14.1. DISPOSIZIONI GENERALI
Telencefalo, o cervello, situato nello spazio sopratentoriale della cavità cranica e è composto da due grandi
emisferi (gemispherium cerebrale),separati tra loro da una profonda fessura longitudinale (fissura longitudinalis cerebrale), in cui è immersa la falce grande cervello (falce cerebrale), che rappresenta una duplicazione del solido meningi. Gli emisferi cerebrali costituiscono il 78% della sua massa. Ciascuna di emisferi cerebrali Esso ha lobi: frontale, parietale, temporale, occipitale e limbico. Coprono le strutture del diencefalo, del tronco cerebrale e del cervelletto situati sotto il tentorio cerebellare (sottotentoriale).
Sebbene il cervello umano nel suo insieme possa svolgere molti compiti e concentrarsi su quelli più specifici rispetto ad altri, la verità è che l'apparato mentale umano è più focalizzato sulla specializzazione, cioè che parti di ciascun emisfero sono focalizzate su compiti specifici Nel caso delle donne, vi è un'interazione più integrale utilizzando entrambi gli emisferi Di più attività, che si chiama multitasking.
Segmentazione delle informazioni
Nel cervello maschile l’informazione è divisa in gruppi o segmenti non necessariamente collegati tra loro. Nel femminile tutto è connesso come un sistema. Questo è il motivo per cui le donne tendono ad essere olistiche. A causa di queste differenze tra il cervello di uomini e donne, gli uomini tendono a concentrarsi completamente sulle attività che stanno svolgendo senza essere distratti da altri aspetti. Le donne, invece, osservano ogni evento del mondo come un quadro completo, in cui ogni parte interagisce.
Ciascuno degli emisferi cerebrali ha tre superfici: superolaterale o convesso (Fig. 14.1a), - convesso, rivolto verso le ossa della volta cranica; interno (Fig. 14.1b), adiacente al grande processo falciforme, e inferiore, o basale (Fig. 14.1c), che ripete il rilievo della base del cranio (le sue fosse anteriore e media) e il tentorio del cervelletto. In ciascun emisfero ci sono tre bordi: superiore, inferiore interno e inferiore esterno, e tre poli: anteriore (frontale), posteriore (occipitale) e laterale (temporale).
Sentimenti tra un uomo e una donna
Ci sono anche differenze tra uomini e donne nei sentimenti e nella percezione del mondo. Negli esseri umani l'apparenza è la funzione dominante, il modo in cui la maggior parte delle informazioni entra nel sistema nervoso. Nelle donne tutti i sistemi funzionano insieme, come le persone cieche, hanno l'udito, l'olfatto e il tatto molto sviluppati, ma a questi si aggiunge la vista.
Anche le dimensioni del cervello differiscono tra uomini e donne, dove tendono ad essere leggermente più grandi. Ciò non significa che siano più intelligenti, perché gli scienziati ritengono che lo scopo di questa espansione sia legato al maggiore volume muscolare maschile e non ai processi mentali. Gli uomini hanno 20 volte più testosterone nel corpo rispetto alle donne. Ecco perché il suo sistema nervoso si manifesta in relazioni più aggressive e dominanti, e perché il sesso è fisicamente più desiderabile. Le donne sono più delicate e la loro mente ricerca il piacere sessuale, concentrandosi sulla sensualità e sull'affetto.
Cavità di ciascuno emisfero cerebrale ammonta a ventricolo laterale del cervello, in questo caso, il ventricolo laterale sinistro viene riconosciuto come il primo, quello destro come il secondo. Il ventricolo laterale ha una parte centrale situata in profondità nel lobo parietale (lobo parietale) e da esso partono tre corna: corno anteriore penetra nel lobo frontale (lobo frontale), inferiore - al temporale (lobo temporale), posteriore - all'occipitale (lobo occipitale). Ciascuno dei ventricoli laterali comunica con il terzo ventricolo del cervello attraverso l'interventricolare Buco di Monroe.
Emisferi cerebrali e abilità cognitive
E infine, diremo che negli esseri umani l'emisfero sinistro del cervello ha una maggiore dominanza. Nel caso delle donne c’è un equilibrio nell’uso di entrambi gli emisferi, sinistro e destro. Ecco perché nelle abilità cognitive possiamo osservare che gli uomini hanno una maggiore tendenza ad essere bravi in matematica e nei calcoli, mentre le donne hanno una facilità per il linguaggio, attività che richiede l'utilizzo di più aree interne.
Ci mostrano come, nel processo evolutivo, funziona sistema nervoso entrambi i sessi erano specializzati per garantire il futuro della specie. Non dimenticare mai che le differenze sono legate alle strutture fisiche e mentali e che nessuno dei due deve sottovalutare il valore umano di ciascun genere. Siamo diversi, ma ci completiamo a vicenda.
Le sezioni centrali della superficie mediale di entrambi gli emisferi sono collegate tra loro da commissure cerebrali, la più massiccia delle quali è il corpo calloso, e da strutture del diencefalo.
Il telencefalo, come altre parti del cervello, è costituito da grigio e materia bianca. La materia grigia si trova in profondità in ciascun emisfero, formando lì nodi sottocorticali e lungo la periferia delle superfici libere dell'emisfero, dove costituisce la corteccia cerebrale.
Nel libro Deficienze di attenzione nei bambini e negli adolescenti, una delle cose menzionate dall'autrice Amanda Cespedes è l'importanza di conoscere il cervello e le sue funzioni come strategia per comprendere e rispondere ai bisogni dei bambini e degli adolescenti. In sostanza, l’apprendimento acquisisce nuove conoscenze. Per raggiungere questo obiettivo, entrano in gioco alcune funzioni cerebrali di base, da quelle cognitive a quelle emotive. In particolare, studiare cervello umano nell'infanzia basata sull'esistenza reti neurali, in cui le informazioni vengono elaborate nello stesso modo in cui un computer elabora i dati.
Domande di base relative alla struttura, alle funzioni dei nodi sottocorticali e alle opzioni quadro clinico quando sono colpiti, sono discussi nei capitoli 5, 6. La corteccia cerebrale ha un'area di circa
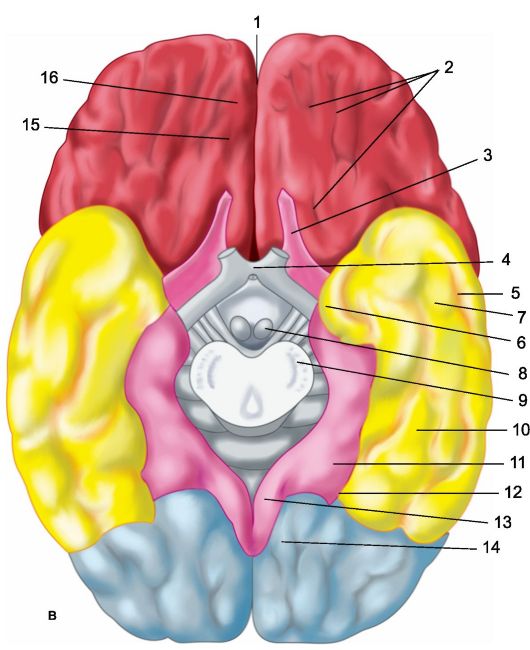
Riso. 14.1.Emisferi del cervello.
Ciò significa che il cervello del bambino non si limita a ricevere informazioni, ma in questo processo centinaia di migliaia di neuroni diventano disponibili per l'apprendimento e il processo informativo che avviene nei due emisferi del cervello: quello destro e quello sinistro. Nel libro ADHD Disabilities in Children and Adolescents di Amanda Cespedes, l'autrice approfondisce questo argomento, evidenziando le caratteristiche di ciascun emisfero e dando esempi specifici situazioni o azioni in cui il funzionamento di entrambi gli emisferi si manifesta nei bambini e negli insegnanti, perché?
Perché solo attraverso una profonda comprensione del cervello è possibile adattare le strategie di apprendimento alle esigenze specifiche. Matura prima della nascita e rafforza le sue reti funzionali durante i primi cinque anni di vita. In questo emisfero si possono distinguere sei talenti: visuoscopico, logica spaziale, ritmo melodico, cinesico, interpersonale e naturalistico. Emisfero destro non è governato dall'analisi oggettiva della realtà esterna e quindi consente l'esistenza di realtà soggettive che promuovono l'immaginazione.
a - superficie superolaterale dell'emisfero sinistro: 1 - solco centrale; 2 - parte orbitale del giro frontale inferiore; I - lobo frontale; 3 - giro precentrale; 4 - solco precentrale; 5 - giro frontale superiore; 6 - giro frontale medio; 7 - parte tegmentale del giro frontale inferiore; 8 - giro frontale inferiore; 9 - scanalatura laterale; II - lobo parietale: 10 - giro postcentrale; 11 - solco postcentrale; 12 - solco intraparietale; 13 - giro sopramarginale; 14 - giro angolare; III - lobo temporale: 15 - giro temporale superiore; 16 - solco temporale superiore; 17 - giro temporale medio; 18 - solco temporale medio; 19 - giro temporale inferiore; IV - lobo occipitale: b - superficie mediale dell'emisfero destro: 1 - lobo paracentrale, 2 - precuneo; 3 - solco parieto-occipitale; 4 - cuneo, 5 - giro linguale; 6 - giro occipitotemporale laterale; 7 - giro paraippocampale; 8 - gancio; 9 - volta; 10 - corpo calloso; 11 - giro frontale superiore; 12 - giro del cingolo; c - superficie inferiore degli emisferi cerebrali: 1 - fessura interemisferica longitudinale; 2 - scanalature orbitali; 3 - nervo olfattivo; 4 - chiasma visivo; 5 - solco temporale medio; 6 - gancio; 7 - giro temporale inferiore; 8 - corpo mastoideo; 9 - base del peduncolo cerebrale; 10 - giro occipitotemporale laterale; 11 - giro paraippocampale; 12 - solco collaterale; 13 - giro del cingolo; 14 - giro linguale; 15 - solco olfattivo; 16 - giro dritto.
Lo stile cognitivo di quest'area del cervello è di tipo eterogeneo, il che significa che procede da un'idea iniziale fino a un punto finale vagamente correlato a quell'idea iniziale. L’intuizione e l’elaborazione olistica sono molto importanti in questo emisfero. Comprendere come funziona questo lato del cervello è utile per creare pratiche pedagogiche basate su di esso, per questo l'autore fornisce una descrizione dettagliata e semplice delle attività che si svolgono su questo lato del cervello del bambino e delle strategie che l'insegnante applica per utilizzare questo stesso emisfero.
Matura dopo i sei mesi di vita e la sua maturazione maggiore termina alla pubertà. I talenti che si basano su questo lato del cervello sono: linguistico, logico simbolico matematico e interpersonale o riflessivo. Emisfero sinistro mette il bambino in contatto con la conoscenza concettuale, ma tale apprendimento è fragile e suscettibile di dimenticare. Si può dire che il consolidamento di questi favorisca il sostegno dell'emisfero destro. Come per l'emisfero destro, Amanda elenca le attività che si svolgono in questa parte del cervello del bambino e dell'insegnante.
3 volte la superficie degli emisferi visibile durante l'esame esterno. Ciò è dovuto al fatto che la superficie degli emisferi cerebrali è piegata e presenta numerose depressioni - solchi (sulci cerebrali) e situato tra di loro convoluzioni (gyri cerebri). La corteccia cerebrale copre l'intera superficie delle circonvoluzioni e dei solchi (da qui il suo altro nome pallio - mantello), a volte penetrando a grandi profondità nella sostanza del cervello.
Impara ad alternare i numeri per formare numeri per eseguire operazioni e risolvere problemi. Comprendere, afferma l'autore, la struttura del cervello nel suo insieme e i suoi meccanismi esecutivi significa avere le condizioni per comprendere il bambino o l'adolescente e intervenire in strategie pedagogiche che si adattino alle esigenze di ciascuno studente. Ma quanto sappiamo del cervello e come applichiamo questa conoscenza per creare strumenti che garantiscano non solo l’apprendimento, ma anche la costruzione della vita sociale, scolastica e familiare?
Quali lobi si distinguono negli emisferi cerebrali?
È un dato di fatto che le conversazioni che i genitori hanno con i propri figli sono fondamentali per i processi di apprendimento. Secondo molti esperti, questo li aiuta a sviluppare il linguaggio, le abilità sociali e altre cose vitali per comprendere il mondo che li circonda. A portata di mano con questo ci sono nuove scoperte che evidenziano l’importanza di comunicare con gli obiettivi.
La gravità e la posizione dei solchi e delle circonvoluzioni degli emisferi cerebrali sono variabili in una certa misura, ma i principali si formano durante il processo di ontogenesi e sono costanti, caratteristici di ogni cervello normalmente sviluppato.
14.2. PRINCIPALI SCANALATURE E GIRILI DEGLI EMISFERI DEL CERVELLO
Superficie superolaterale (convessa) degli emisferi (Fig. 14.1a). Il più grande e il più profondo - laterale solco (sulco laterale),o silviano solco, - separa le parti frontale e anteriore del lobo parietale da quello inferiore Lobo temporale. I lobi frontali e parietali sono separati solco centrale o rolandico(solco centrale), che taglia bordo superiore emisfero ed è diretto lungo la sua superficie convessa verso il basso e in avanti, leggermente prima del solco laterale. Il lobo parietale è separato da quello situato dietro di esso Lobo occipitale I solchi parieto-occipitale e occipitale trasverso corrono lungo la superficie mediale dell'emisfero.
Insula, corteccia limbica
Secondo gli esperti, alcuni studi basati sull'osservazione delle famiglie hanno suggerito che parlare di numeri durante le attività e i giochi quotidiani può predire le successive abilità matematiche dei bambini. Inoltre, le prove affermano anche questo apprendimento precoce promuove la comprensione a lungo termine della matematica e della lingua. Ora, anche se i bambini nascono per imparare i numeri, hanno bisogno di sostegno quando lo fanno stiamo parlando sul collegamento dei nomi dei numeri con numero esatto per imparare a rappresentare diversi numeri di oggetti.
Nel lobo frontale, davanti al giro centrale e parallelo ad esso, il giro precentrale (giro precentrale), O anteriore centrale, giro, che è delimitato anteriormente dal solco precentrale (sulco precentrale). I solchi frontali superiore e inferiore si estendono anteriormente dal solco precentrale, dividendo la superficie convessa delle parti anteriori del lobo frontale in tre giri frontali: superiore, medio e inferiore. (giri frontali superiore, media e inferiore).
Supporto per l'apprendimento dei numeri
Quando i genitori hanno queste conversazioni con i loro figli, possono scoprirlo. Questa comprensione del numero di personaggi è davvero importante quando si inizia a lavorare con quantità molto più grandi e complesse processo scolastico. Tuttavia è importante che i bambini conoscano la sequenza numerica, ma i genitori dovrebbero praticarla con i propri figli molto più che essere considerati tali. Se ciò non viene fatto, il bambino avrà difficoltà a capire cosa significano i nomi dei numeri e come usarli. Pertanto, gli esperti offrono alcune idee per sostenerli prima di affrontare la matematica a scuola.
La sezione anteriore della superficie convessa del lobo parietale è costituita dal solco postcentrale situato dietro il solco centrale. (giro postcentrale), o centrale posteriore, giro. È delimitato posteriormente dal solco postcentrale, da cui si estende all'indietro il solco intraparietale. (sulco intraparietale), separano i lobuli parietali superiori ed inferiori (lobuli parietali superiori e inferiori). Nel lobulo parietale inferiore, a sua volta, si distingue il giro sopramarginale (giro sopramarginale), che circonda la parte posteriore della fessura laterale (Silviana) e il giro angolare (girus angolare), confinante con la parte posteriore della circonvoluzione temporale superiore.
Molto più che leggere sequenza numerica, conta e discute un numero di oggetti visibili. Conta e classifica il numero di oggetti in un insieme. Ciò consentirà ai tuoi bambini di connettersi alla routine di conteggio in base alla dimensione impostata.
Utilizza nomi numerici leggermente più grandi di quelli che tuo figlio già comprende. Guarda i gesti con i numeri dei bambini. Molte volte rispondono correttamente con i gesti ma non con le parole. Anche i tuoi gesti sono utili. Quando conti i numeri con i bambini, indica ogni oggetto che fa parte del conteggio. Usa le dita per mostrare l'importo di cui stai parlando.
Sulla superficie convessa del lobo occipitale del cervello, i solchi sono superficiali e possono variare in modo significativo, per cui anche la natura delle convoluzioni situate tra loro è variabile.
La superficie convessa del lobo temporale è divisa dal solco temporale superiore e inferiore, che hanno una direzione quasi parallela alla fessura laterale (silviana), dividendo la superficie convessa del lobo temporale nelle circonvoluzioni temporali superiore, media e inferiore (gyri temporales superior, media et inferior). Il giro temporale superiore forma il labbro inferiore della fessura laterale (silviana). Sulla sua superficie rivolta
Approfitta della quantità Vita di ogni giorno. Esercitati a contare mentre leggi i libri. Chiedi ai bambini di selezionare un certo numero di elementi durante la routine quotidiana. Confronta i numeri durante il gioco. Parla ai tuoi figli di come usi i numeri nella tua vita quotidiana. Ad esempio, analizza come li usi per sapere cosa acquistare, pianificare i pasti o apparecchiare una tavola.
Se sosteniamo il processo di lettura, perché non fare lo stesso con i numeri? In diverse occasioni, il Centro sull’infanzia in via di sviluppo dell’Università di Harvard ha parlato dell’importanza delle interazioni ricettive e premurose tra adulti e bambini. Questo, affermano i ricercatori del centro, è fondamentale per costruire una solida base cerebrale per l'apprendimento e lo sviluppo futuro. Questa comunicazione o interazione è chiamata “servire e restituire” ed è essenziale per costruire le basi dell’apprendimento e del benessere ottimale.
Ai lati del solco laterale sono presenti numerosi solchi trasversali che evidenziano piccole circonvoluzioni trasversali su di esso (convoluzioni di Heschl), che può essere visto solo allargando i bordi della scanalatura laterale.
La parte anteriore della fessura laterale (Silviana) è una depressione dal fondo ampio, che forma la cosiddetta isola (insula), O insula (Lubus insularis). Viene chiamato il bordo superiore del solco laterale che ricopre quest'isola pneumatico (opercolo).
Struttura del lobo temporale
Ora, per praticare questo “gioco” che coinvolge entrambe le parti, ci sono delle azioni o dei passi che si possono compiere, quali sono? Osserva quando interagisci con i bambini. Stai guardando qualcosa di speciale o stai facendo rumore? Hai un'espressione facciale speciale? Muovi le braccia o le gambe? Queste cose, dicono al centro, sono servizi ed è importante tenerli d'occhio durante tutta la giornata situazioni diverse, in quanto consente loro di conoscere le competenze, gli interessi e i bisogni dei bambini. In questo modo li motiverai a esplorare e rafforzerai il legame che hanno.
Superficie interna (mediale) dell'emisfero (Fig. 14.1b). La parte centrale della superficie interna dell'emisfero è strettamente connessa con le strutture del diencefalo, da cui è delimitata da quelle relative al cervello volta (fornice) E corpo calloso (corpo calloso). Quest'ultimo è delimitato esternamente dal solco del corpo calloso (sulcus corporis callosi), iniziando dalla parte anteriore: il becco (rostro) e termina nella sua estremità posteriore ispessita (splenio). Qui il solco del corpo calloso passa nel profondo solco dell'ippocampo (sulcus hippocampi), che penetra in profondità nella sostanza dell'emisfero, premendolo nella cavità del corno inferiore ventricolo laterale, a seguito del quale si forma il cosiddetto corno di ammonio.
Abbracci, parole affettuose, giochi, espressioni facciali e altre azioni possono essere un modo per ricambiare i favori che i bambini offrono. In questo modo viene loro offerto sostegno e sono motivati a continuare a servire, cosa che secondo il centro è una sorta di ricompensa per la curiosità e l'interesse dei bambini. Non ricevere nulla in cambio da un adulto può essere fonte di stress per loro. D’altra parte, quando ricevono una risposta, possono sapere che i loro pensieri e sentimenti, oltre al loro interesse, contano davvero.
Quando si restituisce un servizio, è importante nominare ciò che i bambini vedono, sentono o fanno. In questo modo creerai importanti connessioni linguistiche a livello cerebrale, ancor prima di imparare a capire le parole e a parlare. Ad esempio, se tuo figlio indica i suoi piedi, puoi dire cose come: "Sì, quelli sono i suoi piedi". Perché è importante? Perché in questo modo lo aiuterai a comprendere il mondo che lo circonda e gli farai sapere cosa aspettarsi da lui. Dare un nome alle cose è anche un modo per dare loro parole che possano usare e dimostrare loro che ci tieni.
Leggermente ritirati dal solco del corpo calloso e dal solco dell'ippocampo, si trovano i solchi calloso-marginale, sottoparietale e nasale, che sono una continuazione l'uno dell'altro. Questi solchi delimitano la parte arcuata esterna della superficie mediale dell'emisfero cerebrale, denominata lobo limbico(lobo limbico). Ci sono due giri nel lobo limbico. La parte superiore del lobo limbico è il giro limbico superiore (marginale superiore) o circondante (girus cinguli), la parte inferiore è formata dal giro limbico inferiore, o giro del cavalluccio marino (girus ippocampo), o giro paraippocampale (girus paraippocampalis), davanti al quale c'è un gancio (uncus).
Intorno al lobo limbico del cervello ci sono formazioni della superficie interna del frontale, parietale, occipitale e Lobo temporale sì. La maggior parte della superficie interna del lobo frontale è occupata dal lato mediale del giro frontale superiore. Situato al confine tra i lobi frontali e parietali dell'emisfero cerebrale lobulo paracentrale (lobulis paracentralis), che è una continuazione dei giri centrali anteriore e posteriore sulla superficie mediale dell'emisfero. Al confine tra i lobi parietale e occipitale è chiaramente visibile il solco parieto-occipitale (sulco parietooccipitale). Si estende indietro dalla parte inferiore di esso solco calcarino (solco calcarinus). Tra questi solchi profondi c'è un giro di forma triangolare noto come cuneo. (cuneo). Davanti al cuneo c'è un giro quadrangolare legato al lobo parietale del cervello: il precuneo.
Superficie inferiore dell'emisfero (Fig. 14.1c). La superficie inferiore dell'emisfero cerebrale è costituita da formazioni dei lobi frontale, temporale e occipitale. Adiacente a linea mediana il lobo frontale è formato dal giro retto (giro retto).È delimitato esternamente dal solco olfattivo (sulcus olphactorius), al quale sono adiacenti le formazioni dell'analizzatore olfattivo di seguito: il bulbo olfattivo e il tratto olfattivo. Lateralmente ad essa, fino alla fessura laterale (silviana), che si estende sulla superficie inferiore del lobo frontale, si trovano piccoli giri orbitali (giro orbitale). Le parti laterali della superficie inferiore dell'emisfero dietro il solco laterale sono occupate dal giro temporale inferiore. Medialmente ad esso si trova il giro temporo-occipitale laterale (giro occipitotemporale laterale), o solco fusiforme. Prima-
i suoi dipartimenti con dentro confinano con il giro ippocampale e quelli posteriori con quello linguale (giro linguale) o giro temporo-occipitale mediale (giro occipitotemporalis mediale). Quest'ultimo con la sua estremità posteriore è adiacente al solco calcarino. Le sezioni anteriori del giro fusiforme e linguale appartengono al lobo temporale e le sezioni posteriori appartengono al lobo occipitale del cervello.
14.3. MATERIA BIANCA DEI GRANDI EMISFERI
La sostanza bianca degli emisferi cerebrali è costituita da fibre nervose, prevalentemente mielinica, che costituisce i percorsi che forniscono connessioni tra neuroni corticali e gruppi di neuroni che formano talamo, nodi sottocorticali e nuclei. La parte principale della sostanza bianca degli emisferi cerebrali si trova nella sua profondità centro semiovale o corona radiata (corona radiata), costituito principalmente da afferente ed efferente proiezione vie che collegano la corteccia cerebrale con i gangli sottocorticali, i nuclei e la sostanza reticolare del diencefalo e tronco encefalico, con segmenti midollo spinale. Si trovano in modo particolarmente compatto tra il talamo e i nodi sottocorticali, dove formano la capsula interna descritta nel capitolo 3.
Vengono chiamate le fibre nervose che collegano parti della corteccia di un emisfero associativo. Quanto più corte sono queste fibre e i legami che formano, tanto più superficiali sono localizzate; connessioni associative più lunghe, situate più in profondità, collegano aree relativamente distanti della corteccia cerebrale (Fig. 14.2 e 14.3).
Vengono chiamate fibre che collegano gli emisferi cerebrali e quindi hanno una direzione trasversale comune commissurale, o adesivo. Le fibre commissurali collegano aree identiche degli emisferi cerebrali, creando la possibilità di combinare le loro funzioni. Si formano tre commissure cervello grande: il più massiccio lo è corpo calloso (corpo calloso), inoltre si compongono le fibre commissurali commissura anteriore, situato sotto il becco del corpo calloso (rostro corporeo colloso) e collegando entrambe le aree olfattive, nonché commissura del fornice (commissura fornicis), oppure la commissura ippocampale, formata da fibre che collegano le strutture dei corni ammoniacali di entrambi gli emisferi.
Nella sezione anteriore del corpo calloso ci sono fibre che collegano i lobi frontali, quindi fibre che collegano i lobi parietali e temporali, e la sezione posteriore del corpo calloso collega i lobi occipitali del cervello. La commissura anteriore e la commissura del fornice uniscono principalmente aree dell'antica e vecchia corteccia di entrambi gli emisferi, inoltre, fornisce una connessione tra il loro giro temporale medio e inferiore;
14.4. SISTEMA OLFATTIVO
Nel processo di filogenesi, lo sviluppo di un grande cervello è associato alla formazione del sistema olfattivo, le cui funzioni contribuiscono alla preservazione della vitalità degli animali e non hanno poca importanza per la vita umana.
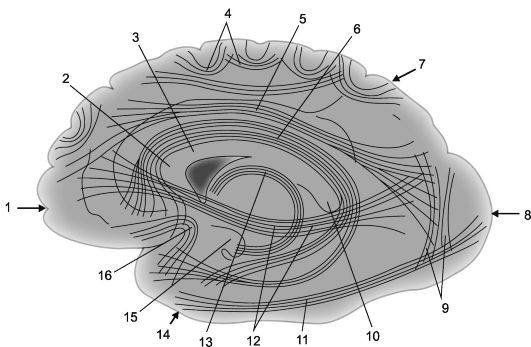
Riso. 14.2.Associazione connessioni cortico-corticali negli emisferi cerebrali [secondo V.P. Vorobyov].
1 - lobo frontale; 2 - genere corpo calloso; 3 - corpo calloso; 4 - fibre arcuate; 5 - trave longitudinale superiore; 6 - giro del cingolo; 7 - lobo parietale, 8 - lobo occipitale; 9 - travi Wernicke verticali; 10 - splenio del corpo calloso;
11 - trave longitudinale inferiore; 12 - fascio subcalloso (fascio inferiore fronto-occipitale); 13 - volta; 14 - lobo temporale; 15 - gancio del giro dell'ippocampo; 16 - ciuffi a uncino (fasciculus uncinatus).
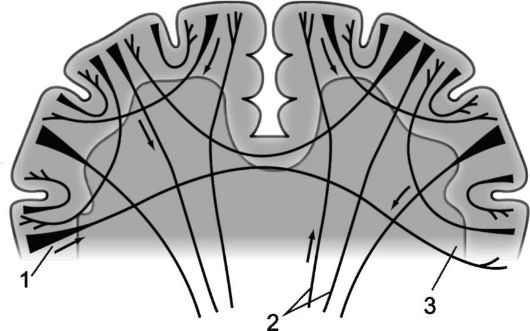
Riso. 14.3.Mieloarchitettura degli emisferi cerebrali.
1 - fibre di proiezione; 2 - fibre commissurali; 3 - fibre associative.
14.4.1. Struttura del sistema olfattivo
I corpi dei primi neuroni del sistema olfattivo si trovano nella mucosa naso, soprattutto la parte superiore del setto nasale e il meato nasale superiore. Le cellule olfattive sono bipolari. I loro dendriti si estendono fino alla superficie della mucosa e terminano qui con recettori specifici, e cluster di assoni nel cosiddetto filamenti olfattivi (fili olfattivi), il cui numero su ciascun lato è di circa venti. Come un fascio di filamenti olfattivi e costituisce il primo nervo cranico, o olfattivo(Fig. 14.4). Questi fili passano nella fossa cranica anteriore (olfattiva, olfattiva) attraverso l'osso etmoidale e terminano in celle situate qui bulbi olfattivi. I bulbi olfattivi e i tratti olfattivi localizzati prossimalmente sono, infatti, una conseguenza delle sporgenze della sostanza cerebrale che si formano durante l'ontogenesi e rappresentano strutture ad essa correlate.
I bulbi olfattivi contengono cellule che sono i corpi dei secondi neuroni via olfattiva, i cui assoni si formano tratti olfattivi (tratti olfattivi), situato sotto i solchi olfattivi, lateralmente alle convoluzioni diritte situate sulla superficie basale lobi frontali. Le vie olfattive sono dirette all'indietro ai centri olfattivi sottocorticali. Avvicinandosi alla placca forata anteriore, le fibre del tratto olfattivo si dividono in fasci mediali e laterali, formando su ciascun lato un triangolo olfattivo. Queste fibre sono successivamente adatte ai corpi dei terzi neuroni dell'analizzatore olfattivo, localizzati
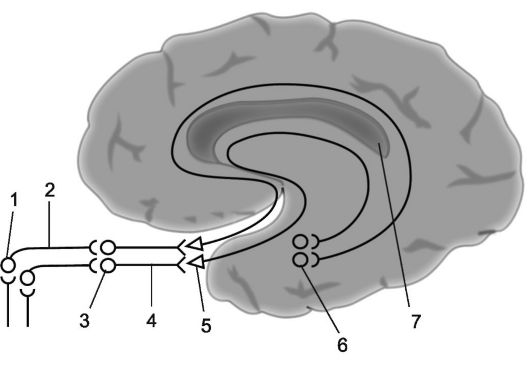
Riso. 14.4.Analizzatore olfattivo.
1 - cellule olfattive; 2 - filamenti olfattivi (insieme costituiscono i nervi olfattivi); 3 - bulbi olfattivi; 4 - tratti olfattivi; 5 - triangoli olfattivi; 6 - giro paraippocampale; 7 - zona di proiezione dell'analizzatore olfattivo (schema semplificato).
nelle aree periamigdala e sottocallosale, nei nuclei del setto pellucido, situato anteriormente alla commissura anteriore. La commissura anteriore collega entrambe le aree olfattive e fornisce anche la loro connessione con il sistema limbico del cervello. Alcuni assoni dei terzi neuroni dell'analizzatore olfattivo, passando attraverso la commissura anteriore del cervello, si incrociano.
Assoni dei terzi neuroni analizzatore olfattivo situato nei centri olfattivi sottocorticali, diretti a corteccia filogeneticamente vecchia la superficie mediobasale del lobo temporale (al giro piriforme e paraippocampale e all'uncus), dove si trova la zona olfattiva di proiezione, o l'estremità corticale dell'analizzatore olfattivo (campo 28, secondo Brodmann).
Il sistema olfattivo è quindi l'unico sistema sensoriale in cui impulsi specifici oltrepassano il talamo nel percorso dai recettori alla corteccia. Allo stesso tempo Il sistema olfattivo ha connessioni particolarmente pronunciate con le strutture limbiche del cervello e le informazioni ricevute attraverso di esso hanno un impatto significativo sullo stato della sfera emotiva e sulle funzioni del sistema nervoso autonomo. Gli odori possono essere piacevoli o sgradevoli, influenzano l'appetito, l'umore e possono causare una serie di reazioni autonomiche, in particolare nausea e vomito.
14.4.2. Studio dell'olfatto e significato dei suoi disturbi per la diagnostica topica
Quando si esamina lo stato dell'olfatto, è necessario scoprire se il paziente percepisce gli odori, se queste sensazioni sono le stesse su entrambi i lati, se il paziente differenzia la natura degli odori percepiti, se ha allucinazioni olfattive - sensazioni parossistiche di un odore che è assente nel ambiente.
Per studiare l'olfatto vengono utilizzate sostanze odorose, il cui odore non è pungente (gli odori pungenti possono causare irritazione dei recettori situati nella mucosa nasale nervo trigemino) ed è nota al paziente (altrimenti è difficile riconoscere la perversione dell'olfatto). L'olfatto viene controllato separatamente su ciascun lato, mentre l'altra narice deve essere chiusa. È possibile utilizzare set appositamente preparati di soluzioni deboli di sostanze odorose (menta, catrame, canfora, ecc.) Possono essere utilizzati anche nel lavoro pratico (); pane di segale, sapone, banana, ecc.).
Diminuzione del senso dell'olfatto - iposmia, mancanza di senso dell'olfatto - anosmia, senso dell'olfatto accentuato - iperosmia, perversione degli odori - disosmia, sensazione dell'olfatto in assenza di stimoli - parosmia, sentimento soggettivo odore sgradevole realmente esistente e causato da patologia organica nel rinofaringe - cacosmia, odori inesistenti che il paziente avverte parossisticamente - allucinazioni olfattive - sono più spesso l'aura olfattiva dell'epilessia del lobo temporale, che può essere causata da vari motivi, in particolare un tumore del lobo temporale.
L'iposmia o l'anosmia su entrambi i lati è solitamente una conseguenza del danno alla mucosa nasale causato da condizioni catarrali acute, influenza, rinite allergica, atrofia della mucosa
naso a causa di rinite cronica e uso a lungo termine gocce vasocostrittrici nel naso. Rinite cronica con atrofia della mucosa nasale (rinite atrofica), la malattia di Sjögren condanna una persona all'anosmia persistente. L'iposmia bilaterale può essere causata da ipotiroidismo, diabete mellito, ipogonadismo, insufficienza renale, contatto prolungato con metalli pesanti, formaldeide, ecc.
Allo stesso tempo l'iposmia o l'anosmia unilaterale è spesso una conseguenza di un tumore intracranico, il più delle volte meningioma della fossa cranica anteriore (olfattiva), che rappresenta fino al 10% dei meningiomi intracranici, nonché alcuni tumori gliali del lobo frontale. I disturbi olfattivi insorgono a causa della compressione del tratto olfattivo dal lato del focolaio patologico e possono essere per un certo periodo l'unico sintomo focale della malattia. La visualizzazione dei tumori può essere fornita mediante scansione TC o MRI. Man mano che i meningiomi della fossa olfattiva si ingrandiscono, di solito si sviluppano disordini mentali, caratteristico della sindrome frontale (vedi Capitolo 15).
Il danno unilaterale alle parti dell'analizzatore olfattivo situate sopra i suoi centri sottocorticali, dovuto all'incrocio incompleto dei percorsi a livello della commissura cerebrale anteriore, di solito non porta ad una diminuzione significativa dell'olfatto. L'irritazione da parte del processo patologico della corteccia delle parti mediobasi del lobo temporale, principalmente il giro paraippocampale e il suo uncus, può causare episodi parossistici allucinazioni olfattive. Il paziente inizia improvvisamente a sentire, senza motivo, un odore, spesso di natura sgradevole (odore di bruciato, di marcio, di marcio, di bruciato, ecc.). Allucinazioni olfattive in presenza di un focolaio epilettogeno nelle regioni mediobasi del lobo temporale del cervello può essere una manifestazione dell'aura di un attacco epilettico. Sconfitta parte prossimale, in particolare l'estremità corticale dell'analizzatore olfattivo, può causare una moderata iposmia bilaterale (più sul lato opposto) e una ridotta capacità di identificare e differenziare gli odori (agnosia olfattiva). Quest'ultima forma di disturbo olfattivo, che si manifesta in età avanzata, è molto probabilmente associata alla disfunzione della corteccia dovuta a processi atrofici nella sua zona di proiezione olfattiva.
14.5. COMPLESSO LIMBICO-RETICOLARE
Nel 1878 P. Broca(Broca P., 1824-1880) chiamato “grande lobo marginale, o limbico” (dal latino limbus - bordo) univa l’ippocampo e il giro del cingolo, collegati tra loro dall'istmo del giro cingolato, situato sopra lo splenio del corpo calloso.
Nel 1937 D. Papets(Papez J.), sulla base di dati sperimentali, ha avanzato un'obiezione motivata al concetto precedentemente esistente del coinvolgimento delle strutture mediobasi degli emisferi cerebrali principalmente nella fornitura dell'olfatto. Lui ha suggerito che la parte principale delle parti mediobasi dell'emisfero cerebrale, allora chiamata cervello olfattivo (rinencefalo), a cui appartiene il lobo limbico, rappresenti la base morfologica meccanismo nervoso comportamento affettivo e combinarli sotto il nome"circolo emotivo" che includeva l'ipotalamo,
nuclei anteriori del talamo, corteccia cingolata, ippocampo e loro connessioni. Da allora queste strutture sono state chiamate anche dai fisiologi intorno a Papetsa.
Concetto "cervello viscerale" suggerito dal P.D. McLean (1949), designando così una complessa associazione anatomo-fisiologica, che dal 1952 cominciò a chiamarsi "sistema limbico". Successivamente si è scoperto che il sistema limbico è coinvolto nell'esecuzione di diverse funzioni, e ora la maggior parte della sua parte, compresi i giri cingolati e ippocampali (paraippocampali), è solitamente combinata nella regione limbica, che ha numerose connessioni con le strutture di la formazione reticolare, ricomponendosi con essa complesso limbico-reticolare, che fornisce una vasta gamma di processi fisiologici e psicologici.
Attualmente a lobo limbico È consuetudine attribuire elementi della vecchia corteccia (archiocorteccia) che ricoprono il giro dentato e il giro ippocampale; antica corteccia (paleocorteccia) dell'ippocampo anteriore; così come la corteccia media o intermedia (mesocorteccia) del giro del cingolo. Termine "sistema limbico" comprende componenti del lobo limbico e strutture correlate - entorinale (occupante maggior parte giro paraippocampale) e la regione del setto, nonché il complesso dell'amigdala e il corpo mastoideo (Duus P., 1995).
Corpo mastoideo collega le strutture di questo sistema con il mesencefalo e con formazione reticolare. Gli impulsi che nascono nel sistema limbico possono essere trasmessi attraverso il nucleo anteriore del talamo al giro del cingolo e alla neocorteccia lungo percorsi formati da fibre associative. Gli impulsi che hanno origine nell'ipotalamo possono raggiungere la corteccia orbitofrontale e il nucleo dorsale mediale del talamo.
Numerose connessioni dirette e di feedback assicurano l'interconnessione e l'interdipendenza delle strutture limbiche e di numerose formazioni del diencefalo e delle parti orali del tronco (nuclei aspecifici del talamo, ipotalamo, putamen, frenulo, formazione reticolare del tronco encefalico), nonché con i nuclei sottocorticali (globus pallidus, putamen, nucleo caudato) e con la nuova corteccia degli emisferi cerebrali, principalmente con la corteccia dei lobi temporali e frontali.
Nonostante le differenze filogenetiche, morfologiche e citoarchitettoniche, molte delle strutture citate (regione limbica, strutture centrali e mediali del talamo, ipotalamo, formazione reticolare del tronco cerebrale) sono solitamente incluse nei cosiddetti complesso limbico-reticolare, che funge da zona di integrazione di molteplici funzioni, garantendo l'organizzazione di reazioni multimodali e olistiche del corpo a varie influenze, che è particolarmente pronunciata in situazioni di stress.
Le strutture del complesso limbico-reticolare hanno un gran numero di ingressi e uscite attraverso i quali transitare cerchi chiusi di numerose connessioni afferenti ed efferenti, garantendo il funzionamento combinato delle formazioni comprese in questo complesso e la loro interazione con tutte le parti del cervello, compresa la corteccia cerebrale.
Nelle strutture del complesso limbico-reticolare c'è una convergenza di impulsi sensibili che sorgono negli intero- ed esterocettori, compresi i campi recettoriali degli organi sensoriali. Su questa base si verifica nel complesso limbico-reticolare sintesi primaria delle informazioni sullo stato dell'ambiente interno del corpo, nonché sui fattori ambientali esterni che influenzano il corpo e si formano bisogni elementari, motivazioni biologiche ed emozioni di accompagnamento.
Il complesso limbico-reticolare determina lo stato della sfera emotiva, partecipa alla regolazione delle relazioni vegetativo-viscerali volte a mantenere la relativa costanza dell'ambiente interno (omeostasi), nonché all'approvvigionamento energetico e alla correlazione degli atti motori. Dal suo stato dipendono il livello di coscienza, la possibilità di movimenti automatizzati, l'attività delle funzioni motorie e mentali, la parola, l'attenzione, la capacità di navigare, la memoria, il cambiamento della veglia e del sonno.
Il danno alle strutture del complesso limbico-reticolare può essere accompagnato da una varietà di sintomi clinici: cambiamenti pronunciati nella sfera emotiva di natura permanente e parossistica, anoressia o bulimia, disturbi sessuali, disturbi della memoria, in particolare segni della sindrome di Korsakoff, in cui il paziente perde la capacità di ricordare eventi attuali (gli eventi attuali vengono conservati in memoria per non più di 2 minuti), disturbi vegetativo-endocrini, disturbi del sonno, disturbi psicosensoriali sotto forma di illusioni e allucinazioni, alterazioni della coscienza, manifestazioni cliniche di mutismo acinetico, crisi epilettiche.
Ad oggi, sono stati condotti numerosi studi per studiare la morfologia, le connessioni anatomiche, la funzione della regione limbica e altre strutture incluse nel complesso limbico-reticolare, tuttavia, è ancora necessaria la fisiologia e le caratteristiche del quadro clinico del suo danno chiarimento oggi. La maggior parte delle informazioni sulla sua funzione lo sono in particolare le funzioni della regione paraippocampale, ottenuto in esperimenti su animali metodi di irritazione, estirpazione o stereotassi. Ottenuto in questo modo i risultati richiedono cautela quando li si estrapolano agli esseri umani. Di particolare importanza sono le osservazioni cliniche di pazienti con lesioni delle regioni mediobasi dell'emisfero cerebrale.
Negli anni 50-60 del XX secolo. Durante lo sviluppo della psicochirurgia, sono apparsi rapporti sul trattamento di pazienti con malattie incurabili disordini mentali e sindrome del dolore cronico attraverso la cingulotomia bilaterale (dissezione del giro cingolato), mentre la regressione dell'ansia, degli stati ossessivi, dell'agitazione psicomotoria, sindromi dolorose, che è stato riconosciuto come prova della partecipazione della corteccia cingolata alla formazione delle emozioni e del dolore. Allo stesso tempo, la bisingulotomia portava a profondi disturbi personali, disorientamento, diminuzione della criticità della propria condizione ed euforia.
Un'analisi di 80 osservazioni cliniche verificate di lesioni dell'ippocampo presso l'Istituto neurochirurgico dell'Accademia russa delle scienze mediche è presentata nella monografia di N.N. Bragina (1974). L'autore giunge alla conclusione che sindrome temporale mediobasale comprende disturbi viscerovegetativi, motori e mentali, solitamente manifestati in un complesso. Tutta la varietà delle manifestazioni cliniche di N.N. Bragin lo riduce a due principali varianti multifattoriali della patologia con una predominanza di fenomeni “irritativi” e “inibitori”.
Il primo di questi include disturbi emotivi, accompagnato da irrequietezza motoria (maggiore eccitabilità, verbosità, pignoleria, sensazione di ansia interna), parossismi di paura, malinconia vitale, vari disturbi viscerovegetativi (cambiamenti del polso, respirazione, disturbi gastrointestinali, aumento della temperatura, aumento della sudorazione, ecc.). Questi pazienti, in un contesto di costante irrequietezza motoria, spesso sperimentavano attacchi di eccitabilità motoria.
Nia. L'EEG di questo gruppo di pazienti era caratterizzato da lievi cambiamenti cerebrali verso l'integrazione (ritmo alfa rapido e acuto, oscillazioni beta diffuse). La stimolazione afferente ripetuta provocava chiare reazioni EEG che, a differenza di quelle normali, non svanivano quando gli stimoli venivano presentati ripetutamente.
La seconda versione ("inibitoria") della sindrome mediobasale è caratterizzata da disturbi emotivi sotto forma di depressione con ritardo motorio (umore di fondo represso, impoverimento e rallentamento del ritmo processo mentale, cambiamenti nella motilità, che ricordano la sindrome acinetico-rigida. I parossismi viscerovegetativi riscontrati nel primo gruppo sono meno tipici. L'EEG dei pazienti di questo gruppo era caratterizzato da cambiamenti cerebrali generali, manifestati nella predominanza di forme lente di attività (ritmo alfa irregolare e lento, gruppi di oscillazioni theta, onde delta diffuse). Degna di nota è stata una forte diminuzione della reattività EEG.
Tra queste due opzioni estreme c'erano anche quelle intermedie con combinazioni transitorie e miste sintomi individuali. Pertanto, alcuni di essi sono caratterizzati da segni relativamente deboli di depressione agitata con aumento attività fisica e affaticamento, con predominanza di sensazioni senestopatiche, sospetto, che arriva in alcuni pazienti a stati paranoidi, delirio ipocondriaco. Un altro gruppo intermedio si distingueva per l'estrema intensità dei sintomi depressivi sullo sfondo della rigidità del paziente.
Questi dati ci permettono di parlare della duplice influenza (attivante e inibitoria) dell'ippocampo e di altre strutture della regione limbica sulle reazioni comportamentali, sulle emozioni, sulle caratteristiche dello stato mentale e sull'attività bioelettrica della corteccia. Attualmente difficile sindromi cliniche questo tipo non dovrebbe essere considerato focale primario. Piuttosto, devono essere considerati alla luce delle idee su un sistema multilivello di organizzazione dell'attività cerebrale.
S.B. Buklina (1997) ha fornito i dati di un esame di 41 pazienti con malformazioni artero-venose nell'area del giro del cingolo. Prima dell'operazione, in 38 pazienti, sono emersi disturbi della memoria, e in cinque di loro c'erano segni della sindrome di Korsakov, in tre pazienti, la sindrome di Korsakov è comparsa dopo l'operazione, mentre la gravità dell'aumento dei difetti di memoria era correlata con la sindrome di Korsakov; grado di distruzione del giro cingolato stesso, nonché con il coinvolgimento del processo patologico delle strutture adiacenti del corpo calloso, mentre la sindrome amnesica non dipendeva dal lato della malformazione e dalla sua localizzazione lungo il lungo giro cingolato.
Le caratteristiche principali delle sindromi amnestiche identificate erano disturbi della riproduzione degli stimoli uditivo-verbali, ridotta selettività delle tracce sotto forma di inclusioni e contaminazioni e incapacità di trattenere il significato durante la trasmissione di una storia. La maggior parte dei pazienti presentava una ridotta criticità nella valutazione della propria condizione. L'autore ha notato la somiglianza di questi disturbi con i difetti amnestici nei pazienti con lesioni frontali, che può essere spiegato dalla presenza di connessioni tra il giro del cingolo e il lobo frontale.
Di più processi patologici comuni nella regione limbica causano gravi disturbi delle funzioni autonomo-viscerali.
corpo calloso(corpo calloso)- la più grande commessura tra gli emisferi cerebrali. Le sue sezioni anteriori, in particolare il ginocchio del callo
corpo (genere corporis callosi), collega i lobi frontali, le sezioni centrali - il tronco del corpo calloso (tronco del corpo callosi)- fornire collegamento tra le sezioni temporale e parietale degli emisferi, le sezioni posteriori, in particolare lo splenio del corpo calloso (splenio del corpo calloso), collegano i lobi occipitali.
Le lesioni del corpo calloso sono solitamente accompagnate da disturbi mentali del paziente. La distruzione della sua sezione anteriore porta allo sviluppo della “psiche frontale” (spontaneità, violazioni del piano d'azione, comportamento, critica, caratteristica di sindrome callosa frontale - acinesia, amimia, aspontaneità, astasia-abasia, aprassia, cogliere i riflessi, demenza). La separazione delle connessioni tra i lobi parietali porta alla perversione comprensione "diagrammi del corpo" E la comparsa di aprassia principalmente nella mano sinistra. Dissociazione dei lobi temporali potrebbe apparire disturbo della percezione dell'ambiente esterno, perdita del corretto orientamento in esso (disturbi amnesici, confabulazioni, sindrome del già visto) e così via.). I focolai patologici nelle parti posteriori del corpo calloso sono spesso caratterizzati da segni di agnosia visiva.
14.6. ARCHITETTURA DELLA CORTECCIA CEREBRALE
La struttura della corteccia cerebrale è eterogenea. Meno complessi nella struttura che si presentano all'inizio del processo di filogenesi corteccia antica (archiocorteccia) e vecchia corteccia (paleocorteccia), imparentato soprattutto al lobo limbico cervello La maggior parte della corteccia cerebrale (95,6%) a causa della sua successiva formazione dal punto di vista filogenico è chiamata nuova corteccia (neocorteccia) ed ha una struttura multistrato molto più complessa, ma anche eterogenea nelle sue diverse zone.
A causa di l'architettura della corteccia è in una certa connessione con la sua funzione, Molta attenzione è stata dedicata al suo studio. Uno dei fondatori della dottrina della citoarchitettura della corteccia fu V.A. Betz (1834-1894), che per primo descrisse le grandi cellule piramidali nel 1874 zona motoria corteccia (cellule di Betz) e determinò i principi di divisione della corteccia cerebrale in aree principali. Successivamente, molti ricercatori hanno dato un grande contributo allo sviluppo della dottrina della struttura della corteccia - A. Cambell, E. Smith, K. Brodmann, Oskar Vogt e Cecilia Vogt , S. Vogt). Grandi risultati nello studio dell'architettura corticale appartengono al team del Brain Institute dell'Accademia delle scienze mediche (S.A. Sarkisov, N.I. Filimonov, E.P. Kononova, ecc.).
Il principale tipo di struttura della nuova corteccia (Fig. 14.5), con cui si confrontano tutte le sue sezioni è la corteccia, costituita da 6 strati (corteccia omotipica, secondo Brodmann).
Lo strato I è molecolare, o zonale, il più superficiale, povero di cellule, le sue fibre hanno direzione prevalentemente parallela alla superficie della corteccia.
Strato II - granulare esterno. È costituito da un gran numero di piccoli granulari densamente localizzati cellule nervose.
Strato III: piramidi piccole e medie, le più larghe. È costituito da cellule piramidali, le cui dimensioni non sono uguali, il che consente nella maggior parte dei campi corticali di dividere questo strato in sottostrati.
Strato IV - granulare interno. È costituito da piccole cellule granulari densamente localizzate di forma rotonda e angolare. Questo strato è il più variabile, in
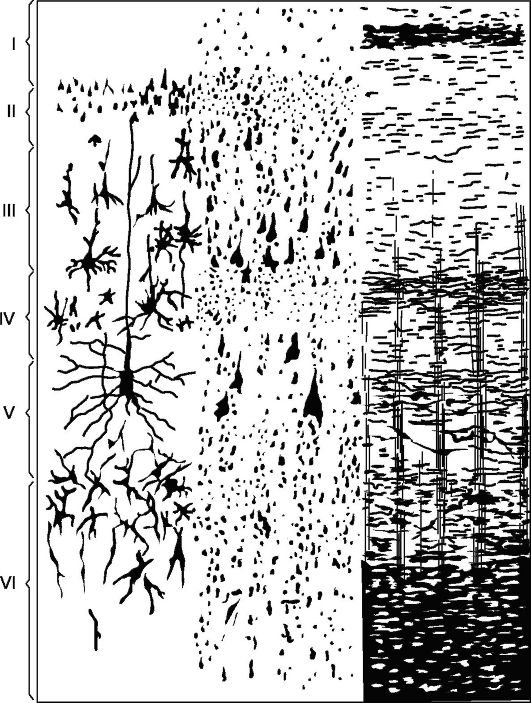
Riso. 14.5.Citoarchitettura e mieloarchitettura della zona motoria della corteccia cerebrale.
A sinistra: I - strato molecolare; II - strato granulare esterno; III - strato di piramidi piccole e medie; IV - strato granulare interno; V - strato di grandi piramidi; VI - strato di cellule polimorfiche; a destra - elementi di mieloarchitettura.
In alcuni campi (ad esempio il campo 17) è diviso in sottostrati, in alcuni punti si assottiglia drasticamente fino a scomparire completamente.
Strato V: grandi piramidi o gangli. Contiene grandi cellule piramidali. In alcune aree del cervello, lo strato è diviso in sottostrati; nella zona motoria è costituito da tre sottostrati, il centro dei quali contiene cellule piramidali giganti di Betz, che raggiungono un diametro di 120 micron.
Strato VI: cellule polimorfiche o multiformi. È costituito principalmente da cellule triangolari a forma di fuso.
La struttura della corteccia cerebrale presenta un gran numero di variazioni dovute a cambiamenti nello spessore dei singoli strati, assottigliamento o scomparsa, o
al contrario, ispessendo e dividendo in sottostrati alcuni di essi (zone eterotipiche, secondo Brodmann).
La corteccia di ciascun emisfero cerebrale è divisa in diverse regioni: occipitale, parietale superiore e inferiore, postcentrale, giro centrale, precentrale, frontale, temporale, limbica, insulare. Ognuno di loro secondo le caratteristiche è suddiviso in una serie di campi, Inoltre, ogni campo ha la propria designazione ordinale convenzionale (Fig. 14.6).
Lo studio dell'architettura della corteccia cerebrale, insieme a studi fisiologici, compresi studi elettrofisiologici e osservazioni cliniche, ha ampiamente contribuito a risolvere il problema della distribuzione delle funzioni nella corteccia.
14.7. CAMPI DI PROIEZIONE E ASSOCIAZIONE DEL CORTAL
Nel processo di sviluppo della dottrina del ruolo della corteccia cerebrale e delle sue singole sezioni nell'esecuzione di determinate funzioni, c'erano punti di vista diversi, a volte opposti. Pertanto, c'era un'opinione sulla rappresentazione strettamente locale nella corteccia cerebrale di tutte le abilità e funzioni umane, fino a quelle mentali più complesse (localizzazionismo, psicomorfologismo). Ad essa si oppose un'altra opinione sull'assoluta equivalenza funzionale di tutte le aree della corteccia cerebrale (equipotenzialismo).
Un importante contributo allo studio della localizzazione delle funzioni nella corteccia cerebrale è stato dato da I.P. Pavlov (1848-1936). Ha identificato le zone di proiezione della corteccia (estremità corticali degli analizzatori singole specie sensibilità) e le zone associative situate tra loro, hanno studiato i processi di inibizione ed eccitazione nel cervello, la loro influenza su stato funzionale corteccia cerebrale. La divisione della corteccia in zone di proiezione e associative contribuisce alla comprensione dell'organizzazione del lavoro della corteccia cerebrale ed è giustificata nella risoluzione problemi pratici, in particolare per la diagnostica topica.
Zone di proiezione fornire principalmente semplici atti fisiologici specifici, principalmente la percezione di sensazioni di una certa modalità. Le vie di proiezione che si avvicinano ad esse collegano queste zone con i territori recettoriali nella periferia che sono in corrispondenza funzionale con esse. Esempi di zone corticali di proiezione sono l'area del giro centrale posteriore (zona tipi comuni sensibilità) o l'area del solco calcarino situata sul lato mediale del lobo occipitale (area visiva proiettiva).
Zone associative la corteccia non ha collegamenti diretti con la periferia. Sono nel mezzo zone di proiezione e hanno numerose connessioni associative con queste proiezioni e altre zone associative. La funzione delle zone associative è quella di implementare analisi più elevate e la sintesi di molti componenti elementari e più complessi. Qui, essenzialmente, si comprendono le informazioni che entrano nel cervello e si formano idee e concetti.
GI Polyakov nel 1969, sulla base di un confronto tra l'architettura della corteccia cerebrale dell'uomo e di alcuni animali, stabilì che l'attività associativa
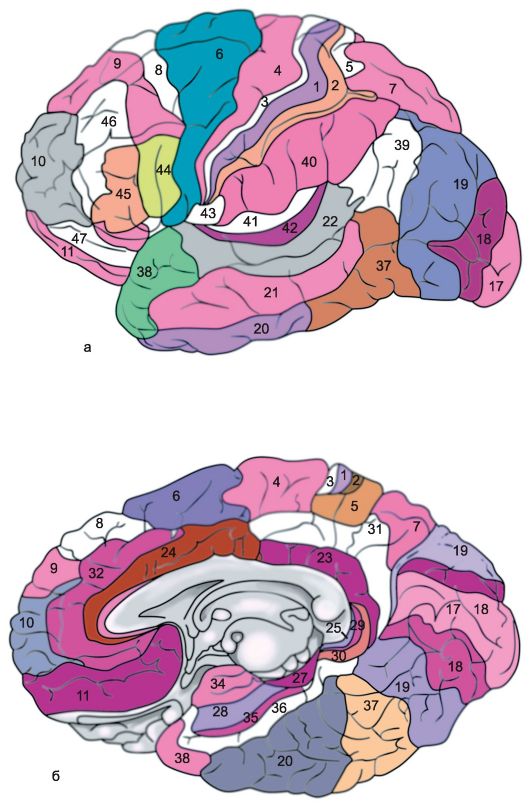
Riso. 14.6.Campi architettonici della corteccia cerebrale [secondo Brodmann]. a - superficie esterna; b - superficie mediale.
le zone nella corteccia cerebrale umana costituiscono il 50%, nella corteccia delle scimmie superiori (antropoidi) - il 20%, nelle scimmie inferiori la stessa cifra è del 10% (Fig. 14.7). Tra le zone associative della corteccia cervello umano, lo stesso autore ha proposto di isolare campi secondari e terziari. I campi associativi secondari sono adiacenti ai campi di proiezione. Effettuano un'analisi e una sintesi di sensazioni elementari che conservano comunque un focus specifico.
Campi associativi terziari si trovano prevalentemente tra quelli secondari e sono zone di sovrapposizione di territori limitrofi. Sono legati principalmente all'attività analitica della corteccia, fornendo funzioni mentali superiori caratteristiche degli esseri umani nelle loro manifestazioni intellettuali e linguistiche più complesse. Maturità funzionale delle attività terziarie
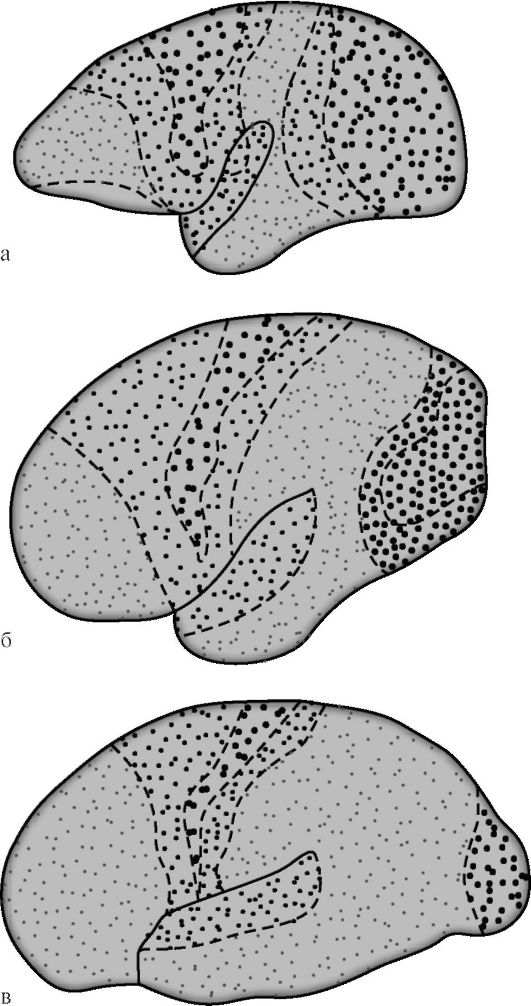
Riso. 14.7. Differenziazione delle zone di proiezione e di associazione della corteccia cerebrale durante l'evoluzione dei primati [secondo G.I. Poliakov]. a - il cervello della scimmia inferiore; b - cervello della grande scimmia; c - cervello umano. I punti grandi indicano le zone di proiezione, i punti piccoli indicano le zone associative. Nelle scimmie inferiori, le zone di associazione occupano il 10% dell'area della corteccia, nelle scimmie superiori - il 20%, negli esseri umani - il 50%.
campi sociali della corteccia cerebrale si verifica più tardi e solo in favorevole contesto sociale. A differenza di altri campi corticali, i campi terziari degli emisferi destro e sinistro sono caratterizzati da un pronunciato asimmetria funzionale.
14.8. DIAGNOSTICA TOPICA DELLE LESIONI DELLA CORTECCIA CEREBRALE
14.8.1. Manifestazioni di danno alle zone di proiezione della corteccia cerebrale
Nella corteccia di ciascun emisfero cerebrale, dietro il giro centrale, ci sono 6 zone di proiezione.
1. Nella parte anteriore del lobo parietale, nella regione del giro centrale posteriore (campi citoarchitettonici 1, 2, 3) situato zona di proiezione di tipi generali di sensibilità(Fig. 14.4). Le aree della corteccia qui situate ricevono impulsi sensibili provenienti lungo le vie di proiezione di tipi generali di sensibilità dall'apparato recettore della metà opposta del corpo. Quanto più alta è la sezione di questa zona di proiezione della corteccia, tanto più basse sono le parti della metà opposta del corpo che hanno connessioni di proiezione. Parti del corpo che hanno una ricezione estesa (lingua, superficie palmare mani), corrispondono a parti insufficientemente grandi dell'area delle zone di proiezione, mentre altre parti del corpo (arti prossimali, tronco) hanno una piccola area di rappresentazione corticale.
L'irritazione da parte di un processo patologico della zona corticale di tipi generali di sensibilità porta ad un attacco di parestesia in parti del corpo corrispondenti alle aree irritate della corteccia cerebrale (crisi jacksoniana sensibile), che può svilupparsi in un parossismo generalizzato secondario. Il danno all'estremità corticale dell'analizzatore di tipi generali di sensibilità può causare lo sviluppo di ipoestesia o anestesia nell'area corrispondente della metà opposta del corpo, mentre l'area di ipoestesia o anestesia può essere di tipo circolatorio verticale o di tipo segmentale radicolare. Nel primo caso, il disturbo della sensibilità si manifesta sul lato opposto al focolaio patologico nella zona delle labbra, del pollice o nella parte distale dell'arto con un bordo circolare, a volte come un calzino o un guanto. Nel secondo caso, la zona di sensibilità ridotta ha la forma di una striscia e si trova lungo il bordo interno o esterno del braccio o della gamba; ciò è spiegato dal fatto che il lato interno degli arti è rappresentato nella parte anteriore e il lato esterno nelle sezioni posteriori della zona di proiezione dell'analizzatore di tipi generali di sensibilità.
2. Zona di proiezione visiva situato nella corteccia della superficie mediale del lobo occipitale nell'area del solco calcarino (campo 17). In questo campo, il IV strato (granulare interno) della corteccia è separato in due sottostrati da un fascio di fibre mieliniche. Sezioni separate del campo 17 ricevono impulsi da certe sezioni delle metà omonime delle retine di entrambi gli occhi; in questo caso giungono impulsi provenienti dalle parti inferiori delle omonime metà della retina
corteccia del labbro inferiore del solco calcarino e impulsi provenienti da parti superiori retina, sono diretti alla corteccia del labbro superiore.
Il danno alla zona di proiezione visiva da parte del processo patologico porta alla comparsa di un quadrante o di un'emianopsia omonima completa sul lato opposto del fuoco patologico. Danno bilaterale ai campi corticali 17 o alle aree di proiezione che conducono ad essi percorsi visivi può portare alla completa cecità. L'irritazione della corteccia della zona di proiezione visiva può causare la comparsa di allucinazioni visive sotto forma di fotopsia nelle parti corrispondenti delle metà opposte dei campi visivi.
3. Zona di proiezione uditiva situato nella corteccia del giro di Heschl labbro inferiore fessura laterale (silviana) (aree 41 e 42), che fanno, infatti, parte della circonvoluzione temporale superiore. L'irritazione di questa zona della corteccia può causare allucinazioni uditive (attacchi di sensazioni di rumore, squillo, fischio, ronzio, ecc.). La distruzione della zona di proiezione uditiva da un lato può causare una leggera diminuzione dell'udito in entrambe le orecchie, soprattutto in quella opposta al focus patologico.
4 e 5. Zone di proiezione olfattiva e gustativa Sono sulla superficie mediale del giro a volta (area limbica) del cervello. Il primo di essi si trova nel giro paraippocampale (campo 28). La zona di proiezione del gusto è solitamente localizzata nella corteccia opercolare (area 43). L'irritazione delle zone di proiezione dell'olfatto e del gusto può provocarne la perversione o portare allo sviluppo di corrispondenti allucinazioni olfattive e gustative. La perdita unilaterale della funzione delle zone di proiezione dell'olfatto e del gusto può causare una leggera diminuzione dell'olfatto e del gusto rispettivamente su entrambi i lati. La distruzione bilaterale delle estremità corticali degli stessi analizzatori si manifesta rispettivamente con l'assenza di odore e gusto su entrambi i lati.
6. Zona di proiezione vestibolare. La sua localizzazione non è stata specificata. Allo stesso tempo, è noto che l'apparato vestibolare presenta numerose connessioni anatomiche e funzionali. È possibile che la localizzazione della rappresentazione del sistema vestibolare nella corteccia non sia stata ancora chiarita perché è polifocale. N.S. Blagoveshchenskaya (1981) ritiene che nella corteccia cerebrale le zone di proiezione vestibolare siano rappresentate da diversi complessi anatomico-funzionali interagenti tra loro, che si trovano nel campo 8, alla giunzione dei fronti frontale, temporale e Lobo parietale e nella zona delle circonvoluzioni centrali si presume che ciascuna di queste aree della corteccia svolga le proprie funzioni. Il campo 8 è un centro dello sguardo arbitrario, la sua irritazione provoca una rotazione dello sguardo nella direzione opposta al focus patologico, cambiamenti nel ritmo e nella natura del nistagmo sperimentale, soprattutto subito dopo un attacco epilettico. Nella corteccia del lobo temporale sono presenti strutture la cui irritazione provoca vertigini, che si manifesta, in particolare, nell'epilessia del lobo temporale; il danno alle aree di rappresentazione delle strutture vestibolari nella corteccia del giro centrale influenza lo stato del tono dei muscoli striati. Osservazioni cliniche suggeriscono che le vie vestibolari nucleo-corticali subiscono una parziale decussazione.
Va sottolineato che i segni di irritazione delle zone di proiezione elencate possono essere una manifestazione dell'aura di un attacco epilettico corrispondente per natura.
IP Pavlov riteneva possibile considerare come zona di proiezione la corteccia del giro precentrale, che influenza le funzioni motorie e il tono muscolare prevalentemente della metà opposta del corpo, con la quale è collegata principalmente dalle vie corticonucleare e corticospinale (piramidale). il cosidetto analizzatore motorio. Questa zona occupa Innanzitutto il campo 4, sul quale la metà opposta del corpo viene proiettata prevalentemente in forma invertita. Questo campo contiene la maggior parte delle cellule piramidali giganti (cellule di Betz), i cui assoni costituiscono il 2-2,5% di tutte le fibre del tratto piramidale, nonché cellule piramidali medie e piccole, che, insieme agli assoni dello stesso le cellule situate nell'area adiacente al campo 4 e al campo 6 più esteso, partecipano alla realizzazione delle connessioni cortico-muscolari monosinaptiche e polisinaptiche. Le connessioni monosinaptiche forniscono principalmente azioni dirette all'obiettivo veloci e precise, a seconda delle contrazioni dell'individuo striato muscoli.
Il danno all'area motoria inferiore di solito porta allo sviluppo sul lato opposto brachiofacciale (omerofacciale) sindrome o sindrome linguo-facciale-brachiale, che si osservano spesso in pazienti con disturbi circolazione cerebrale nel bacino dell'arteria cerebrale media, si evidenzia una paresi combinata dei muscoli del viso, della lingua e del braccio, principalmente della spalla di tipo centrale.
L'irritazione della corteccia della zona motoria (campi 4 e 6) porta alla comparsa di spasmi nei muscoli o nei gruppi muscolari proiettati in questa zona. Più spesso si tratta di crisi locali di tipo epilettico jacksoniano, che possono trasformarsi in una crisi epilettica generalizzata secondaria.
14.8.2. Manifestazioni di danno ai campi associativi della corteccia cerebrale
Tra le zone di proiezione della corteccia ci sono campi associativi. Ricevono impulsi principalmente dalle cellule delle zone di proiezione della corteccia. Nei campi associativi, l'analisi e la sintesi delle informazioni a cui è stata sottoposta lavorazione primaria nei campi di proiezione. Le zone associative della corteccia del lobulo parietale superiore forniscono la sintesi delle sensazioni elementari, quindi qui si formano tipi complessi di sensibilità, come il senso di localizzazione, il senso di peso, il senso spaziale bidimensionale, nonché il senso di localizzazione; sensazioni cinestetiche complesse.
Nell'area del solco interparietale esiste una zona associativa che fornisce la sintesi delle sensazioni emanate da parti del proprio corpo. Il danno a quest'area della corteccia porta a autotopagnosia, quelli. al misconoscimento o all'ignoranza di parti del proprio corpo, o a pseudomelia - la sensazione di avere un braccio o una gamba in più, nonché anosognosia - mancanza di consapevolezza del difetto fisico insorto in relazione alla malattia (ad esempio paralisi o paresi di un arto). Tipicamente, tutti i tipi di autotopagnosia e anosognosia si verificano quando il processo patologico si trova a destra.
Il danno al lobulo parietale inferiore può manifestarsi come un disturbo nella sintesi delle sensazioni elementari o nell'incapacità di confrontare le sensazioni complesse sintetizzate con ciò che una volta era simile nella percezione.
allo stesso modo, sulla base dei risultati dei quali avviene il riconoscimento” (V.M. Bekhterev). Ciò si manifesta con una violazione del senso spaziale bidimensionale (grafoestesia) e del senso spaziale tridimensionale (stereognosi) - astereognosi.
In caso di danno alle zone premotorie del lobo frontale (campi 6, 8, 44), di solito si verifica atassia frontale, in cui la sintesi di impulsi afferenti (afferenza cinestetica), segnalando il cambiamento di posizione delle parti del corpo nello spazio durante i movimenti , è interrotto.
Quando la funzione della corteccia delle parti anteriori del lobo frontale, che ha connessioni con l'emisfero opposto del cervelletto (connessioni frontopontino-cerebellari), è compromessa, si verificano disturbi statocinetici sul lato opposto al focus patologico (atassia frontale). Le violazioni delle forme di statocinetica a sviluppo tardivo - stazione eretta e camminata eretta - sono particolarmente evidenti. Di conseguenza, il paziente sperimenta incertezza e andatura instabile. Mentre si cammina, il corpo si appoggia all'indietro (sintomo di Henner) mette i piedi in linea retta (passeggiata della volpe) a volte quando si cammina si verifica un “intreccio” delle gambe. Alcuni pazienti con danni alle parti anteriori dei lobi frontali sviluppano un fenomeno peculiare: in assenza di paralisi e paresi e della capacità di muovere completamente le gambe, i pazienti non riescono a stare in piedi (astasia) e camminare (Abasia).
Il danno alle zone associative della corteccia è spesso caratterizzato dallo sviluppo di manifestazioni cliniche di disturbi delle funzioni mentali superiori (vedi Capitolo 15).
Panoramica generale della struttura degli emisferi cerebrali
Gli emisferi cerebrali sono la parte più massiccia del cervello. Coprono il cervelletto e il tronco encefalico. Gli emisferi cerebrali costituiscono circa il 78% della massa cerebrale totale. Durante lo sviluppo ontogenetico dell'organismo, gli emisferi cerebrali si sviluppano dalla vescicola cerebrale del tubo neurale, pertanto questa parte del cervello è chiamata anche telencefalo.
Gli emisferi cerebrali sono divisi lungo la linea mediana da una profonda fessura verticale negli emisferi destro e sinistro.
Nella parte centrale, entrambi gli emisferi sono collegati tra loro da una grande commessura: il corpo calloso. Ogni emisfero ha lobi; frontale, parietale, temporale, occipitale e insulare.
I lobi degli emisferi cerebrali sono separati tra loro da profondi solchi. I più importanti sono tre solchi profondi: il centrale (Rolandiano) che separa il lobo frontale dal parietale, il laterale (Silviano) che separa il lobo temporale dal parietale, il parieto-occipitale che separa il lobo parietale dall'occipitale sulla superficie interna del l'emisfero.
Ogni emisfero ha una superficie superolaterale (convessa), inferiore e interna.
Ogni lobo dell'emisfero ha circonvoluzioni cerebrali separate l'una dall'altra da solchi. La parte superiore dell'emisfero è ricoperta da una corteccia, un sottile strato di materia grigia, costituito da cellule nervose.
La corteccia cerebrale è la formazione più giovane del sistema nervoso centrale in termini evolutivi. Nell'uomo raggiunge il suo massimo sviluppo. La corteccia cerebrale è di grande importanza nella regolazione delle funzioni vitali del corpo, nell’attuazione di forme di comportamento complesse e nello sviluppo delle funzioni neuropsichiche.
Sotto la corteccia si trova la sostanza bianca degli emisferi, costituita da processi di cellule nervose - conduttori. A causa della formazione delle circonvoluzioni cerebrali, la superficie totale della corteccia cerebrale aumenta in modo significativo. area totale La corteccia degli emisferi è grande 1200 cm 2, con 2/3 della sua superficie situata nella profondità dei solchi e 1/3 sulla superficie visibile degli emisferi. Ogni lobo del cervello ha un significato funzionale diverso.
a – superficie superolaterale: 1 – giro frontale inferiore; 2 – giro frontale medio; 3 – giro frontale superiore; 4 – giro centrale anteriore; 5 – solco centrale (rolandico); 6 – giro centrale posteriore; 7 – lobulo parietale superiore; 8 – lobulo parietale inferiore; 9 – solco sopramarginale; 10 – scanalatura angolare (angolare); 11 – solco parieto-occipitale; 12 – giro temporale inferiore;
13 – giro temporale medio; 14 – giro temporale superiore; 15 – fessura laterale (silviana);
b – superficie interna: 1 lobulo paracentrale; 2 – scanalatura centrale; 3 – giro del cingolo: 4 – corpo calloso; 5 – solco parieto-occipitale; 6 – cuneo; 7 – solco calcarino; 8 – giro linguale; 9 – giro ippocampale (giro paraippocampale).
Struttura del lobo frontale
Il lobo frontale occupa le parti anteriori degli emisferi. È separato dal lobo parietale dal solco centrale e dal lobo temporale dal solco laterale. Il lobo frontale ha quattro giri: uno verticale - il giro precentrale e tre orizzontali - i giri frontali superiore, medio e inferiore. Le circonvoluzioni sono separate l'una dall'altra da scanalature.
Sulla superficie inferiore dei lobi frontali si distinguono il retto e il giro orbitale. Il giro retto si trova tra il bordo interno dell'emisfero, il solco olfattivo e il bordo esterno dell'emisfero.
Nelle profondità del solco olfattivo si trovano il bulbo olfattivo e il tratto olfattivo.
Il lobo frontale umano costituisce il 25-28% della corteccia; il peso medio del lobo frontale è di 450 g.
Funzione dei lobi frontaliè associato all'organizzazione dei movimenti volontari, ai meccanismi motori della parola, alla regolazione di forme complesse di comportamento e ai processi di pensiero. Diversi centri funzionalmente importanti sono concentrati nelle circonvoluzioni del lobo frontale. Il giro centrale anteriore è una "rappresentazione" della zona motoria primaria con una proiezione rigorosamente definita delle parti del corpo. Il viso è “situato” nel terzo inferiore del giro, la mano è nel terzo medio, la gamba è in terzo superiore. Il tronco è rappresentato nelle parti posteriori del giro frontale superiore. Pertanto, una persona viene proiettata nel giro centrale anteriore a testa in giù e a testa in giù.
Il giro centrale anteriore, insieme al giro posteriore adiacente e alle parti del giro frontale, svolge un ruolo funzionale molto importante. È il centro dei movimenti volontari. Nel profondo della corteccia del giro centrale, la via motoria principale inizia dalle cosiddette cellule piramidali - il motoneurone centrale - piramidale, corticospinale, sentiero. I processi periferici dei motoneuroni lasciano la corteccia, si riuniscono in un unico potente fascio, attraversano la sostanza bianca centrale degli emisferi ed entrano nel tronco cerebrale attraverso la capsula interna; all'estremità del tronco encefalico decussano parzialmente (passando da un lato all'altro) e poi scendono nel midollo spinale. Questi processi finiscono con materia grigia midollo spinale. Lì entrano in contatto con il motoneurone periferico e gli trasmettono gli impulsi dal motoneurone centrale. Di percorso piramidale vengono trasmessi gli impulsi di movimento volontario.
C'è anche nelle parti posteriori del giro frontale superiore extrapiramidale centro della corteccia, strettamente connesso anatomicamente e funzionalmente con le formazioni del cosiddetto sistema extrapiramidale. Sistema extrapiramidale – sistema di propulsione, aiutando a realizzare il movimento volontario. Questo è un sistema per “fornire” movimenti volontari. Essendo filogeneticamente più antico, il sistema extrapiramidale nell’uomo provvede alla regolazione automatica degli atti motori “appresi”, mantenendo il generale tono muscolare, disponibilità periferica sistema muscoloscheletrico per eseguire movimenti, ridistribuzione del tono muscolare durante i movimenti. Inoltre, è coinvolto nel mantenimento della postura normale.
Aree della corteccia motoria si trovano principalmente nel giro precentrale (aree 4 e 6) e nel lobulo paracentrale sulla superficie mediale dell'emisfero. Esistono aree primarie e secondarie: i campi 4 e 6. Questi campi sono motori, ma in base alle loro caratteristiche, secondo una ricerca del Brain Institute, sono diversi. Nella corteccia motoria primaria(campo 4) sono presenti neuroni che innervano i motoneuroni dei muscoli del viso, del tronco e degli arti.
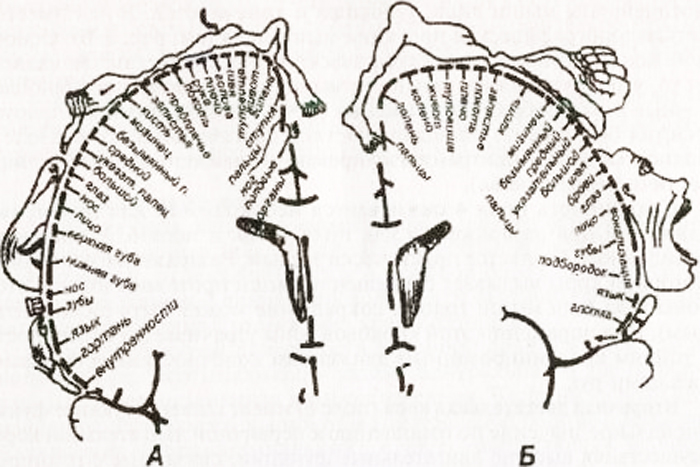
Schema di proiezione autotopica della sensibilità generale e delle funzioni motorie nella corteccia cerebrale (secondo W. Penfield):
A – proiezione corticale della sensibilità generale; B – proiezione corticale del sistema motorio. Le dimensioni relative degli organi riflettono l'area della corteccia cerebrale da cui possono essere provocate le sensazioni e i movimenti corrispondenti
Ha una chiara proiezione topografica dei muscoli del corpo. Il modello principale di rappresentazione topografica è che la regolazione dell'attività dei muscoli che forniscono i movimenti più accurati e vari (parola, scrittura, espressioni facciali) richiede la partecipazione di ampie aree della corteccia motoria. Il campo 4 è completamente occupato da centri di movimenti isolati, il campo 6 è occupato solo parzialmente (sottocampo 6a).
La preservazione del campo 4 risulta necessaria per ottenere movimenti quando vengono stimolati sia il campo 4 che il campo 6. Nel neonato il campo 4 è quasi maturo. La stimolazione della corteccia motoria primaria provoca la contrazione muscolare lato opposto corpo (per i muscoli della testa la contrazione può essere bilaterale). Quando questa zona corticale viene danneggiata, si perde la capacità di compiere movimenti fini e coordinati degli arti e soprattutto delle dita.
Corteccia motoria secondaria(campo 6) ha un significato funzionale dominante in relazione alla corteccia motoria primaria, svolgendo funzioni motorie superiori associate alla pianificazione e alla coordinazione dei movimenti volontari. Qui si registra in massima misura il negativo in lento aumento. potenziale di prontezza, che si verificano circa 1 s prima dell'inizio del movimento. La corteccia dell'area 6 riceve la maggior parte degli impulsi dai gangli della base e dal cervelletto ed è coinvolta nella ricodificazione delle informazioni sui movimenti complessi.
L'irritazione della corteccia dell'area 6 provoca movimenti coordinati complessi, ad esempio rotazione della testa, degli occhi e del busto nella direzione opposta, contrazioni cooperative dei flessori o degli estensori sul lato opposto. La corteccia premotoria contiene centri motori associati funzioni sociali umano: il centro della parola scritta nella parte posteriore del giro frontale medio (campo 6), il centro motorio del linguaggio di Broca nella parte posteriore del giro frontale inferiore (campo 44), che fornisce la parola, così come il centro motorio musicale (campo 45), fornendo il tono del discorso, la capacità di cantare. La parte inferiore del campo b (sottocampo boro), situata nella zona del pneumatico, reagisce alla corrente elettrica con movimenti di masticazione ritmici. I neuroni della corteccia motoria ricevono input afferenti attraverso il talamo dai recettori muscolari, articolari e cutanei, dai gangli della base e dal cervelletto. La principale uscita efferente della corteccia motoria ai centri motori staminali e spinali sono le cellule piramidali dello strato V.
Nella parte posteriore del giro frontale medio si trova il centro oculomotore frontale, che controlla la concomitante e simultanea rotazione della testa e degli occhi (il centro di rotazione della testa e degli occhi nella direzione opposta). L'irritazione di questo centro fa girare la testa e gli occhi nella direzione opposta. La funzione di questo centro è di grande importanza nell'attuazione dei cosiddetti riflessi di orientamento (o riflessi “cos'è questo?”), che sono molto importanti per preservare la vita degli animali.
Anche la corteccia frontale degli emisferi cerebrali riceve Partecipazione attiva nella formazione del pensiero, organizzazione di attività mirate, pianificazione a lungo termine.
Struttura del lobo parietale
Il lobo parietale occupa le superfici laterali superiori dell'emisfero. Dal lobo frontale, il lobo parietale è limitato davanti e lateralmente dal solco centrale, dal lobo temporale in basso - dal solco laterale, dall'occipitale - da una linea immaginaria che va dal bordo superiore del parieto-occipitale solco al bordo inferiore dell'emisfero.
Sulla superficie superolaterale del lobo parietale ci sono tre giri: uno verticale - centrale posteriore e due orizzontali - parietale superiore e parietale inferiore. La parte del giro parietale inferiore, che circonda la parte posteriore del solco laterale, è chiamata regione sopramarginale (sopramarginale), la parte che circonda il giro temporale superiore è la regione nodale (angolare).
Il lobo parietale, come il lobo frontale, costituisce una parte significativa degli emisferi cerebrali. In termini filogenetici, è diviso in una vecchia sezione - la circonvoluzione centrale posteriore, una nuova - la circonvoluzione parietale superiore e una più recente - la circonvoluzione parietale inferiore.
La funzione del lobo parietale è associata alla percezione e all'analisi degli stimoli sensoriali e dell'orientamento spaziale. Diversi centri funzionali sono concentrati nelle circonvoluzioni del lobo parietale.
Nel giro centrale posteriore, i centri di sensibilità vengono proiettati con una proiezione corporea simile a quella del giro centrale anteriore. Il viso è proiettato nel terzo inferiore del giro, il braccio e il busto sono proiettati nel terzo medio e la gamba nel terzo superiore. Nel giro parietale superiore ci sono centri responsabili specie complesse sensibilità profonda: senso muscolo-articolare, spaziale bidimensionale, senso del peso e dell'ampiezza del movimento, senso di riconoscere gli oggetti al tatto.
Posteriormente alle parti superiori della circonvoluzione centrale posteriore è situato un centro che fornisce la capacità di riconoscere il proprio corpo, le sue parti, le loro proporzioni e le relative posizioni (campo 7).
I campi 1, 2, 3 della regione postcentrale costituiscono il nucleo corticale principale dell'analizzatore cutaneo. Insieme al campo 1, il campo 3 è la zona di proiezione primaria e il campo 2 è la zona di proiezione secondaria della pelle. analizzatore. La regione postcentrale è collegata da fibre efferenti alle formazioni subcorticali e staminali, alle aree precentrali e ad altre aree della corteccia cerebrale. Pertanto, la parte corticale dell'analizzatore sensibile è localizzata nel lobo parietale.
Aree sensoriali primarie – si tratta di aree della corteccia sensoriale, la cui irritazione o distruzione provoca cambiamenti evidenti e permanenti nella sensibilità del corpo (nuclei analizzatori, secondo I.P. Pavlov). Sono costituiti principalmente da neuroni unimodali e formano sensazioni della stessa qualità. Alle primarie aree sensoriali di solito c'è una chiara rappresentazione spaziale (topografica) delle parti del corpo e dei loro campi recettoriali.
Intorno alle zone sensoriali primarie sono meno localizzate aree sensoriali secondarie i cui neuroni rispondono all'azione di diversi stimoli, ad es. sono multimodali.
Le aree sensoriali più importanti sono corteccia parietale giro postcentrale e la parte corrispondente del lobulo paracentrale sulla superficie mediale degli emisferi, che è designata come area somatosensoriale I. Qui c'è una proiezione della sensibilità della pelle sul lato opposto del corpo dai recettori tattili, del dolore, della temperatura, della sensibilità interocettiva e della sensibilità del sistema muscolo-scheletrico - dai recettori muscolari, articolari e tendinei.
Oltre all'area somatosensoriale I, si distinguono le dimensioni più piccole somatosensoriale zona II, situato al confine dell'intersezione del solco centrale con il bordo superiore del lobo temporale, nella profondità del solco laterale. Il grado di localizzazione delle parti del corpo qui è meno pronunciato.
Situato nel lobo parietale inferiore centri di prassi. La prassi si riferisce a movimenti mirati che sono stati automatizzati nel processo di ripetizione ed esercizio, che si sviluppano nel processo di apprendimento e di pratica costante durante tutta la vita di un individuo. Camminare, mangiare, vestirsi, elemento meccanico della scrittura, varie tipologie attività lavorativa(ad esempio, i movimenti del conducente durante la guida, la falciatura, ecc.) sono prassi. La prassi è la più alta manifestazione della funzione motoria insita nell'uomo. Viene effettuato come risultato dell'attività combinata di varie aree della corteccia cerebrale.
Nelle parti inferiori del giro centrale anteriore e posteriore c'è centro di analisi degli impulsi interocettivi organi interni e navi. Il centro ha stretti collegamenti con le formazioni vegetative sottocorticali.
Struttura del lobo temporale
Il lobo temporale occupa la superficie inferolaterale degli emisferi. Dai lobi frontali e parietali, il lobo temporale è limitato dal solco laterale. Sulla superficie superolaterale del lobo temporale ci sono tre giri: superiore, medio e inferiore.
Il giro temporale superiore è situato tra la fessura silviana e quella temporale superiore, quello centrale è tra i solchi temporali superiore e inferiore, e quello inferiore è tra il solco temporale inferiore e la fessura midollare trasversa. Sulla superficie inferiore del lobo temporale si distinguono il giro temporale inferiore, il giro occipitotemporale laterale e il giro ippocampale (gamba del cavalluccio marino).
La funzione del lobo temporale è associata alla percezione delle sensazioni uditive, gustative, olfattive, all'analisi e alla sintesi dei suoni del linguaggio e ai meccanismi di memoria. Il principale centro funzionale della superficie laterale superiore del lobo temporale è situato nel giro temporale superiore. Qui si trova il centro della parola uditivo o gnostico (centro di Wernicke).
Una zona di proiezione primaria ben studiata lo è corteccia uditiva(campi 41, 42, 52), che si trova nella profondità del solco laterale (corteccia del giro temporale trasversale di Heschl). Comprende anche la corteccia di proiezione del lobo temporale centro di analisi vestibolare● nella circonvoluzione temporale superiore e media (campi 20 e 21).
L'area di proiezione olfattiva si trova nel giro dell'ippocampo, soprattutto nella sua sezione anteriore (il cosiddetto uncus). Accanto alle zone di proiezione olfattiva ci sono le zone gustative.
I lobi temporali svolgono un ruolo importante nell'organizzazione dei processi mentali complessi, in particolare della memoria.
Struttura del lobo occipitale
Il lobo occipitale occupa le parti posteriori degli emisferi. Sulla superficie convessa dell'emisfero, il lobo occipitale non ha confini netti che lo separano dai lobi parietale e temporale, ad eccezione della parte superiore del solco parieto-occipitale, che, situato sulla superficie interna dell'emisfero, separa il lobo parietale dal lobo occipitale. I solchi e le convoluzioni della superficie superolaterale del lobo occipitale non sono costanti e hanno una struttura variabile. Sulla superficie interna del lobo occipitale è presente un solco calcarino che separa il cuneus (un lobulo triangolare del lobo occipitale) dalla circonvoluzione linguale e dalla circonvoluzione occipitotemporale.
La funzione del lobo occipitale è associata alla percezione e all'elaborazione delle informazioni visive, all'organizzazione di complessi processi di percezione visiva - In questo caso, la metà superiore della retina è proiettata nell'area del cuneo, ricevendo luce dai campi inferiori di visione; nella regione del giro lingulare si trova la metà inferiore della retina dell'occhio, che percepisce la luce dai campi visivi superiori.
Situato nella corteccia occipitale area visiva primaria(corteccia di parte del giro sfenoide e del lobulo linguale, area 17). Qui c'è una rappresentazione topica dei recettori retinici. Ogni punto della retina corrisponde ad una propria sezione della corteccia visiva, mentre la zona della macula ha un'area di rappresentazione relativamente ampia. A causa della decussazione incompleta delle vie visive, le stesse metà della retina vengono proiettate nell'area visiva di ciascun emisfero. La presenza di una proiezione retinica in entrambi gli occhi di ciascun emisfero è la base della visione binoculare. Vicino al campo 17 c'è una corteccia area visiva secondaria(campi 18 e 19). I neuroni di queste zone sono multimodali e rispondono non solo alla luce, ma anche agli stimoli tattili e uditivi. La sintesi avviene in quest'area visiva vari tipi sensibilità, sorgono e vengono riconosciute immagini visive più complesse.
Insula, corteccia limbica
Isola, o il cosiddetto lobulo chiuso, è situato nella profondità del solco laterale. L'insula è separata dalle sezioni adiacenti adiacenti da una scanalatura circolare. La superficie dell'insula è divisa dalla scanalatura centrale longitudinale nelle parti anteriore e posteriore. Nell'isola viene proiettato un analizzatore del gusto.
Corteccia limbica. Sulla superficie interna degli emisferi sopra il corpo calloso si trova il giro del cingolo. Questo giro passa attraverso un istmo dietro il corpo calloso nel giro vicino al cavalluccio marino - il giro paraippocampale. Il giro cingolato, insieme al giro paraippocampale, costituisce il giro a volta.
Le superfici interna e inferiore degli emisferi sono combinate nella cosiddetta corteccia limbica (marginale) insieme al nucleo dell'amigdala dal gruppo dei nuclei sottocorticali, il tratto olfattivo e il bulbo, le aree dei lobi frontale, temporale e parietale del cervello corteccia, nonché la regione subtubercolare e la formazione reticolare del tronco. La corteccia limbica si unisce in una sola sistema funzionale– complesso limbico-reticolare. La funzione principale di queste parti del cervello non è tanto quella di fornire comunicazione mondo esterno, quanta regolazione del tono corticale, delle pulsioni e della vita affettiva. Regolano le funzioni complesse e sfaccettate degli organi interni e le reazioni comportamentali. Il complesso limbico-reticolare è il più importante sistema integrativo del corpo. Sistema limbicoè importante anche nella formazione della motivazione. La motivazione (o pulsione interna) comprende reazioni istintive ed emotive complesse (alimentari, difensive, sessuali). Il sistema limbico è coinvolto anche nella regolazione del sonno e della veglia.
Anche la corteccia limbica funziona funzione importante senso dell'olfatto. Odore: la percezione di ciò che è nell'aria sostanze chimiche. Il cervello olfattivo umano fornisce il senso dell'olfatto, nonché l'organizzazione di forme complesse di reazioni emotive e comportamentali. Il cervello olfattivo fa parte del sistema limbico.
Il cervello olfattivo è costituito da due sezioni: periferica e centrale. Dipartimento periferico rappresentato dal nervo olfattivo, bulbi olfattivi, centri olfattivi primari. La sezione centrale comprende il giro del cavalluccio marino: l'ippocampo, il giro dentato e quello a volta.
L'apparato recettore olfattivo è situato nella mucosa nasale. Attraverso il sistema di conduttori nervosi, le informazioni dai recettori vengono trasmesse alla sezione corticale dell'analizzatore olfattivo.
La parte corticale dell'analizzatore olfattivo si trova nel giro del cingolo, nel giro del cavalluccio marino e nell'uncus del cavalluccio marino, che insieme formano una regione chiusa a forma di anello. La sezione periferica dell'analizzatore olfattivo è collegata alle aree corticali di entrambi gli emisferi.
Il meccanismo fisiologico per la percezione degli odori da parte dell'analizzatore olfattivo non è del tutto chiaro. Esistono due ipotesi principali che spiegano la natura di questo processo da posizioni diverse. Secondo un'ipotesi, l'interazione tra le molecole odorose e i chemocettori avviene come una chiave e una serratura, cioè il tipo di molecola corrisponde a un recettore speciale. Un’altra ipotesi si basa sul presupposto che le molecole di una sostanza odorosa possiedano una certa onda vibratoria, sulla quale i recettori olfattivi sono “sintonizzati”. Le molecole che hanno vibrazioni simili devono avere un'onda comune e, di conseguenza, emanare odori simili.
Il termine “cervello olfattivo” in relazione alla fisiologia umana è alquanto arbitrario e non rivela pienamente la sua funzione multiforme e universale. La “localizzazione” del collegamento centrale del cervello olfattivo negli emisferi cerebrali non è casuale ed è il risultato dell’enorme ruolo “informativo” che l’olfatto ha svolto nel processo di evoluzione durante l’adattamento all’ambiente esterno e la regolazione di reazioni comportamentali complesse. Procurarsi il cibo, scegliere un individuo del sesso opposto, prendersi cura della prole, l'integrità del territorio, organizzare comunità di gruppo all'interno di una specie: tutte queste funzioni quotidiane in molti animali vengono eseguite con la partecipazione diretta di un sistema di ricezione olfattiva finemente costruito e del capacità di un numero di animali, in base a ciò, di inviare segnali differenziati sottili nell'ambiente esterno specifico sostanze odorose– segnali informatori.
Nella vita umana, l'olfatto ha perso il significato biologico informativo che aveva negli animali. Il sistema olfattivo umano è progettato sia per svolgere una funzione ristretta, “propria”, sia per una sorta di “carica” delle emozioni. Il potere d'influenza degli odori sulla sfera emotiva, il fatto che essi costituiscano il più importante “substrato alimentare delle emozioni”, è ben noto fin dall'antichità nella storia umana.
Il senso dell'olfatto di una persona può variare. Di norma queste variazioni sono insignificanti, ma in alcuni casi l'acutezza dell'olfatto può essere molto elevata (degustatori nell'industria dei profumi).
Poiché l'analizzatore olfattivo svolge un ruolo importante nella regolazione delle emozioni, esso dipartimento centrale appartengono al sistema limbico, chiamato figurativamente il “comune denominatore” di molte reazioni emotive e viscerosomatiche del corpo.
Centro di analisi del gusto si trova nelle immediate vicinanze del centro dell'analizzatore olfattivo, cioè nel corno dell'amo e dell'uncino, ma inoltre nella parte più bassa della circonvoluzione centrale posteriore (area 43), nonché nell'insula. Come l'analizzatore olfattivo, il centro svolge la funzione di proiezione, memorizzazione e riconoscimento delle immagini del gusto.
Situato al confine dei lobi temporale, occipitale e parietale centro di analisi del parlato scritto(campo 39), che è strettamente connesso con il centro Wernicke del lobo temporale, con il centro dell'analizzatore visivo del lobo occipitale, nonché con i centri del lobo parietale. Il centro di lettura fornisce il riconoscimento e l'archiviazione delle immagini della lingua scritta.
Informazioni di base Di L'ambiente e l'ambiente interno del corpo, ricevuto nella corteccia sensoriale, viene trasferito per ulteriore elaborazione alla corteccia associativa, dopo di che viene avviata una reazione comportamentale (se necessario) con la partecipazione obbligatoria della corteccia motoria.
corpo calloso– una lamina sottile arcuata, filogeneticamente giovane, collega le superfici mediane di entrambi gli emisferi. Esteso Parte di mezzo Il corpo calloso nella parte posteriore si ispessisce e nella parte anteriore si piega e si piega verso il basso in modo arcuato. Il corpo calloso collega le parti filogeneticamente più giovani degli emisferi e svolge un ruolo importante nello scambio di informazioni tra di loro.
Architettura della corteccia cerebrale
Viene chiamato lo studio delle caratteristiche strutturali della struttura della corteccia architettonica. Le cellule della corteccia cerebrale sono meno specializzate dei neuroni di altre parti del cervello; tuttavia, alcuni gruppi di essi sono anatomicamente e fisiologicamente strettamente correlati a determinate parti specializzate del cervello.
La struttura microscopica della corteccia cerebrale è diversa nelle sue diverse parti. Queste differenze morfologiche nella corteccia ci hanno permesso di identificare campi citoarchitettonici corticali separati. Esistono diverse opzioni per la classificazione dei campi corticali. La maggior parte dei ricercatori identifica 50 campi citoarchitettonici. La loro struttura microscopica è piuttosto complessa.
La corteccia è composta da 6 strati di cellule e relative fibre. Il tipo principale di struttura della corteccia è a sei strati, tuttavia non è uniforme ovunque. Ci sono aree della corteccia in cui uno degli strati è espresso in modo significativo e l'altro è espresso debolmente. In altre aree della corteccia alcuni strati sono suddivisi in sottostrati, ecc.
È stato stabilito che le aree della corteccia associate a una funzione specifica hanno una struttura simile. Le aree della corteccia che sono vicine nel loro significato funzionale negli animali e nell'uomo hanno una certa somiglianza nella struttura. Quelle parti del cervello che svolgono funzioni puramente umane (parola) sono presenti solo nella corteccia umana e sono assenti negli animali, anche nelle scimmie.
L'eterogeneità morfologica e funzionale della corteccia cerebrale ha permesso di identificare i centri della vista, dell'udito, dell'olfatto, ecc., che hanno una loro localizzazione specifica. Tuttavia non è corretto parlare del centro corticale come di un gruppo di neuroni strettamente limitato. La specializzazione delle aree della corteccia si forma nel processo della vita. All'inizio infanzia Le zone funzionali della corteccia si sovrappongono tra loro, quindi i loro confini sono vaghi e indistinti. Solo nel processo di apprendimento, accumulo propria esperienza nell'attività pratica si verifica una graduale concentrazione delle zone funzionali in centri separati gli uni dagli altri.
La sostanza bianca degli emisferi cerebrali è costituita da conduttori nervosi. In base alle caratteristiche anatomiche e funzionali, le fibre della sostanza bianca si dividono in associative, commissurali e di proiezione.
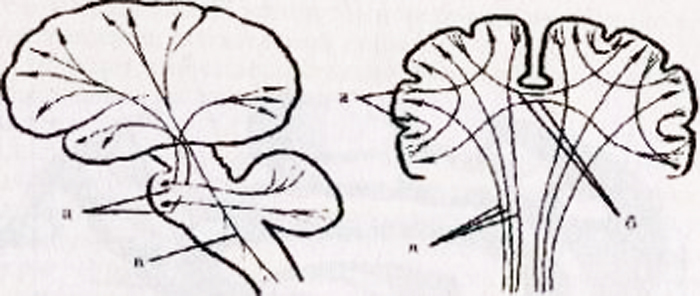
a – fibre associative; b – fibre commissurali; fibre di proiezione b.
Associativo le fibre si combinano varie aree corteccia all'interno di un emisfero. Queste fibre sono corte e lunghe. Le fibre corte solitamente hanno una forma arcuata e collegano i giri adiacenti. Le fibre lunghe collegano aree distanti della corteccia. Le fibre commissali sono solitamente chiamate quelle fibre che collegano aree topograficamente identiche degli emisferi destro e sinistro. Le fibre commissurali formano tre commissure: la commissura bianca anteriore, la commissura del fornice e il corpo calloso. La commissura bianca anteriore collega le aree olfattive degli emisferi destro e sinistro. La commessura del fornice collega il giro dell'ippocampo degli emisferi destro e sinistro. La maggior parte delle fibre commissurali passa attraverso il corpo calloso, collegando aree simmetriche di entrambi gli emisferi del cervello.
Proiettivo sono quelle fibre che collegano gli emisferi cerebrali con le parti sottostanti del cervello: il tronco e il midollo spinale. Le fibre di proiezione contengono percorsi che trasportano informazioni afferenti (sensibili) ed efferenti (motrici).
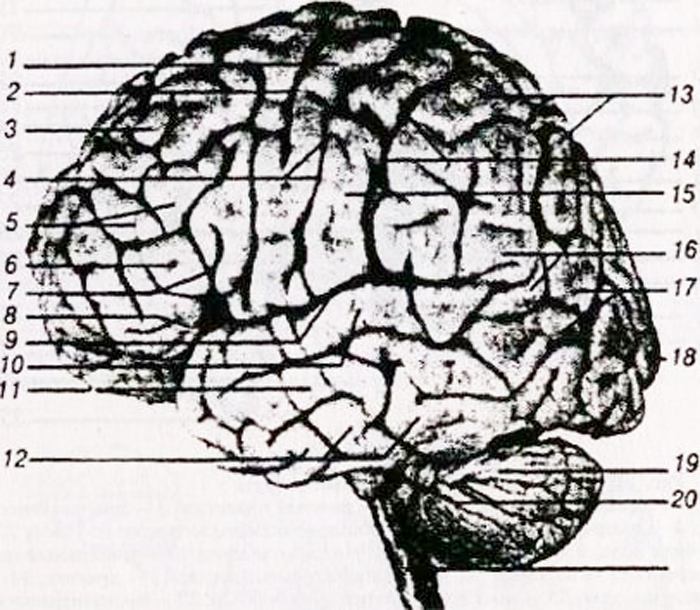
1 – giro precentrale; 2 – solco precentrale; 3 – giro frontale superiore; 4 – scanalatura centrale; 5 – giro frontale medio;
b – giro frontale inferiore; 7 – ramo ascendente del solco laterale;
8 – ramo orizzontale del solco laterale; 9 – ramo posteriore del solco laterale; 10 – giro temporale superiore; 11 – giro temporale medio;
12 – giro temporale inferiore; 13 – lobulo parietale; 14 – solco postcentrale; 15 – giro postcentrale; 16 – giro sopramarginale;
17 – giro angolare; 18 – lobo occipitale; 19 – cervelletto; 20 – fessura orizzontale del cervelletto; 21 – midollo allungato

