri somatici vengono trasmessi in avanti al talamo, dove la maggior parte delle fibre visive vengono interrotte, e tutti questi segnali dal talamo vengono trasmessi agli emisferi lungo potenti fasci nervosi. Simili connessioni talamiche con i gangli della base sono sorti in gruppi di vertebrati poco organizzati e sono stati maggiormente sviluppati negli uccelli. A differenza degli uccelli, nei mammiferi la maggior parte delle fibre passa attraverso lo striato e diverge verso la superficie della neocorteccia. Ad esso affluisce così un insieme completo di dati sensoriali, sulla base dei quali nella corteccia vengono prese le corrispondenti “decisioni” motorie.
Come già accennato, alcuni segnali vengono trasmessi dalla corteccia al cervelletto attraverso il ponte e forniscono i necessari effetti regolatori. La neocorteccia ha collegamenti anche con lo striato e perfino con l'ipotalamo - e quindi con il sistema autonomo sistema nervoso. Tuttavia, la maggior parte dei comandi motori viene inviata lungo il percorso piramidale (tractus corticospinalis), un percorso speciale fascio nervoso, che direttamente, senza deviazioni, va dalla corteccia cerebrale attraverso il mesencefalo alle aree motorie somatiche del tronco encefalico e midollo spinale. In questo caso, le fibre della parte laterale di questo percorso si intersecano e si innervano il lato opposto corpo (cioè la fibra sinistra innerva lato destro corpo, e viceversa), e le fibre ventrali rimangono collegate al loro lato del corpo. Il tratto piramidale è presente solo nei mammiferi, il che dimostra chiaramente la posizione dominante in essi della neocorteccia. Questa struttura raggiunge il suo massimo sviluppo nelle scimmie e, soprattutto, nell'uomo, svolgendo un ruolo importante nella deambulazione eretta. Nei marsupiali, gli assoni piramidali raggiungono solo toracico, e nei monotremi il percorso piramidale è completamente assente. 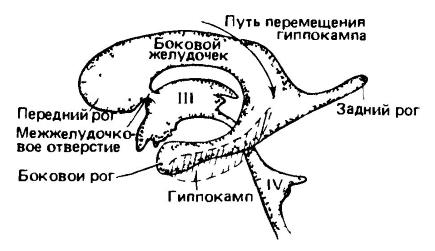
Ventricoli del cervello umano; vista dal lato sinistro. I ventricoli sono rappresentati come un calco, ma il tessuto cerebrale non è raffigurato. Con la crescita dell'emisfero proencefalo, il ventricolo laterale si allarga all'indietro per formarsi corno posteriore nel lobo occipitale e nella sua parte laterale - in basso e in avanti per formare il corno laterale nel lobo temporale. Crescite simili, dirette all'indietro e verso il basso, hanno portato a cambiamenti nella posizione varie parti cervello L'ippocampo, che si sviluppava in posizione dorsale sulla superficie mediale dell'emisfero, si spostava indietro e verso il basso in posizione ventrale nei mammiferi altamente sviluppati. (Dopo Romer e Parsons, 1992.) Poiché la neocorteccia è un sottile foglio di materiale cellulare stratificato sottostante la massa fibrosa bianca del cervello, il semplice aumento del volume degli emisferi non può produrre una crescita proporzionata della corteccia. Allo stesso tempo, nelle forme avanzate l'area della corteccia può aumentare notevolmente a causa del suo ripiegamento. Le pieghe così formate sono chiamate convoluzioni (giri) e gli spazi profondi tra loro sono chiamati solchi (solchi). Entrambi contengono componenti morfologici comuni. Nel caso più semplice, c'è una profonda fessura silviana che separa il lobo frontale (lobus frontalis) dal lobo temporale (lobus temporalis). Quindi, sopra e anteriormente alla fessura silviana, appare una fessura Rolandica che corre trasversalmente, separando il lobo frontale dal lobo parietale (lobus parietalis) sovrastante. Nei primati, un solco trasversale separa il piccolo lobo occipitale posteriore (lobus occipitalis). Oltre ai solchi principali si formano molti solchi aggiuntivi; il loro numero è particolarmente elevato nei primati e nelle balene dentate. In precedenza, si credeva che in alcuni casi i solchi segnassero i confini morfologici corrispondenti a determinate aree della corteccia. Tuttavia, ulteriori studi hanno mostrato l'assenza di una connessione fissa tra la distribuzione del ripiegamento e unità strutturale corteccia (ad eccezione del solco nasale e, in una certa misura, del solco centrale nei primati, di cui parleremo più avanti). È interessante notare che il ripiegamento corticale si è sviluppato in modo completamente indipendente in diversi tronchi evolutivi dei mammiferi. Nei mammiferi relativamente primitivi, come i monotremi, i marsupiali e alcuni placentati (insettivori, chirotteri, roditori, lagomorfi), la corteccia è più modestamente sviluppata e ha una superficie liscia. 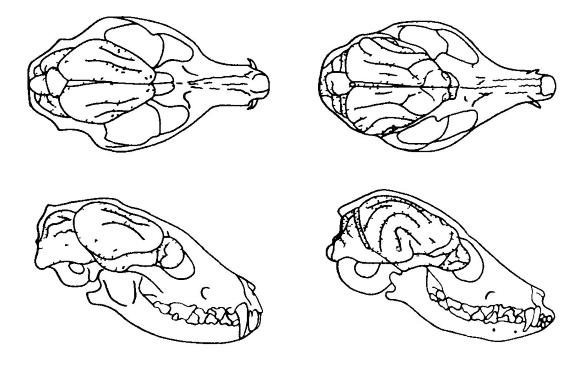
La posizione del cervello nel cranio dei canidi fossili e viventi. C'è un notevole aumento delle dimensioni e della complessità del cervello, in particolare degli emisferi del prosencefalo. Esperocione ( Hesperocyon gregarius) (a sinistra) - una forma oligocenica vissuta circa 30 milioni di anni fa. Fennec ( Vulpes zerda) (sulla destra) - forma moderna dimensioni simili. (Dopo Romer e Parsons, 1992.) La materia grigia della neocorteccia è caratterizzata da una struttura istologica complessa. Nei mammiferi placentari, ci sono 6 strati di cellule sovrapposte e fibre incastonate tra di loro; questo distingue notevolmente la neocorteccia dalle aree preservate della corteccia del paleopallio e dell'archipallio, dove si possono distinguere solo da 2 a 4 strati di cellule. Secondo le stime esistenti, i mammiferi con soprattutto cervello grande, il numero di cellule nella neocorteccia può raggiungere miliardi.
La sostanza bianca, situata sotto la materia grigia, oltre al ventaglio di connessioni che vanno dalla corteccia alle parti sottostanti del cervello e ritorno, comprende un numero enorme di fibre trasversali intrecciate che collegano diverse aree della corteccia stessa. La commissura così formata è allungata all'indietro (secondo la velocità di crescita degli emisferi) ed è divisa in due placche fuse lungo il bordo posteriore. Quello inferiore, più sottile e deviato verso il basso dal bordo anteriore, è il fornice (fornix), la commissura della corteccia dell'archipallio (cioè l'ippocampo). La commissura orizzontale superiore, più spessa, appartiene alla nuova corteccia ed è chiamata corpo calloso. Questa educazione consente di combinare la memoria di entrambi gli emisferi e aumenta significativamente la capacità di apprendimento del cervello. Il corpo calloso è presente solo nelle placentari a causa del notevole sviluppo della neocorteccia; i monotremi e i marsupiali ne sono sprovvisti. Inoltre, tutti i mammiferi hanno una commissura anteriore (commissura anteriore), che collega le aree olfattive della corteccia. 
Disposizione strato per strato delle cellule nervose nella corteccia telencefalo mammifero (secondo Naumov e Kartashev, 1979). Il complesso sistema di "conduttori" che collega tutte le parti della corteccia suggerisce che la materia grigia è, in linea di principio, educazione unificata, tutte le cui parti hanno le stesse capacità di svolgere qualsiasi funzione degli emisferi cerebrali. In una certa misura questo è vero: gli esperimenti dimostrano che negli animali da laboratorio è possibile distruggere una parte significativa della nuova corteccia senza causare danni permanenti. attività normale. I dati sugli infortuni e sulla morbilità confermano che ciò vale anche per cervello umano. Allo stesso tempo, è chiaro che alcune aree della corteccia sono normalmente associate allo svolgimento di funzioni molto specifiche. Sopra sono state menzionate le aree del paleopallium e dell'archipallium, destinate principalmente all'analisi delle informazioni olfattive e conservate, rispettivamente, sotto forma di lobo piriforme e ippocampo. La differenziazione delle singole aree avviene anche nella corteccia del neopallio. La parte anteriore degli emisferi contiene l'area motoria. Situato qui Lobo frontale tra le altre cose, controlla la comunicazione animale, compresa la comunicazione acustica; nell'uomo è associato alla parola, cioè il secondo sistema d'allarme. La parte posteriore degli emisferi è associata alla percezione delle sensazioni. Nell'occipitale e Lobi Temporali Ci sono aree che controllano rispettivamente la vista e l'udito. Più avanti, vicino all'area motoria, ci sono le aree che ricevono segnali tattili e propriocettivi. Nei primati, il solco centrale (sulcus centralis), attraversando l'apice dell'emisfero dalla superficie mediale a quella laterale, delimita (sebbene non del tutto accuratamente) l'area motoria da quella sensoriale. Lungo il bordo anteriore del solco centrale si trovano in ordine lineare aree motorie specifiche che servono ciascuna parte del corpo e degli arti. Lungo il bordo posteriore del solco centrale si trovano nello stesso ordine le aree di percezione sensoriale delle parti corrispondenti del corpo.
Pertanto, in molti mammiferi quasi tutta la superficie della neocorteccia è occupata da aree più o meno strettamente associate a determinati aspetti sensoriali o funzioni motorie. Sebbene il solco centrale possa essere assente, la placenta generalmente mantiene una disposizione lineare simile delle aree sensoriali e motorie una di fronte all'altra. Nei marsupiali (e tra i placentati, negli xenoartridi), la “marcatura” delle aree corporee è più o meno la stessa, ma le aree sensoriali non sono separate da quelle motorie, ma si alternano con esse. Ma, ad esempio, negli esseri umani, queste aree funzionali specifiche occupano relativamente poco spazio sulla superficie della neocorteccia. Tra di loro sorsero vaste zone di materia grigia (una di queste aree particolarmente grande occupa maggior parte lobo frontale) che non sono associati a specifiche funzioni sensoriali o motorie. Pertanto, queste aree sono spesso chiamate “punti bianchi”, anche se, come dimostra il danno a queste aree, sono dove si trovano le nostre capacità mentali superiori, tra cui l’apprendimento, l’iniziativa, la lungimiranza e il giudizio. Allo stesso tempo, ci sono anche aree che possono essere rimosse senza gravi conseguenze per l’attività intellettuale. 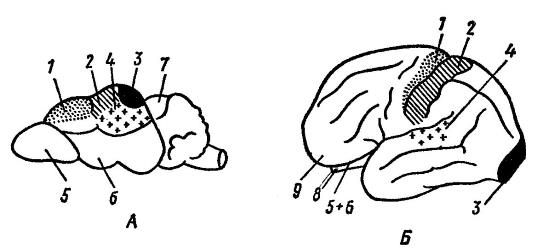
Centri funzionali della corteccia cerebrale del toporagno ( Sorex sp.) (A) e umano ( Homo sapiens
) (B) (secondo Naumov e Kartashev, 1979):
1 - centro motorio; 2 - centro della sensibilità muscolocutanea; 3 - centro visivo; 4- centro uditivo; 5 - bulbo olfattivo; 6 - lobi olfattivi; 7 - tetto del mesencefalo; 8 - cervelletto; 9 - lobo frontale. L'evoluzione del cervello è fortemente influenzata da ambiente esterno e l'attività motoria (procurarsi il cibo, difensiva). Allo stesso tempo, lo sviluppo di varie parti del cervello è determinato principalmente dai metodi di ricerca del cibo: in un cane ( Canis lupus), utilizzando l'olfatto in questo processo, l'area olfattiva è più sviluppata; in un gatto ( Felis silvestris), ricerca del cibo mediante visione - visiva; nel macaco ( Macaca mulatta), utilizzando la vista e l'udito: visivo e uditivo.
Di solito si presume che la dimensione degli emisferi cerebrali determini differenze nelle capacità mentali dei diversi mammiferi. In un certo senso questo è vero, ma con notevoli riserve. Il cervello più grande è composto da Di più cellule nervose. Se la superficie della corteccia è in qualche modo correlata all’intelligenza, allora è ovvio che di due cervelli della stessa dimensione, quello più sviluppato sarà quello con la superficie scanalata, e quello meno sviluppato sarà quello con la superficie scanalata. quello con una superficie liscia. Anche la dimensione dell'animale stesso influisce sul volume del cervello. Ciò accade se non altro perché il cervello deve avere aree più grandi per servire connessioni sensoriali e motorie più estese. Tuttavia, l’aumento delle dimensioni del cervello non è del tutto proporzionale alla massa corporea, quindi gli animali di grandi dimensioni tendono ad avere cervelli relativamente più piccoli senza alcuna apparente perdita di capacità mentale. Pertanto, la dimensione assoluta del cervello non è un criterio assoluto di intelligenza. Ciò è certamente indicato dal fatto che il cervello della balena può avere un volume cinque volte più grande del cervello umano.
Confronto tra il cervello di alcuni mammiferi:
1 - cavallo; 2 - cane; 3 - canguro; 4 - persona; 5 - elefante. Quota percentuale cervello nella massa dell'intero corpo è chiamato indice di cefalizzazione. Nei grandi insettivori è circa lo 0,6%, in quelli piccoli - fino all'1,2%, nei grandi cetacei - circa 0,3% e in quelli piccoli - fino all'1,7%. Nella maggior parte dei primati, l'indice di cefalizzazione è dell'1-2%. Negli esseri umani raggiunge il 2-3% e alcune piccole scimmie dal naso largo hanno un cervello la cui massa costituisce fino al 7% del loro peso corporeo. Allo stesso tempo, nei rettili e negli uccelli moderni l'indice di cefalizzazione varia dallo 0,05 allo 0,5%.
Quella che segue è la massa cerebrale di alcuni mammiferi (la massa dell'animale è indicata tra parentesi):
Opossum della Virginia ( Didelphis virginiana) - 7,6 g (5 kg);
koala ( Phascolarctos cinereus) - 19,2 g (8 kg);
elefante della savana ( Loxodonta africana) - 6000 g (5000 kg);
riccio comune ( Erinaceus europaeus) - 3,3 g (1 kg);
topo domestico ( Mus musculus) - 0,3 g (0,02 kg);
ratto grigio ( Rattus norvegicus) - 2 g (0,3 kg);
scoiattolo comune ( Sciurus vulgaris) - 7 g (0,4 kg);
Coniglio europeo ( Oryctolagus cuniculus) - 11 g (3 kg);
cavallo domestico ( Equus ferus) - 530 g (500 kg);
rinoceronte nero ( Diceros bicornis) - 500 g (1200 kg);
Cervo dalla coda bianca ( Odocoileus virginianus) - 500 g (200 kg);
giraffa ( Giraffa camelopardalis) - 680 g (800 kg);
pecore domestiche ( Ovis orientalis) - 140 g (55 kg);
toro domestico ( Bos primigenius) - 490 g (700 kg);
Cammello battriano ( Camelus bactrianus) - 762 g (700 kg);
ippopotamo ( Ippopotamo anfibio) - 580 g (3500 kg);
delfino comune ( Delphinus delphis) - 815 g (60 kg);
narvalo ( Monodonte monocero) - 2997 g (1578 kg);
capodoglio ( Fiseter macrocefalo) - 8028 g (35833 kg);
balena Blu ( Balaenoptera musculus) - 3636 g (50900 kg);
gatto domestico ( Felis silvestris) - 25 g (3 kg);
un leone ( Panthera leone) - 270 g (250 kg);
volpe comune ( Vulpes vulpes) - 53 g (4,5 kg);
cane domestico ( Canis lupus) - 64 g (10 kg);
orso polare ( Ursus maritimus) - 500 g (700 kg);
tricheco ( Odobenus rosmarus) - 1130 g (700 kg);
marmotta Geldi ( Callimico goeldii) - 7 g (0,2 kg);
cappuccino dalla fronte bianca ( Cebus albifrons) - 57 g (1 kg);
macaco rhesus ( Macaca mulatta) - 88 g (6,5 kg);
babbuino ( Papio cinocefalo) - 200 g (25 kg);
gibbone d'argento ( Moloch ilobate) - 112 g (6,5 kg);
Orango del Kalimantan ( Pongo pygmaeus) - 413 g (50 kg);
gorilla occidentale ( Gorilla gorilla) - 506 g (126 kg);
scimpanzé comune ( Pan trogloditi) - 430 g (55 kg);
persona ragionevole ( Homo sapiens) - 1400 g (72 kg).
Dagli esempi sopra riportati è chiaro che nei mammiferi più piccoli il cervello è quasi sempre relativamente più grande e, all’aumentare della dimensione del corpo dell’animale, la dimensione relativa del cervello diminuisce. Ciò è particolarmente pronunciato tra le specie di mammiferi strettamente imparentate, ad esempio nei gatti ( Felis silvestris) e il leone ( Panthera leone). Anche i cani di varie razze sono molto convincenti in questo senso. Se la massa corporea è la più piccola e la più razze di grandi dimensioni sono approssimativamente nel rapporto di 1:33, quindi le masse cerebrali delle stesse razze sono correlate come 1:3. 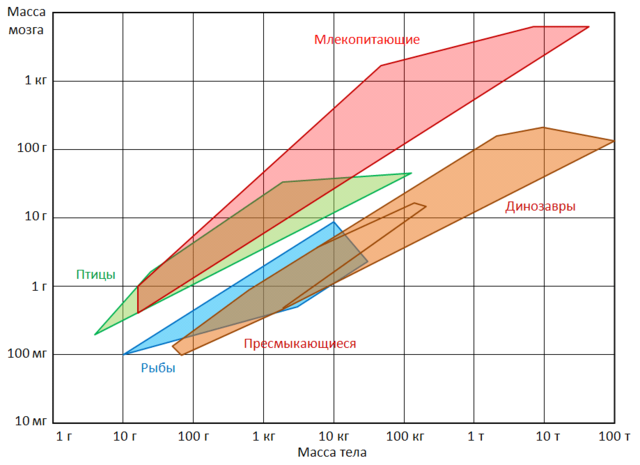
Intervalli di valori di massa cerebrale e corporea per alcuni gruppi di vertebrati. Negli animali domestici, privati della necessità di procurarsi il cibo e di difendersi dai nemici, le dimensioni del cervello sono notevolmente ridotte. Ad esempio, il volume del cervello di un lupo ( Canis lupus) 30% in più rispetto ad un cane della stessa taglia. È interessante notare che questi cambiamenti riguardano non solo gli animali tradizionalmente domestici, ma anche i rappresentanti delle specie a vita libera tenute in cattività per qualche tempo. Sì, volpi ( Vulpes vulpes), nati in natura, ma fin dai primi giorni di vita in cattività, hanno un cervello più piccolo rispetto ai loro parenti che vivono in condizioni naturali. In questo caso, le differenze raggiungono il 20%, che corrisponde approssimativamente alla differenza nel volume del cervello degli animali selvatici e dei veri animali domestici. Il restringimento del cervello, sebbene non così pronunciato (circa il 5%), è stato riscontrato nei lupi in cattività ( Canis), furetti ( Mustela), ratti ( Ratto). In questo caso, la diminuzione non colpisce tutte le parti del cervello, ma solo quelle aree associate al funzionamento degli organi di senso. La cosa più notevole è che il peso del cervello degli animali domestici liberati aumenta. Ad esempio, i gatti selvatici hanno cervelli che sono circa il 10% più grandi rispetto ai loro omologhi domestici. Un significativo ingrandimento del cervello è stato riscontrato anche nei conigli selvatici ( Oryctolagus cuniculus) sulle Isole Kerguelen. Asini selvatici ( Equus asinus)V Sud America aveva il 15% grande cervello rispetto a quelli fatti in casa. È anche interessante che il cervello di un uomo di Neanderthal ( Homo neanderthalensis) e l'homo sapiens paleolitico ( Homo sapiens) erano leggermente più grandi del cervello degli esseri umani moderni.
È stato accertato che molti mammiferi presentano asimmetria motoria, cioè l'uso predominante della metà destra o sinistra del corpo. Ad esempio, quando si studiano i cavalli non domati ( Equus ferus) hanno registrato con quale gamba gli animali iniziano a camminare, da quale lato preferiscono aggirare gli ostacoli e da quale lato preferiscono sdraiarsi in una stalla sul fieno. Di conseguenza, la maggior parte delle fattrici erano destrimani e la maggior parte degli stalloni erano mancini. Circa il 10% dei cavalli non mostrava alcuna preferenza né per l’arto destro né per quello sinistro. Secondo le osservazioni, circa il 90% dei trichechi ( Odobenus rosmarus) estrai i molluschi dal fango marino con la pinna destra. Cullando i bambini, circa l'80% delle femmine di scimpanzé ( Padella) e gorilla ( Gorilla) premono la testa sul lato sinistro del petto (circa la stessa percentuale si osserva tra le donne). Ratti ( Ratto), alla ricerca di cibo utilizzando le vibrisse situate su lato destro museruole, sono più prede dei loro parenti mancini.

