Refleksi- hermoston suorittama kehon reaktio reseptorien ärsytykseen.
refleksikaari - polku, jota pitkin hermoimpulssi kulkee refleksin toteuttamisen aikana. Koostuu 5 osasta:
- reseptori - herkkä muodostus, joka pystyy reagoimaan tietyntyyppisiin ärsykkeisiin;
- herkkä neuroni johtaa impulsseja aivoihin;
- interkalaarinen neuroni yhdistää herkkiä ja toimeenpanohermosoluja, sijaitsee selkäytimessä tai aivoissa;
- toimeenpaneva (motorinen) neuroni johtaa impulsseja aivoista toimeenpanoelimiin;
- toimeenpanovirasto- lihakset (sopimukset), rauhaset (salaisuudet) jne.
Ehdottomia refleksejä
- läsnä syntymästä lähtien
- elämän aikana ne eivät muutu eivätkä katoa;
- ovat samat kaikissa saman lajin organismeissa;
- sopeuttaa keho pysyviin olosuhteisiin.
Ehdolliset refleksit
- hankittu elämän aikana;
- voi muuttua tai kadota;
- jokaisella organismilla on omansa;
- mukauttaa keho muuttuviin olosuhteisiin.
Testit
1) Mikä linkki toimii heijastuskaaren alussa
A) interkalaarinen neuroni
B) herkkä neuroni
B) reseptori
D) toimeenpaneva neuroni
2. Missä järjestyksessä heijastuskaaren komponentit ovat mukana refleksin toteutuksessa
A) toimeenpaneva elin, motorinen neuroni, interkalaarinen neuroni, sensorinen neuroni, reseptori
B) interkalaarinen neuroni, sensorinen neuroni, motorinen neuroni, reseptori, toimeenpaneva elin
C) reseptori, sensorinen neuroni, interkalaarinen neuroni, motorinen neuroni, toimeenpaneva elin
D) sensorinen neuroni, interkalaarinen neuroni, reseptori, toimeenpaneva elin, motorinen neuroni
3. Refleksikaaren alkuvaihe sylkirefleksissä
A) sylkirauhanen
B) reseptori
B) interkalaarinen neuroni
D) toimeenpaneva neuroni
4. ärsykkeiden toiminta aiheuttaa tapahtuman hermo impulssi V
A) sensoriset neuronit
B) motoriset neuronit
B) reseptorit
D) interkalaariset neuronit
5. Herätys suunnataan herkkää neuronia pitkin
A) keskushermostoon
B) toimeenpanevalle elimelle
B) reseptoreihin
D) lihaksille
6. Reseptorit ovat herkkiä muodostelmia, jotka
A) välittää impulsseja keskushermostoon
B) välittää hermoimpulsseja interkalaarisista hermosoluista toimeenpanoelimiin
C) havaita ärsykkeitä ja muuntaa ärsykkeiden energian hermostuneeksi viritysprosessiksi
D) vastaanottaa hermoimpulsseja sensorisista neuroneista
7. Hermoimpulssit välittyvät aivoihin hermosolujen kautta
A) moottori
B) intercalary
B) herkkä
D) johtaja
8. Refleksikaari päättyy
A) johtaja
B) herkkä neuroni
B) reseptori
D) interkalaarinen neuroni
9. Hermoimpulssit reseptoreista keskushermostoon johtavat
A) sensoriset neuronit
B) motoriset neuronit
C) sensoriset ja motoriset neuronit
D) interkalaariset ja motoriset neuronit
10. Hermomuodostelmia ihmiskehossa, havaitsevat ärsytystä ulkoisesta ympäristöstä
A) hermot
B) reseptorit
B) aivot
D) hermosolmukkeet
11. Ärsykkeiden muuttuminen hermoimpulssiksi tapahtuu vuonna
A) motoriset hermot
B) aivot
B) reseptorit
D) selkäydin
12. Hermoimpulssit välittyvät ihmisen aisteista neuronien kautta
A) herkkä
B) moottori
B) lisää
D) johtaja
13. Havaitse tiedot ja muunna se hermoimpulsseiksi
A) reseptorit
B) hermosolmukkeet
B) toimeenpanoelimet
D) interkalaariset neuronit
14. Ehdollinen refleksi
A) periytyy jälkeläisille, mutta se ei säily koko elämän ajan
B) elimistö hankkii ja menettää helposti elämän aikana
C) periytyvät jälkeläisiltä vanhemmilta ja säilyy
D) on kehon jatkuva reaktio tiukasti määriteltyihin ympäristön ärsykkeisiin
15. Esimerkki ehdollista refleksistä kissalla
A) syljeneritys syömisen aikana
B) tassun vetäytyminen palovamman aikana
B) reaktio lempinimeen
D) reaktio terävään ääneen
16. Syljeneritys ihmisillä sitruunan silmissä - refleksi
A) ehdollinen
B) ehdoton
B) suojaava
D) ohjeellinen
17) Ehdolliset refleksit ihmisillä muodostuvat prosessissa
A) yksilöllinen kehitys
B) lajien muodostuminen
C) historiallinen kehitys
D) alkion muodostuminen
18. Esimerkki ehdollisesta refleksistä on
A) syljeneritys leikattua sitruunaa näkemällä
B) syljeneritys ruokinnan aikana
C) pupillin supistuminen voimakkaalle valolle altistettuna
D) käden vetäminen pois äkillisellä injektiolla
19. Lapsen reaktio maitopulloon on refleksi, joka
A) on peritty
B) muodostuu ilman aivokuoren osallistumista pallonpuoliskot
B) hankitaan eliniän aikana
D) säilyy läpi elämän
20. Suojaava aivastelurefleksi
A) ei ole peritty
B) on ehdollinen
B) heikkenee ajan myötä
D) ominaisuus kaikille lajin yksilöille
21. Reaktioita, joita ihmiset ja eläimet saavat elämän aikana ja jotka tarjoavat sopeutumisen muuttuviin ympäristöolosuhteisiin, kutsutaan
A) ehdolliset refleksit
B) vaistot
B) ehdottomat refleksit
D) refleksit, jotka ovat perinnöllisiä
22. Ehdoton refleksi
A) on peritty
B) hankittu elämän aikana
C) tuotetaan tietyille signaaleille
D) on erilaisten ulkoisten signaalien taustalla
23. Ihmisten ja eläinten ehdottomat refleksit tarjoavat
C) uusien motoristen taitojen hallitseminen kehon toimesta
24. Ihmisen reaktio vihreä väri liikennevalo on refleksi
A) synnynnäinen
B) hankittu
B) ehdoton
D) perinnöllinen
25. Korosta mahanestettä vasteena ruoan ärsykereseptoreille suuontelon
A) jännitys
B) jarrutus
B) ehdoton refleksi
D) itsesääntely
26. Ehdollisten refleksien biologinen merkitys keholle on, että ne
A) ylläpitää sisäisen ympäristön vakautta
B) lajikohtainen ja säilyy läpi elämän
B) edistää vuorovaikutusta toiminnallisia järjestelmiä
D) tarjota sopeutumista muuttuviin ympäristöolosuhteisiin
27. Elämän aikana eläimet hankkivat refleksejä
A) ehdoton
B) perinnöllinen
C) ominaisuus tietyn lajin kaikille yksilöille
D) antaa heidän selviytyä muuttuvissa olosuhteissa
28. Ehdollinen refleksi ihmisillä on
A) pupillien supistuminen kirkkaassa valossa
B) pään kääntäminen terävälle äänelle
C) syljen eritystä, kun ruoka joutuu suuhun
D) reaktio sanan merkitykseen
29. Esimerkki ehdollisesta refleksistä on
1) kyky ajaa pyörää
2) syljeneritys syödessä
3) halu nukkua väsyneenä
4) aivastelu ja yskiminen
30. Ihmisten ja eläinten ehdolliset refleksit tarjoavat
A) organismin sopeutuminen jatkuviin ympäristöolosuhteisiin
B) kehon sopeutuminen muuttuvaan ulkoiseen maailmaan
C) uusien motoristen taitojen organismien kehittäminen
D) kouluttajan käskyjen erottaminen eläinten välillä
31. Synnynnäiset refleksit
A) ovat organismin lajiominaisuuksia
B) lajin yksittäisille yksilöille ominaista
C) vaatia täytäntöönpanoa koskevia lisäehtoja
D) eivät ole periytyviä
päämuoto hermostunut toiminta on refleksi. Refleksi - kehon kausaalinen reaktio ulkoisen tai sisäisen ympäristön muutoksiin, joka suoritetaan keskusyksikön pakollisella osallistumisella hermosto vasteena reseptoristimulaatiolle. Refleksien vuoksi tapahtuu kehon minkä tahansa toiminnan ilmaantuminen, muutos tai lakkaaminen.
Hermopolkua, jota pitkin viritys leviää refleksien toteutuksen aikana, kutsutaan refleksikaari.
Refleksikaaret koostuvat viidestä komponentista: 1) reseptori; 2) afferentti hermopolku; 3) refleksikeskus; 4) efferentti hermopolku; 5) efektori (työkappale).
Reseptori- Tämä on herkkä hermopääte, joka havaitsee ärsytystä. Reseptoreissa ärsykkeen energia muunnetaan hermoimpulssin energiaksi. Erota: 1) exteroreseptoreita- kiihtynyt ympäristön ärsytyksen vaikutuksesta (ihon, silmien, sisäkorva, nenän ja suun limakalvot); 2) interoreseptoreita- havaita ärsykkeitä kehon sisäisestä ympäristöstä (reseptorit sisäelimet, alukset); 3) proprioreseptoreita- reagoida yksittäisten kehon osien asennon muutokseen avaruudessa (lihasten, jänteiden, nivelsiteiden, nivelpussien reseptorit).
Afferentti hermopolku joita edustavat reseptorineuronien prosessit, jotka kuljettavat viritteitä keskushermostoon.
refleksikeskus koostuu ryhmästä hermosoluja, jotka sijaitsevat keskushermoston eri tasoilla ja välittävät hermoimpulsseja afferentista efferentille hermoreitille.
efferentti hermopolku johtaa hermoimpulsseja keskushermostosta efektoriin.
Efektori- toimeenpaneva elin, jonka toiminta muuttuu hermoimpulssien vaikutuksesta, jotka tulevat siihen refleksikaaren muodostumien kautta. Effektorit voivat olla lihaksia tai rauhasia.
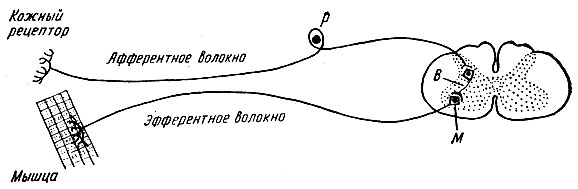
refleksikaaria voi olla yksinkertainen tai monimutkainen. Yksinkertainen refleksikaari koostuu kahdesta hermosolusta - havaitsevasta ja efektorista, joiden välissä on yksi synapsi. Kaavio tällaisesta kahden neuronin refleksikaaresta on esitetty kuvassa 1. 71.
Esimerkki yksinkertaisesta heijastuskaaresta on jännerefleksikaaret, kuten polvilumpion heijastuskaari.
Useimpien refleksien refleksikaarit eivät sisällä kahta, vaan suuremman määrän hermosoluja: reseptorin, yhden tai useamman interkalaarin ja efektorin. Tällaisia refleksikaaria kutsutaan monimutkaisiksi, monineuronaaleiksi. Kaavio kompleksisesta (kolmen neuronista) refleksikaaresta on esitetty kuvassa. 72.
Nyt on todettu, että efektorin vasteen aikana useat työelimessä olevat hermopäätteet kiihtyvät. Hermoimpulssit tulevat nyt efektorista jälleen keskushermostoon ja kertovat sille toimivan elimen oikeasta vasteesta. Siten refleksikaaret eivät ole avoimia, vaan rengasmuodostelmia.
Refleksit ovat hyvin erilaisia. Ne voidaan luokitella useiden kriteerien mukaan: 1) biologinen merkitys(ruoka, puolustus, seksuaalinen); 2) ärtyneiden reseptorien tyypistä riippuen: eksteroseptiiviset, interoseptiiviset ja proprioseptiiviset; 3) vasteen luonteen mukaan: motorinen tai motorinen (toimeenpanoelin - lihas), erittävä (efektori - rauta), vasomotorinen (verisuonten supistuminen tai laajeneminen).
Kaikki koko organismin refleksit voidaan jakaa kahteen osaan suuria ryhmiä: ehdoton ja ehdollinen. Niiden välisiä eroja käsitellään luvussa XII.
Hermoston toiminnalla on refleksi. Jopa XVII vuosisadalla. Ranskalainen filosofi ja matemaatikko René Descartes kuvaili refleksitoimintaa. Hän pani merkille kehon reaktion ärsytykseen ja ehdotti polun olemassaoloa, jota pitkin hermostuneisuus kulkee. Itse termin "refleksi" esitti myöhemmin - 1700-luvulla - tšekkiläinen tiedemies J. Prochazka (latinan sanasta "reflex" - heijastuva toiminta). Myöhemmin I. M. Sechenov työssään "Aivojen refleksit" osoitti, että hermoston vasteet monenlaisiaärsytykset etenevät refleksimekanismin mukaan, ts. kaikki tietoiset ja tiedostamattomat teot ovat refleksiperäisiä. Refleksi ymmärretään kehon erityiseksi reaktioksi sisäisen ympäristön ärsykkeeseen tai ulkopuolelta vastaanotetuksi keskushermoston pakollisella osallistumisella. Refleksejä kutsutaan yleensä hermoston toiminnallisiksi yksiköiksi.
Refleksikaaren refleksien mekanismit
Refleksit voidaan luokitella eri kriteerien mukaan. Joten kaaren sulkemisasteesta riippuen, ts. refleksikeskuksen sijainnin mukaan refleksit jaetaan selkäytimeen (refleksi sulkeutuu selkäytimeen), bulbariin (heijastuskeskus - ydin), mesenkefaaliset (refleksikaaren rakenne on suljettu keskiaivoissa), väliaivo- ja kortikaalinen refleksikeskukset sijaitsevat telencephalon ja aivokuori, vastaavasti. Effektoriominaisuuden mukaan ne ovat somaattisia, kun refleksin efferenttipolku tuottaa luustolihasten motorista hermotusta, ja vegetatiivisia, kun sisäelimet ovat efektoreina. Ärsytyneiden reseptorien tyypistä riippuen refleksit jaetaan eksteroseptiivisiin (jos reseptori havaitsee tietoa ulkoisesta ympäristöstä), proprioseptiivisiin (refleksikaari alkaa tuki- ja liikuntaelinten reseptoreista) ja interoseptiivisiin (sisäelinten reseptoreista). Interoseptiiviset refleksit puolestaan jaetaan viskero-viskeraalisiin (refleksikaari yhdistää kaksi sisäelintä), viskero-lihaksiseen (reseptorit sijaitsevat lihas-jännelaitteistossa, efektori on sisäelin) ja viskero-kutaanisiin (reseptorit ovat paikallinen ihoon, työelimiin - sisäelimet). Pavlovin mukaan refleksit jaetaan ehdollisiin (kehittyneet elämän aikana, kullekin yksilölle ominaiset) ja ehdoitta (synnynnäiset, lajikohtaiset: ruoka, seksuaalinen, puolustava-motorinen, homeostaattinen jne.).
Miten refleksikaari toimii?
Refleksin tyypistä riippumatta sen refleksikaari sisältää reseptorin, afferentin polun, hermokeskuksen, efferenttipolun, työelimen ja palautteen. Poikkeuksen muodostavat aksonirefleksit, joiden refleksikaaren rakenne sijaitsee yhdessä neuronissa: aistiprosessit synnyttävät keskipituisia impulsseja, jotka neuronin kehon läpi kulkevat aksonia pitkin keskushermostoon ja haaraa pitkin. aksonin impulssit saavuttavat efektorin. Tällaiset refleksit johtuvat metasympaattisen hermoston toiminnasta, joiden kautta suoritetaan esimerkiksi verisuonten sävyn ja ihorauhasten toiminnan säätelymekanismeja.
Ärsytyksen havaitseminen ja sen muuntaminen viritysenergiaksi suoritetaan refleksikaarien reseptorien avulla. Herätyksen reseptorienergialla on paikallisen vasteen luonne, mikä on tärkeää virityksen voimakkuuden asteittamisessa.
Reseptorien rakenteen ja alkuperän perusteella ne voidaan jakaa primaarisiin sensorisiin, sekundaarisiin sensorisiin ja vapaisiin hermopäätteisiin. Edellisessä neuroni itse toimii reseptorina (se kehittyy neuroepiteelistä); ärsykkeen ja ensimmäisen afferentin neuronin välillä ei ole välirakenteita. Primaaristen sensoristen reseptorien paikallinen vaste - reseptoripotentiaali - on myös generaattoripotentiaali, ts. indusoimalla toimintapotentiaalin afferentin kuidun kalvon yli. Ensisijaisia sensorisia reseptoreja ovat näkö-, haju-, kemo- ja baroreseptorit. sydän- ja verisuonijärjestelmästä.
Toissijaisesti tunnistavat solut ovat ei-hermostoperäisiä erityisrakenteita, jotka ovat vuorovaikutuksessa pseudo-unipolaaristen sensorisolujen dendriittien kanssa synaptisten neuroreseptorikontaktien avulla. Reseptoripotentiaali, joka syntyy ärsykkeen vaikutuksesta sekundaarisen tunnistavissa soluissa, ei ole generaattori eikä aiheuta toimintapotentiaalin ilmaantumista afferentin kuidun kalvolle. Eksitatorinen postsynaptinen potentiaali syntyy vain reseptorisolun välittäjän vapautumismekanismin kautta. Ärsykkeen voimakkuuden gradaatio tapahtuu erittämällä eri määriä välittäjää (mitä enemmän välittäjä vapautuu, sitä voimakkaampi ärsyke).
Toissijaisia aistisoluja ovat kuulo-, vestibulaari-, kaulavaltimo-, tunto- ja muut reseptorit. Joskus toiminnan erityispiirteistä johtuen tähän ryhmään kuuluvat fotoreseptorit, jotka anatomisesta näkökulmasta ja neuroepiteelistä lähtönsä vuoksi ovat toissijaisesti tunnistavia.
Vapaat hermopäätteet ovat pseudo-unipolaaristen sensorisolujen haarautuvia dendriittejä, ja niitä löytyy lähes kaikissa ihmiskehon kudoksissa.
Sen ärsykkeen energialuonteen mukaan, johon reseptori reagoi, ne jaetaan mekanoreseptoreihin (taktiili-, baroreseptorit, tilavuusreseptorit, kuulo-, vestibulaariset; ne yleensä havaitsevat mekaanisen ärsytyksen solukasvun avulla), kemoreseptoreihin ( haju), verisuonten kemoreseptorit, keskushermosto, fotoreseptorit (havaitsevat ärsytystä sauva- ja kartiomaisten solujen kasvamien kautta), lämpöreseptorit (reagoivat "kylmä-lämpimän" muutokseen - Rufini-kappaleet ja limakalvojen Krause-pullot) ja nosiseptorit (kapseloimattomat kipupäätteet).
Reseptorin jälkeiset heijastuskaaren muodostelmat
Refleksikaarien rakenteen post-reseptorin muodostuminen on pseudounipolaarisen sensorisen neuronin muodostama afferenttipolku, jonka runko sijaitsee selkäydin ganglio, ja muodostuu aksoneja takajuuret selkäydin. Afferentin reitin tehtävänä on johtaa informaatiota keskuslinkille, lisäksi tässä vaiheessa tietoa koodataan. Näihin tarkoituksiin selkärankaisten kehossa käytetään binäärikoodia, joka koostuu impulssien purskeista (volleyista) ja niiden välisistä aukoista. On olemassa kaksi päätyyppiä koodausta: taajuus ja spatiaalinen.
Ensimmäinen on eri impulssien muodostuminen purskeessa, eri määrä purskeita, niiden kesto ja niiden välisten taukojen kesto riippuen reseptoriin kohdistetun stimulaation voimakkuudesta. Spatiaalinen koodaus suorittaa ärsykkeen voimakkuuden gradaatiota, johon liittyy erilainen määrä hermosäikeitä, joita pitkin viritys tapahtuu samanaikaisesti.
Afferenttireitti koostuu pääasiassa A-α, A-β ja A-δ kuiduista.
Kuitujen läpi kulkenut hermoimpulssi tulee refleksikeskukseen, joka anatomisessa mielessä on kokoelma neuroneja, jotka sijaitsevat tietyllä keskushermoston tasolla ja osallistuvat tämän refleksin muodostumiseen. Refleksikeskuksen tehtävänä on analysoida ja syntetisoida tietoa sekä vaihtaa tietoa afferentista efferentille.
Refleksikaaritoiminnot
Riippuen hermoston osastosta (somaattinen ja autonominen), refleksit, joiden keskus sijaitsee selkäytimessä, eroavat interkalaaristen hermosolujen sijainnista. Joten somaattisen hermoston kohdalla refleksikeskus sijaitsee selkäytimen etu- ja takasarvien välissä. Autonomisen hermoston refleksikeskus (välisten hermosolujen runko) sijaitsee takasarvet. Hermoston somaattiset ja autonomiset osat eroavat myös efferenttien hermosolujen sijainnista. Somaattisen hermoston motoristen neuronien ruumiit sijaitsevat selkäytimen etusarvissa, preganglionisten hermosolujen rungot autonominen järjestelmä- keskisarvien tasolla.
Molempien solutyyppien aksonit muodostavat refleksikaaren efferentin polun. Somaattisessa hermostossa se on jatkuvaa, se koostuu A-α-tyypin kuiduista. Ainoat poikkeukset ovat A-y-säikeet, jotka johtavat virityksen selkäytimen soluista lihaskarojen intrafusaalisiin kuituihin. Autonomisen hermoston efferenttireitti katkeaa autonominen ganglio sijaitsee tai intramuraalinen (pari sympaattinen osa) tai lähellä selkäydintä (yksin tai sisällä sympaattinen runko- sympaattinen osa). Preganglioninen kuitu kuuluu B-kuituihin ja postganglioninen kuitu C-ryhmään.
Hermoston somaattisen osan työelin on poikkijuovainen luurankolihas, vegetatiivisessa kaaressa efektori on rauhanen tai lihas (sileä tai poikkijuovainen sydän). Välillä tehokkaalla tavalla ja työelin on kemiallinen myoneuraalinen tai neurosekretiivinen synapsi.
Refleksikaari sulkeutuu renkaaksi käänteisen afferentaation ansiosta - impulssien virtaus efektorireseptoreista takaisin refleksikeskukseen. Palautetoiminto - signalointi keskushermostolle suoritetusta toiminnasta. Jos sitä ei suoriteta tarpeeksi, hermokeskus innostuu - refleksi jatkuu. Myös käänteisen afferentaation ansiosta keskushermoston perifeeristä toimintaa ohjataan.
Erota negatiivinen ja positiivinen palaute. Ensimmäinen, suorittaessaan tiettyä toimintoa, käynnistää mekanismin, joka estää tämän toiminnon. Positiivinen palaute koostuu jo suoritettavan toiminnon lisästimulaatiosta tai jo masentuneen toiminnon estämisestä. Positiivinen käänteinen afferentaatio on harvinaista, koska se saattaa biologisen järjestelmän epävakaaseen asentoon.
Yksinkertaiset (monosynaptiset) refleksikaarit koostuvat vain kahdesta neuronista (afferentti ja efferentti) ja eroavat toisistaan vain proprioseptiivisissä reflekseissä. Loput kaaret sisältävät kaikki edellä mainitut komponentit.
Refleksikaari on neuronien ketju ääreisreseptorista keskushermoston kautta perifeeriseen efektoriin. Refleksikaaren elementit ovat perifeerinen reseptori, afferenttireitti, yksi tai useampi interneuroni, efferenttipolku ja efektori.
Kaikki reseptorit osallistuvat tiettyihin reflekseihin, joten niiden afferenttisäikeet toimivat vastaavan refleksikaaren afferenttipoluna. Interneuronien lukumäärä on aina suurempi kuin yksi, paitsi monosynaptinen venytysrefleksi. Efferenttireittiä edustavat joko autonomisen hermoston motoriset aksonit tai postganglioniset kuidut, ja efektoreita ovat luustolihakset ja sileät lihakset, sydän ja rauhaset.
Aikaa ärsykkeen alkamisesta efektorin vasteeseen kutsutaan refleksiajaksi. Useimmissa tapauksissa sen määrää pääasiassa johtumisaika afferentissa ja efferentissä reitissä sekä refleksikaaren keskiosassa, johon pitäisi lisätä reseptorissa olevan ärsykkeen muuttumisaika eteneväksi impulssiksi, aika. transmissio keskushermoston synapsien kautta (synaptinen viive), aika, joka siirtyy efferentistä reitistä efektoriin ja efektorin aktivaatioaika.
Refleksikaaret on jaettu useisiin tyyppeihin
1. Monosynaptiset refleksikaaret - vain yksi keskushermostossa sijaitseva synapsi osallistuu tällaiseen kaariin. Tällaiset refleksit ovat hyvin yleisiä kaikilla selkärankaisilla, ne ovat mukana säätelyssä lihasten sävy ja asento (esim. polven nykiminen). Näissä kaarissa hermosolut eivät pääse aivoihin, ja refleksit suoritetaan ilman sen osallistumista, koska ne ovat stereotyyppisiä eivätkä vaadi ajattelua tai tietoista päätöstä. Ne ovat taloudellisia asiaan liittyvien keskushermosolujen lukumäärän suhteen, eivätkä ne tarvitse aivojen toimintaa.
2. Polysynaptiset selkäydinrefleksikaaret - niissä on vähintään kaksi keskushermostossa sijaitsevaa synapsia, koska kaaressa on kolmas neuroni - intercalary eli välihermosolu. Täällä on synapseja sensorisen hermosolun ja interneuronien välillä sekä interkalaarien ja motoristen neuronien välillä. Tällaiset refleksikaaret antavat keholle mahdollisuuden suorittaa automaattisia tahattomia reaktioita, jotka ovat tarpeen sopeutuakseen ulkoisen ympäristön muutoksiin (esim. pupillirefleksi tai tasapainon säilyttäminen liikkeessä) ja kehon itseensä tapahtuviin muutoksiin (hengitysnopeuden, verenpaineen säätelyyn jne.).
3. Polysynaptiset refleksikaaret, joissa on mukana sekä selkäydin että aivot - tämäntyyppisissä refleksikaarissa selkäytimessä on synapsi aistihermosolun ja aivoihin impulsseja lähettävän hermosolun välillä.
Refleksit voidaan luokitella eri kriteerien mukaan. Joten kaaren sulkemisasteesta riippuen, ts. refleksikeskuksen sijainnin mukaan refleksit jaetaan selkäytimeen (refleksi sulkeutuu selkäytimeen), bulbar (refleksikeskus on pitkittäisydin), mesenkefaalisiin (refleksikaari on kiinni väliaivoissa), väliaivoissa ja väliaivoissa. aivokuoren refleksikeskukset sijaitsevat suurten pallonpuoliskojen telencephalonissa ja aivokuoressa.
Effektoriominaisuuden mukaan ne ovat somaattisia, kun refleksin efferenttipolku tuottaa luustolihasten motorista hermotusta, ja vegetatiivisia, kun sisäelimet ovat efektoreina.
Ärsytyneiden reseptorien tyypistä riippuen refleksit jaetaan eksteroseptiivisiin (jos reseptori havaitsee tietoa ulkoisesta ympäristöstä), proprioseptiivisiin (refleksikaari alkaa tuki- ja liikuntaelinten reseptoreista) ja interoseptiivisiin (sisäelinten reseptoreista).
Interoseptiiviset refleksit puolestaan jaetaan viskero-viskeraalisiin (refleksikaari yhdistää kaksi sisäelintä), viskero-lihaksiseen (reseptorit sijaitsevat lihas-jännelaitteistossa, efektori on sisäelin) ja viskero-kutaanisiin (reseptorit ovat paikallinen ihoon, työelimiin - sisäelimet).
Pavlovin mukaan refleksit jaetaan ehdollisiin (kehittyneet elämän aikana, kullekin yksilölle ominaiset) ja ehdoitta (synnynnäiset, lajikohtaiset: ruoka, seksuaalinen, puolustava-motorinen, homeostaattinen jne.).
Refleksin tyypistä riippumatta sen refleksikaari sisältää reseptorin, afferentin polun, hermokeskuksen, efferenttipolun, työelimen ja palautteen. Poikkeuksena ovat aksonirefleksit, joiden refleksikaari sijaitsee yhdessä neuronissa: aistiprosessit synnyttävät keskipituisia impulsseja, jotka neuronin kehon läpi kulkevat aksonia pitkin keskushermostoon ja aksonin haaraa pitkin. , impulssit saavuttavat efektorin. Tällaiset refleksit johtuvat metasympaattisen hermoston toiminnasta, joiden kautta suoritetaan esimerkiksi verisuonten sävyn ja ihorauhasten toiminnan säätelymekanismeja.
Ärsytyksen havaitseminen ja sen muuntaminen viritysenergiaksi suoritetaan refleksikaarien reseptorien avulla. Herätyksen reseptorienergialla on paikallisen vasteen luonne, mikä on tärkeää virityksen voimakkuuden asteittamisessa.
Reseptorien rakenteen ja alkuperän perusteella ne voidaan jakaa primaarisiin sensorisiin, sekundaarisiin sensorisiin ja vapaisiin hermopäätteisiin. Edellisessä neuroni itse toimii reseptorina (se kehittyy neuroepiteelistä); ärsykkeen ja ensimmäisen afferentin neuronin välillä ei ole välirakenteita. Primaaristen sensoristen reseptorien paikallinen vaste - reseptoripotentiaali - on myös generaattoripotentiaali, ts. indusoimalla toimintapotentiaalin afferentin kuidun kalvon yli. Primaarisia sensorisia reseptoreja ovat kardiovaskulaarijärjestelmän näkö-, haju-, kemo- ja baroreseptorit.
Toissijaisesti tunnistavat solut ovat ei-hermostoperäisiä erityisrakenteita, jotka ovat vuorovaikutuksessa pseudo-unipolaaristen sensorisolujen dendriittien kanssa synaptisten neuroreseptorikontaktien avulla. Reseptoripotentiaali, joka syntyy ärsykkeen vaikutuksesta toissijaisesti tunnistavissa soluissa, ei ole generaattori eikä aiheuta toimintapotentiaalin ilmaantumista afferentin kuidun kalvolle. Eksitatorinen postsynaptinen potentiaali syntyy vain reseptorisolun välittäjän vapautumismekanismin kautta. Ärsykkeen voimakkuuden gradaatio tapahtuu erittämällä eri määriä välittäjää (mitä enemmän välittäjä vapautuu, sitä voimakkaampi ärsyke).
Toissijaisia aistisoluja ovat kuulo-, vestibulaari-, kaulavaltimo-, tunto- ja muut reseptorit. Joskus toiminnan erityispiirteistä johtuen tähän ryhmään kuuluvat fotoreseptorit, jotka anatomisesta näkökulmasta ja neuroepiteelistä lähtönsä vuoksi ovat toissijaisesti tunnistavia.
Vapaat hermopäätteet ovat pseudo-unipolaaristen sensorisolujen haarautuvia dendriittejä, ja niitä löytyy lähes kaikissa ihmiskehon kudoksissa.
Sen ärsykkeen energialuonteen mukaan, johon reseptori reagoi, ne jaetaan mekanoreseptoreihin (taktiili-, baroreseptorit, volumoreseptorit, kuulo-, vestibulaariset; ne yleensä havaitsevat mekaanisen ärsytyksen solukasvun avulla), kemoreseptoreihin (hajureseptoreihin). ), verisuonten kemoreseptorit, keskushermosto , fotoreseptorit (havaitsevat ärsytystä sauvan ja kartion muotoisten solujen kasvun kautta), lämpöreseptorit (reagoivat "kylmä-lämpimän" muutokseen - Rufini-kappaleet ja limakalvojen Krause-pullot) ja nosiseptorit (kapseloimattomat kipupäätteet).
Reseptorin jälkeinen refleksikaarien muodostus on pseudounipolaarisen sensorisen hermosolun muodostama afferenttipolku, jonka runko sijaitsee selkäytimen gangliossa ja aksonit muodostavat selkäytimen takajuuret. Afferentin reitin tehtävänä on johtaa informaatiota keskuslinkille, lisäksi tässä vaiheessa tietoa koodataan. Näihin tarkoituksiin selkärankaisten kehossa käytetään binäärikoodia, joka koostuu impulssien purskeista (volleyista) ja niiden välisistä aukoista. On olemassa kaksi päätyyppiä koodausta: taajuus ja spatiaalinen.
Ensimmäinen on eri impulssien muodostuminen purskeessa, eri määrä purskeita, niiden kesto ja niiden välisten taukojen kesto riippuen reseptoriin kohdistetun stimulaation voimakkuudesta. Spatiaalinen koodaus suorittaa ärsykkeen voimakkuuden gradaatiota, johon liittyy erilainen määrä hermosäikeitä, joita pitkin viritys tapahtuu samanaikaisesti.
Afferenttireitin koostumus sisältää pääasiassa A-b-, A-c- ja A-d-kuituja.
Kuitujen läpi kulkenut hermoimpulssi tulee refleksikeskukseen, joka anatomisessa mielessä on kokoelma neuroneja, jotka sijaitsevat tietyllä keskushermoston tasolla ja osallistuvat tämän refleksin muodostumiseen. Refleksikeskuksen tehtävänä on analysoida ja syntetisoida tietoa sekä vaihtaa tietoa afferentista efferentille.
Riippuen hermoston osastosta (somaattinen ja autonominen), refleksit, joiden keskus sijaitsee selkäytimessä, eroavat interkalaaristen hermosolujen sijainnista. Joten somaattisen hermoston kohdalla refleksikeskus sijaitsee selkäytimen etu- ja takasarvien välissä. Autonomisen hermoston refleksikeskus (kallienvälisten hermosolujen ruumiit) sijaitsee takasarvissa. Hermoston somaattiset ja autonomiset osat eroavat myös efferenttien hermosolujen sijainnista. Somaattisen hermoston motoristen hermosolujen ruumiit sijaitsevat selkäytimen etusarvissa, autonomisen järjestelmän preganglionisten hermosolujen ruumiit sijaitsevat keskisarvien tasolla.
Molempien solutyyppien aksonit muodostavat refleksikaaren efferentin polun. Somaattisessa hermostossa se on jatkuvaa, se koostuu kuiduista tyyppi A-b. Ainoat poikkeukset ovat A-g-säikeet, jotka johtavat virityksen selkäytimen soluista lihaskarojen intrafusaalisiin kuituihin. Autonomisen hermoston efferenttipolku katkeaa autonomisessa gangliossa, joka sijaitsee joko intramuraalisesti (parasympaattinen osa) tai lähellä selkäydintä (erikseen tai sympaattiseen runkoon - sympaattinen osa). Preganglioninen kuitu kuuluu B-kuituihin, postganglioninen kuitu kuuluu C-ryhmään.
Hermoston somaattisen osan työelin on poikkijuovainen luurankolihas, vegetatiivisessa kaaressa efektori on rauhanen tai lihas (sileä tai poikkijuovainen sydän). Efferentin reitin ja työelimen välillä on kemiallinen myoneuraalinen tai neurosekretiivinen synapsi.
Refleksikaari sulkeutuu renkaaksi käänteisen afferentaation takia - impulssien virtaus efektorireseptoreista takaisin refleksikeskukseen. Palautetoiminto - signalointi keskushermostolle suoritetusta toiminnosta. Jos sitä ei suoriteta tarpeeksi, hermokeskus innostuu - refleksi jatkuu. Myös käänteisen afferentaation ansiosta keskushermoston perifeeristä toimintaa ohjataan.
Erota negatiivinen ja positiivinen palaute. Ensimmäinen, suorittaessaan tiettyä toimintoa, käynnistää mekanismin, joka estää tämän toiminnon. Positiivinen palaute koostuu jo suoritettavan toiminnon lisästimulaatiosta tai jo masentuneen toiminnon estämisestä. Positiivinen käänteinen afferentaatio on harvinaista, koska se saattaa biologisen järjestelmän epävakaaseen asentoon.
Yksinkertaiset (monosynaptiset) refleksikaarit koostuvat vain kahdesta neuronista (afferentti ja efferentti) ja eroavat toisistaan vain proprioseptiivisissä reflekseissä. Loput kaaret sisältävät kaikki edellä mainitut komponentit.
Hermosäikeiden fysiologiset ominaisuudet ja toiminnallinen merkitys
Hermokuiduilla on suurin virittyvyys, suurin virityksen johtumisnopeus, lyhin tulenkestävä ajanjakso ja korkea labilisuus. Tämä tarjotaan korkeatasoinen aineenvaihduntaprosessit ja alhainen kalvopotentiaali.
Tehtävä: hermoimpulssien johtaminen reseptoreista keskushermostoon ja päinvastoin.
Hermosäikeiden rakenteelliset ominaisuudet ja tyypit
Hermosäike - aksoni - on peitetty solukalvolla.
Hermokuituja on 2 tyyppiä:
Myelinisoimattomat hermosäikeet - yksi kerros Schwann-soluja, niiden välissä - rakomaisia tiloja. solukalvo koko yhteydenpidon ajan ympäristöön. Kun ärsytystä käytetään, ärsykkeen vaikutuskohdassa tapahtuu viritystä. Myelinisoimattomilla hermokuiduilla on kaikkialla sähkögeenisiä ominaisuuksia (kyky tuottaa hermoimpulsseja).
Myelinoidut hermosäikeet - peitetty Schwann-solukerroksilla, jotka paikoin muodostavat Ranvier-solmukkeita (alueet, joissa ei ole myeliiniä) 1 mm:n välein. Ranvierin sieppauksen kesto on 1 µm. Myeliinivaippa suorittaa troofisia ja eristäviä toimintoja (korkea vastus). Myeliinin peittämillä alueilla ei ole sähkögeenisiä ominaisuuksia. Heillä on Ranvierin sieppaukset. Herätys tapahtuu Ranvierin sieppauksessa lähimpänä ärsykkeen vaikutuspaikkaa. Ranvierin sieppauksissa korkea tiheys Na-kanavat, joten jokaisessa Ranvierin sieppauksessa hermoimpulssien lisääntyminen tapahtuu.
Ranvierin sieppaukset toimivat toistimina (tuottavat ja vahvistavat hermoimpulsseja).
Hermokuitua pitkin tapahtuvan virityksen johtamismekanismi
1885 - L. German - pyöreät virrat syntyvät hermosäikeen virittyneen ja virittymättömän osion välillä.
Ärsyttävän aineen vaikutuksesta kudoksen ulko- ja sisäpinnan (alueet, joilla on erilaisia varauksia) välillä on potentiaaliero. Näiden alueiden välillä syntyy sähkövirta (Na + -ionien liike). Hermokuidun sisällä syntyy virta positiivisesta napasta negatiiviseen napaan, eli virta suuntautuu virittyneeltä alueelta virittymättömälle. Tämä virta poistuu virittymättömän alueen läpi ja saa sen latautumaan. Hermosäidun ulkopinnalla virta kulkee virittymättömältä alueelta virittyneelle alueelle. Tämä virta ei muuta viritetyn alueen tilaa, koska se on tulenkestävässä tilassa.
Todiste pyöreän virtauksen olemassaolosta: hermosäike asetetaan sisään NaCl-liuos ja rekisteröi virityksen nopeus. Sitten hermokuitu asetetaan öljyyn (resistanssi kasvaa) - johtumisnopeus laskee 30%. Sen jälkeen hermosäike jätetään ilmaan - viritysnopeus pienenee 50%.
Myelinisoituneiden ja myelinisoitumattomien hermosäikeiden kautta tapahtuvan virityksen johtamisen ominaisuudet:
myeliinikuidut - niillä on korkea vastus, sähkögeeniset ominaisuudet vain Ranvierin solmuissa. Ärsykkeen vaikutuksesta viritys tapahtuu Ranvierin lähimmässä leikkauspisteessä. Naapurin sieppaus polarisaatiotilassa. Tuloksena oleva virta aiheuttaa viereisen leikkauspisteen depolarisaation. Ranvierin solmuissa on korkea Na-kanavien tiheys, joten jokaisessa seuraavassa solmussa syntyy hieman suurempi (amplitudi) toimintapotentiaali, jonka vuoksi viritys etenee ilman vähennystä ja voi hypätä useiden solmujen yli. Tämä on Tasakin suolainen teoria. Todiste teorialle on, että hermosäikeeseen ruiskutettiin lääkkeitä, jotka estävät useita sieppauksia, mutta sen jälkeen tallennettiin virityksen johtuminen. Tämä on erittäin luotettava ja kannattava menetelmä, koska pienet vauriot eliminoidaan, viritysnopeus kasvaa ja energiakustannukset pienenevät;
myelinoitumattomat kuidut - pinnalla on sähkögeenisiä ominaisuuksia kauttaaltaan. Siksi pieniä pyöreitä virtoja esiintyy muutaman mikrometrin etäisyydellä. Viritys on jatkuvasti kulkevan aallon muotoa.
Tämä menetelmä on vähemmän kannattava: korkeat energiakustannukset (Na-K-pumpun toiminnalle), alhaisempi viritysnopeus.
Hermosäikeiden luokittelu
Hermosäidut luokitellaan seuraavasti:
toimintapotentiaalin kesto;
kuidun rakenne (halkaisija);
virityksen nopeus.
Seuraavat hermokuitujen ryhmät erotetaan:
ryhmä A (alfa, beeta, gamma, delta) - lyhin toimintapotentiaali, paksuin myeliinivaippa, suurin viritysnopeus;
ryhmä B - myeliinivaippa on vähemmän korostunut;
Ryhmä C - ei myeliinivaippaa.
Dendriittien ja aksonien morfologiset erot
1. Yksittäisessä neuronissa on useita dendriittejä, aksoni on aina yksi.
2. Dendriitit ovat aina lyhyempiä kuin aksoni. Jos dendriittien koko ei ylitä 1,5-2 mm, aksonit voivat olla 1 m tai enemmän.
3. Dendriitit siirtyvät tasaisesti pois solurungosta ja niillä on vähitellen vakiohalkaisija huomattavan matkan ajan.
4. Dendriitit haarautuvat yleensä alle terävä kulma, ja oksat on suunnattu poispäin solusta. Aksonit luovuttavat kollateraalit useimmiten suorassa kulmassa, kollateraalien orientaatio ei liity suoraan solurungon sijaintiin.
5. Dendriittien haarautumiskuvio samantyyppisissä soluissa on vakiompi kuin näiden solujen aksonin haarautuminen.
6. Kypsien hermosolujen dendriitit on peitetty dendriittipiikillä, jotka puuttuvat somasta ja dendriittirunkojen alkuosista. Aksoneilla ei ole piikkiä.
7. Dendriiteillä ei koskaan ole pulpista kuorta. Aksoneja ympäröi usein myeliini.
8. Dendriiteillä on säännöllisempi tilaorganisaatio mikrotubuluksista, aksoneja hallitsevat neurofilamentit ja mikrotubulukset ovat vähemmän järjestyneitä
9. Dendriiteissä, erityisesti niiden proksimaalisissa osissa, on endoplasmista retikulumia ja ribosomeja, jotka eivät ole aksoneissa.
10. Dendriittien pinta on useimmissa tapauksissa kosketuksessa synoptisten plakkien kanssa ja siinä on aktiivisia vyöhykkeitä, joilla on postsynaptinen erikoistuminen.
Dendriittien rakenne
Jos dendriittien geometriasta, niiden oksien pituudesta, suunnasta on suhteellisen paljon kirjallisuutta, niin n. sisäinen rakenne, sytoplasman yksittäisten komponenttien rakenteesta, on vain erillistä hajatietoa. Tämä tieto tuli mahdolliseksi vasta, kun elektronimikroskooppiset tutkimukset otettiin käyttöön neurohistologiassa.
Dendriitin tärkeimmät ominaispiirteet, jotka erottavat sen elektronimikroskooppisilla leikkeillä:
1) myeliinivaipan puute,
oikean mikrotubulusjärjestelmän läsnäolo,
3) aktiivisten synapsien vyöhykkeiden läsnäolo niissä, joilla on selvästi ilmaistu dendriitin sytoplasman elektronitiheys,
4) poistuminen piikien dendriitin yhteisestä rungosta,
5) haarasolmujen erityisesti järjestetyt vyöhykkeet,
6) ribosomien sisällyttäminen,
7) rakeisen ja ei-rakeisen endoplasmisen retikulumin läsnäolo proksimaalisilla alueilla.
Dendriittisen sytoplasman merkittävin piirre on lukuisten mikrotubulusten läsnäolo. Ne tunnistetaan hyvin sekä poikittais- että pitkittäisleikkauksilla. Dendriitin proksimaalisesta osasta alkaen mikrotubulukset kulkevat yhdensuuntaisesti dendriitin pitkän akselin kanssa sen distaalisiin haaroihin. Mikrotubulukset seuraavat dendriitissä rinnakkain toistensa kanssa yhdistämättä tai leikkaamatta toisiaan. Poikkileikkauksissa voidaan nähdä, että yksittäisten putkien väliset etäisyydet ovat vakioita. Yksittäiset dendriittitiehyet ulottuvat melko pitkiä matkoja noudattaen usein käyriä, jotka voivat olla dendriittien kulkua pitkin. Tubulusten lukumäärä on suhteellisen vakio dendriitin poikkileikkauksen pinta-alayksikköä kohti ja on noin 100 per 1 µm. Tämä luku on tyypillinen kaikille dendriiteille, jotka on otettu eri osastoja keskus- ja ääreishermosto, eri tyyppejä eläimet.
Mikrotubulusten tehtävänä on aineiden kuljettaminen hermosolujen prosesseja pitkin.
Kun mikrotubulukset tuhoutuvat, aineiden kuljetus dendriitissä voi häiriintyä, jolloin prosessien pääteosat eivät pääse virtaamaan ravinteita ja energia-aineet solurungosta. Dendriitit, ylläpitääkseen synaptisten kontaktien rakennetta äärimmäisissä olosuhteissa ja varmistaakseen siten hermosolujen välisen vuorovaikutuksen toiminnan, korvaavat puutteen ravinteita johtuen niiden viereisistä rakenteista (synaptiset plakit, pehmeiden kuitujen monikerroksinen myeliinivaippa sekä gliasolujen fragmentit).
Jos patogeenisen tekijän vaikutus eliminoidaan ajoissa, dendriitit palauttavat mikrotubulusten rakenteen ja oikean tilaorganisaation, mikä palauttaa aineen kuljetusjärjestelmän, joka on luontainen normaaleille aivoille. Jos patogeenisen tekijän vahvuus ja kesto ovat merkittäviä, endosytoosi-ilmiöt voivat adaptiivisen funktionsa sijaan tulla kohtalokkaaksi dendriiteille, koska fagosytoosiin muodostuneita fragmentteja ei voida hyödyntää ja ne kerääntyvät dendriittien sytoplasmaan, johtavat sen peruuttamattomaan tilaan. vahingoittaa.
Mikrotubulusten järjestäytymisen rikkominen johtaa voimakkaaseen muutokseen eläinten käyttäytymisessä. Eläimillä, joissa dendriiteissä olevat mikrotubulukset tuhoutuivat kokeessa, havaittiin monimutkaisten käyttäytymismuotojen hajoamista, kun taas yksinkertaiset ehdolliset refleksit säilyivät. Ihmisillä tämä voi johtaa vakaviin häiriöihin korkeamman hermoston toiminnassa.
Se tosiasia, että dendriitit ovat herkin paikka patologisen tekijän vaikutukselle mielisairaus, jotkut amerikkalaisten tutkijoiden teokset todistavat. Kävi ilmi, että seniilissä dementiassa (syanoottinen dementia) ja Alzheimerin taudissa Golgi-menetelmällä käsitellyt aivovalmisteet eivät paljasta hermosolujen prosesseja. Dendriittien rungot näyttävät olevan palaneet ja hiiltyneet. Näiden prosessien havaitsematta jättäminen aivojen histologisissa valmisteissa liittyy todennäköisesti myös mikrotubulusten ja neurofilamenttien järjestelmän rikkomiseen näissä prosesseissa.
Löytyy dendriiteistä. Ne seuraavat yhdensuuntaisesti dendriitin pitkän akselin kanssa, ne voivat sijaita erikseen tai kerätä nippuihin, mutta ne eivät sijaitse tiukasti sytoplasmassa. Todennäköisesti ne voivat yhdessä mikrotubulusten kanssa olla neurofibrillejä.
Kaikille keskushermoston dendriiteille on ominaista pinnan kasvu, joka johtuu useista dikotomisista jakautumisista. Tässä tapauksessa jakovyöhykkeille muodostetaan erityisiä laajennuspaikkoja tai haarasolmuja.
Normaalianalyysi osoittaa, että haarasolmussa, johon kaksi dendriittihaaraa lähestyy, kukin omaa signaaliaan kuljettaen, voidaan suorittaa seuraavat toiminnot. Haarasolmun kautta yhteinen runko ja edelleen hermosolun kehoon:
tai signaali yhdestä haarasta,
tai vain toisesta
tai kahden signaalin vuorovaikutuksen tulos,
tai signaalit kumoavat toisensa.
Haarasolmun sytoplasma sisältää melkein kaikki keholle ominaiset komponentit hermosolu, ja kohdat eroavat rakenteeltaan jyrkästi jakamalla saadun yhteisen dendriittirungon ja oksien sytoplasmasta. Haarasolmuissa on lisääntynyt määrä mitokondrioita, rakeinen ja sileä verkkokalvo, yksittäisten ribosomien klusterit ja ruusukeiksi kootut ribosomit ovat näkyvissä. Nämä komponentit (rakeinen ja sileä verkkokalvo, ribosomit) osallistuvat suoraan proteiinisynteesiin. Mitokondrioiden kerääntyminen näihin paikkoihin osoittaa oksidatiivisten prosessien intensiteetin.
Dendriittien toiminnot
Haluan huomauttaa, että suurimmat vaikeudet, joita tutkija kohtaa tutkiessaan dendriittien toimintaa, on tiedon puute dendriittikalvon ominaisuuksista (toisin kuin hermosolun kalvon), joka johtuu siitä, että mikroelektrodia ei ole mahdollista viedä. dendriittiin.
Arvioimalla dendriittien yleisgeometriaa, synapsien jakautumista ja sytoplasman erityistä rakennetta dendriittihaaroittumispaikoilla voidaan puhua erityisistä hermosolulokuksista, joilla on oma toimintansa. Yksinkertaisin asia, joka voidaan katsoa haaroittumiskohtien dendriittisiksi kohtiksi, on troofinen toiminto.
Edellä olevasta seuraa, että dendriittien sytoplasma sisältää paljon ultraa rakenneosat pystyvät suorittamaan tärkeät tehtävänsä. Dendriitissä on tiettyjä lokuksia, joissa sen työllä on omat ominaisuutensa.
Hermosolun lukuisten dendriittihaarojen päätarkoitus on muodostaa yhteys muiden hermosolujen kanssa. nisäkkäiden aivokuoressa iso osuus aksodendriset yhteydet osuvat kosketuksiin erikoistuneiden dendriittien - dendriittipiikkien - kanssa. Dendriittikärjet ovat fylogeneettisesti hermoston nuorimpia muodostumia. Ontogeniassa ne kypsyvät paljon myöhemmin kuin muut hermorakenteet ja edustavat hermosolun plastisinta laitetta.
Yleensä dendriittiselkärangalla on tyypillinen muoto nisäkkäiden aivokuoressa. (Kuva 2). Päädendriittirungosta lähtee suhteellisen kapea varsi, joka päättyy jatkeeseen - päähän. On todennäköistä, että tämä dendriittisen lisäosan muoto (pään läsnäolo) liittyy toisaalta synaptisen kosketuksen alueen kasvuun aksonipäätteen kanssa, ja toisaalta se palvelee mahtuu erikoistuneita organelleja selkärangan sisälle, erityisesti piikkilaitteistoon, jota esiintyy vain nisäkkään aivokuoren dendriittikärjeissä. Tässä suhteessa analogia synaptisen aksonin päätteen muodon kanssa, kun ohut preterminaalinen kuitu muodostaa jatkeen, näyttää sopivalta. Tämä laajeneminen (synaptinen plakki) muodostaa laajan kosketuksen hermotun substraatin kanssa ja sisältää sisällä suuren joukon ultrarakenteisia komponentteja (synaptisia rakkuloita, mitokondrioita, neurofilamentteja, glykogeenirakeita).
On olemassa hypoteesi (jonka jakaa ja kehittänyt erityisesti Nobel-palkittu F. Crick), että selkärangan geometria voi muuttua aivojen toiminnallisesta tilasta riippuen. Tässä tapauksessa selkärangan kapea kaula voi laajentua ja itse selkäranka litistyy, mikä lisää akso-selkärangan kontaktin tehokkuutta.
Jos dendriittisten piikien muoto ja koko nisäkkäiden aivokuoressa voivat vaihdella jonkin verran, niin vakiona niissä on tietyn selkärangan läsnäolo. Se on toisiinsa yhdistettyjen tubulusten (säiliö) kompleksi, joka sijaitsee yleensä selkärangan päässä. Todennäköisesti tämä organelli liittyy erittäin tärkeisiin toimintoihin, jotka ovat ominaisia fylogeneettisesti nuorimmille aivomuodostelmille, koska piikkilaitteisto löytyy pääasiassa aivokuoresta ja vain korkeammista eläimistä.
Kaikesta huolimatta selkäranka on dendriitin johdannainen, siitä puuttuu neurofilamentteja ja dendriittitiehyitä, sen sytoplasmassa on karkea- tai hienorakeinen matriisi. Yksi vielä ominaisuus Selkäranka aivokuoressa on pakollinen synaptisten kontaktien läsnäolo, joissa on aksonipäätteitä. Selkärangan sytoplasmassa on erityisiä komponentteja, jotka erottavat sen dendriittivarresta. Selkärangan sytoplasmassa on mahdollista havaita erikoinen kolmikko: aktiivisten vyöhykkeiden subsynaptinen erikoistuminen - piikkilaitteisto - mitokondriot. Ottaen huomioon mitokondrioiden suorittamat monimutkaiset ja tärkeät toiminnot, voidaan myös odottaa monimutkaisia toiminnallisia ilmenemismuotoja "triadeissa" synaptisen siirron aikana. Voidaan sanoa, että dendriittiselkärangan sytoplasma ja piikkinen laite voivat olla suoraan yhteydessä synaptiseen toimintaan.
Dendriittikärjet ja dendriittien päät ovat myös erittäin herkkiä äärimmäisille tekijöille. Kaikentyyppisten myrkytysten yhteydessä (esimerkiksi alkoholi, hypoksinen, raskasmetallit - lyijy, elohopea jne.) Aivokuoren solujen dendriiteistä löydettyjen piikien lukumäärä muuttuu. Todennäköisesti piikit eivät katoa, mutta niiden sytoplasmiset komponentit häiriintyvät ja ne ovat huonommin kyllästyneitä suoloilla. raskasmetallit. Koska selkärangat ovat yksi hermosolujen välisten kontaktien rakenneosista, niiden toimintahäiriöt johtavat vakavaan aivojen toiminnan heikkenemiseen.
Joissakin tapauksissa äärimmäisen tekijän lyhytaikaisella vaikutuksella voi ensi silmäyksellä syntyä paradoraalinen tilanne, kun aivosolujen dendriiteistä löydettyjen piikien määrä ei vähene, vaan kasvaa. Joten tämä havaitaan kokeellisen aivoiskemian aikana sen alkuvaiheessa. Samanaikaisesti tunnistettujen piikien määrän lisääntymisen kanssa toimiva tila aivot. Tässä tapauksessa hypoksia on tekijä, joka edistää lisääntynyttä aineenvaihduntaa hermokudoksessa, parempaan varastointiin, jota ei käytetä normaali ympäristö ja kehoon kerääntyneiden toksiinien nopea palaminen. Ultrarakenteellisesti tämä ilmenee selkärangan sytoplasman intensiivisemmällä kehityksellä, selkärangan vesisäiliöiden kasvulla ja laajentumisella. Todennäköisesti tämä ilmiö hypoksian positiivisesta vaikutuksesta havaitaan, kun henkilö kokee suuria fyysinen harjoitus hypoksian olosuhteissa valloittaa vuorenhuiput. Nämä vaikeudet kompensoidaan sitten intensiivisemmällä tuottavalla työllä sekä aivoissa että muissa elimissä.
Dendriittien muodostuminen
Dendriitit ja niiden neuronaaliset yhteydet muodostuvat aivojen ontogeneettisen kehityksen aikana. Lisäksi dendriitit, erityisesti apikaalit, jäävät nuorilla yksilöillä vapaiksi jonkin aikaa muodostamaan uusia kontakteja. Solurunkoa lähempänä sijaitsevat dendriitin osat liittyvät todennäköisesti vahvempiin ja yksinkertaisempiin luonnollisiin ehdollisiin reflekseihin ja päät jätetään uusien yhteyksien ja assosiaatioiden muodostumiseen.
Aikuisena dendriiteissä ei ole enää hermosolujen välisistä kontakteista vapaita alueita, mutta ikääntymisen myötä dendriitin päät kärsivät ensin ja kontaktien kyllästymisen kannalta.
vanhoissa yksilöissä ne muistuttavat dendriittejä lapsuus. Tämä johtuu sekä kuljetusproteiinien syntetisointiprosessien heikkenemisestä solussa että aivojen heikentyneestä verenkierrosta. Ehkä juuri tässä piilee morfologinen perusta sellaiselle neurologiassa ja in jokapäiväinen elämä se, että kun vanhojen ihmisten on vaikea oppia uutta, he usein unohtavat ajankohtaiset tapahtumat ja muistavat menneen hyvin. Sama havaitaan myrkytyksen yhteydessä.
Kuten jo todettiin, dendriittipuun lisääntyminen ja komplikaatio fylogiassa on välttämätöntä paitsi havainnolle suuri numero saapuvia pulsseja, mutta myös esikäsittelyä varten.
Keskushermoston neuronien dendriiteillä on kauttaaltaan synaptinen toiminta, eivätkä pääteosat ole tässä millään tavalla keskimmäisiä huonompia. Jos me puhumme apikaalisten dendriittien distaalisista (terminaalisista) osista pyramidaaliset neuronit aivokuoreen, niin niiden osuus interneuronaalisten vuorovaikutusten toteuttamisessa on jopa merkittävämpi kuin proksimaalisten. Sinne lisää itse rungossa ja apikaalisen dendriitin oksissa olevat terminaaliset synaptiset plakit yhdistyvät useammilla kontakteilla dendriittikärkien päällä.
Tutkiessaan tätä ongelmaa elektronimikroskopian avulla, tutkijat vakuuttuivat myös siitä, että dendriittien päätyosat ovat tiiviisti synaptisten plakkien peitossa ja ovat siten suoraan mukana hermosolujen välisissä vuorovaikutuksissa. elektronimikroskopia osoitti myös, että dendriitit voivat muodostaa kontakteja keskenään. Nämä kontaktit voivat olla joko rinnakkaisia, joille useimmat kirjoittajat antavat sähkötonisia ominaisuuksia, tai tyypillisiä epäsymmetrisiä synapseja, joissa on hyvin määriteltyjä organelleja, jotka tarjoavat kemiallisen siirtymisen. Tällaiset dendro-dendriittiset kontaktit ovat vasta alkamassa herättää tutkijoiden huomion. Joten dendriitti koko pituudeltaan suorittaa synaptista toimintoa. Kuinka dendriitin pinta on mukautettu tarjoamaan kontakteja aksonipäätteiden kanssa?
Dendriitin pintakalvo on suunniteltu käytettäväksi maksimaalisesti hermosolujen välisissä kontakteissa. Koko dendriitti on kuoppainen syvennyksillä, taitoksilla, taskuilla, siinä on erilaisia epäsäännöllisyyksiä, kuten mikrokasvustoa, piikkejä, sienimäisiä lisäyksiä jne. Kaikki nämä dendriittirunkojen kohokuviot vastaavat sisään tulevien synaptisten päätteiden muotoa ja kokoa. Ja sisään eri osastoja hermostoon ja eri eläimillä dendriittipinnan kohokuviolla on erityispiirteitä. Tietenkin merkittävin dendriittikalvon kasvu on dendriittiselkä.
Dendriitit ovat erittäin herkkiä erilaisten äärimmäisten tekijöiden vaikutukselle. Niiden rikkomukset johtavat moniin sairauksiin, kuten mielenterveyshäiriöihin.
Refleksikaari koostuu:
- reseptorit - havaitsevat ärsytystä.
- herkkä (keskeinen, afferentti) hermosäike, joka välittää virityksen keskustaan
- hermokeskus, jossa tapahtuu viritys vaihtuu sensorisista neuroneista motorisiin neuroneihin
- motorinen (keskipakoinen, efferentti) hermosäike, joka kuljettaa virityksen keskushermostosta työelimeen
- efektori - toimiva elin, joka suorittaa vaikutuksen, reaktion vastauksena reseptorin ärsytykseen.
Reseptorit ja vastaanottavat kentät
Reseptori- solu havaitsee ärsytystä.
vastaanottava kenttä- Tämä on anatominen alue, joka ärsyyntyessään aiheuttaa tämän refleksin.
Primaaristen sensoristen reseptorien vastaanottavat kentät on järjestetty yksinkertaisimmalla tavalla. Esimerkiksi ihon pinnan kosketus- tai nosiseptiivinen vastaanottava kenttä on yksittäisen aistisäikeen haara.
Reseptiivisen kentän eri osissa sijaitsevilla reseptorilla on erilainen herkkyys riittävälle stimulaatiolle. Erittäin herkkä vyöhyke sijaitsee yleensä reseptiivisen kentän keskellä, ja herkkyys laskee lähempänä reseptiivisen kentän reunaa.
Sekundaaristen sensoristen reseptorien vastaanottavat kentät on järjestetty samalla tavalla. Erona on, että afferentin kuidun haarat eivät pääty vapaasti, vaan niillä on synaptiset kontaktit herkkien reseptorisolujen kanssa. Maku-, vestibulaari- ja akustiset reseptiiviset kentät järjestetään tällä tavalla.
päällekkäisiä vastaanottavia kenttiä. Herkän pinnan samaa aluetta (esimerkiksi ihoa tai verkkokalvoa) hermottavat useat herkät hermosäikeitä, jotka haarautumisineen menevät päällekkäin yksittäisten afferenttien hermojen vastaanottavien kenttien kanssa.
Reseptiivisten kenttien päällekkäisyyden vuoksi kehon kokonaisaistipinta kasvaa.
Refleksien luokittelu.
Koulutustyypin mukaan:
Ehdollinen (hankittu) - vastaa nimeen, sylki koirasta valoon.
Ehdoton (synnynnäinen) - vilkkuva nieleminen, polvi.
Sijainnin mukaan reseptorit:
Exteroseptiivinen (iho, näkö, kuulo, haju)
Interoseptiivinen (sisäelinten reseptoreista)
Proprioseptiivinen (lihasten, jänteiden, nivelten reseptoreista)
Efektoreille:
Somaattiset tai motoriset (refleksit luurankolihas);
Vegetatiiviset sisäelimet - ruoansulatus, sydän-, eritys-, eritys- jne.
Biologisen alkuperän mukaan:
Puolustava tai suojaava (vaste kosketuskivun jakautumiseen)
Ruoansulatuskanava (ärsyttävä reseptori suuontelossa.)
Seksuaalinen (hormonit veressä)
Arvioitu (pään käännös, vartalo)
Moottori
Posotoninen (tukea kehon asentoja)
Synapsien lukumäärän mukaan:
Monosynaptinen, jonka kaaret koostuvat afferenteista ja efferenteistä neuroneista (esimerkiksi polvi).
Polysynaptinen, jonka kaaret sisältävät myös yhden tai useamman välihermosolun ja niissä on 2 tai useampia synaptisia kytkimiä. (somaattiset ja vegetatiiviset viittaukset).
Disynaptinen (2 synapsia, 3 neuronia).
Vastauksen luonteen mukaan:
Moottori \ moottori (lihassupistukset)
Eritys (eritysrauhasen eritys)
Vasomotorinen (verisuonten laajeneminen ja kaventuminen)
Sydän (muutos. Sydänlihaksen työ.)
Keston mukaan:
vaiheittainen (nopea) käden poisto
tonisoiva (hidas) asennon ylläpito
Hermokeskuksen sijainnin mukaan:
Selkäydin (SM-neuronit ovat mukana) - Käden vetäminen pois kuumista segmenteistä 2-4, polven nykiminen.
refleksit aivoissa
Bulbar (medulla oblongata) - silmäluomien sulkeminen koskettaessa. sarveiskalvoon.
Mesencephalic (keskellä m) - näön maamerkki.
Väliaivot (väliaivot) - hajuaisti
Kortikaalinen (kuori BP GM) - ehdollinen. viite.
Ominaisuudet hermokeskukset.
1. Herätyksen yksipuolinen eteneminen.
Viritys välittyy afferentista efferenttihermosolulle (syy: synapsin rakenne).
Hidastaa virityksen siirtymistä.
Hoitoaine Monien synapsien esiintyminen riippuu myös ärsyttävän aineen voimakkuudesta (summatuksesta) ja fysikaalisesta tilasta. CNS (väsymys).
3. Summaus vaikutusten summaus, kynnyksen alapuolella olevat ärsykkeet.
Väliaikainen: viite Ed. Imp-sa ei ole vielä ohi, mutta jälki. jo saapunut.
Spatiaalinen: useiden sekoitus. Suvanto Ne ovat pöllöehtoisia. Kuvat. Viite.
Fasilitaatio- ja okkluusiokeskus.
Helpotuksen keskus - tapahtuu optimaalisen ärsykkeen (maksimivaste) vaikutuksesta - ilmestyi. Apukeskus.
Toiminnassa min irr. (alennettu otv. Rektsiya) oli okkluusio.
Assimilaatio ja viritysrytmin muunnos.
Transformaatio - muutos hermoimpulssin taajuudessa, kun se kulkee hermokeskuksen läpi. Taajuutta voidaan lisätä tai vähentää.
Assimilaatio (tanssi, päivittäiset rutiinit)
Seuraus
Viive vasteen lopussa ärsykkeen lakkaamisen jälkeen. Liittyy verenkiertohermoon. Imp. Suljettu Neuronien piirit.
lyhyt aika (sekunnin murto-osat)
pitkä (sekuntia)
Hermokeskusten rytminen toiminta.
Hermoimpulssien taajuuden lisääntyminen tai lasku, joka liittyy synapsin ominaisuuksiin ja hermosolujen integratiiviseen kestoon.
8. Hermokeskusten plastisuus.
Mahdollisuus rakentaa uudelleen kiinteistön toimivuus toimintojen tehokkaampaa säätelyä varten, uusien refleksien toteuttaminen, jotka eivät aiemmin olleet tälle keskukselle ominaisia, tai toiminnan palauttaminen. Synps-kerroksen ytimessä on muutos ostoskeskuksen ytimessä.
Muutokset kiihtyvyydessä kemikaalien vaikutuksen alaisena.
Suuri herkkyys todellisille eroille.
Hermokeskusten väsymys.
Liittyy synapsien suureen väsymykseen. Vähennä tunteita. Reseptorit.
Keskushermoston koordinaatiotoiminnan yleiset periaatteet.
Jarrutus- erityinen ner. prosenttia ilmentyy resp. reaktiot.
Lähentymisperiaate
Konvergenssi on eri afferenttireittejä pitkin tulevien impulssien konvergenssia missä tahansa keskushermosolussa tai hermokeskuksessa.
2 . Konvergenssin periaate liittyy läheisesti periaatteeseen yhteinen lopullinen polku avaa Sherrinkton. Monet eri ärsykkeet voivat herättää saman motorisen hermosolun ja saman motorisen vasteen. Tämä periaate johtuu afferenttien ja efferenttien erisuuruisesta määrästä.
Eron periaate
Tämä on yhden neuronin kosketus monien muiden kanssa.
Säteilytys ja virityksen keskittyminen.
Viritysprosessin leviämistä muihin hermokeskuksiin kutsutaan säteilytys (vaaleissa- yhteen suuntaan , yleistetty- laaja).
Jonkin ajan kuluttua säteilytys korvataan virityskonsentraation ilmiöllä samassa keskushermoston alkupisteessä.
Säteilytysprosessilla on positiivinen (uusien ehdollisten refleksien muodostuminen) ja negatiivinen (herätys- ja estoprosessien välisten hienovaraisten suhteiden rikkominen, mikä johtaa motorisen aktiivisuuden häiriöön) rooli.
Vastavuoroisuuden periaate (hidastaa)
Joidenkin solujen viritys aiheuttaa toisten eston interkalaarisen neuronin kautta.
Hallitseva periaate
Ukhtomsky muotoili dominanssiperiaatteen hermokeskusten toiminnan toimintaperiaatteeksi. termi hallitseva tarkoittaa keskushermoston virityksen hallitsevaa painopistettä, joka määrää kehon nykyisen toiminnan.
Hallitsevan fokuksen periaatteet :
Lisääntynyt hermokeskusten kiihtyvyys;
Herätyksen jatkuvuus ajan myötä;
Kyky summata ulkopuolisia ärsykkeitä;
Inertia (kyky ylläpitää kiihottumista pitkään ärsytysvaikutuksen päättymisen jälkeen); kyky aiheuttaa konjugaattiestoja.

