Reflekss- ķermeņa reakcija uz receptoru kairinājumu, ko veic nervu sistēma.
reflekss loks - ceļš, pa kuru refleksa īstenošanas laikā iziet nervu impulss. Sastāv no 5 daļām:
- receptors - jutīgs veidojums, kas spēj reaģēt uz noteikta veida stimulu;
- jutīgs neirons vada impulsus smadzenēm;
- starpkalārais neirons savieno jutīgos un izpildneironus, atrodas muguras smadzenēs vai smadzenēs;
- izpildvaras (motorais) neirons vada impulsus no smadzenēm uz izpildorgāniem;
- izpildaģentūra- muskuļi (kontrakti), dziedzeri (noslēpumi) utt.
Beznosacījumu refleksi
- klāt no dzimšanas
- dzīves laikā tie nemainās un nepazūd;
- ir vienādi visos vienas sugas organismos;
- pielāgot ķermeni pastāvīgiem apstākļiem.
Nosacīti refleksi
- iegūts dzīves laikā;
- var mainīties vai pazust;
- katram organismam ir savs;
- pielāgot ķermeni mainīgajiem apstākļiem.
Pārbaudes
1) Kura saite kalpo par refleksa loka sākumu
A) starpkalāra neirons
B) jutīgs neirons
B) receptors
D) izpildu neirons
2. Kādā secībā refleksa realizācijā tiek iekļautas refleksa loka sastāvdaļas
A) izpildorgāns, motorais neirons, starpkalārais neirons, sensorais neirons, receptors
B) starpkalārais neirons, sensorais neirons, motorais neirons, receptors, izpildorgāns
C) receptors, sensorais neirons, starpkalārais neirons, motorais neirons, izpildorgāns
D) sensorais neirons, starpkalārais neirons, receptors, izpildorgāns, motorais neirons
3. Refleksa loka sākuma stadija siekalu refleksā
A) siekalu dziedzeris
B) receptors
B) starpkalāra neirons
D) izpildu neirons
4. Stimulu darbība izraisa rašanos nervu impulss V
A) sensorie neironi
B) motorie neironi
B) receptori
D) starpkalārie neironi
5. Uzbudinājums tiek virzīts gar jutīgo neironu
A) uz centrālo nervu sistēmu
B) izpildinstitūcijai
B) uz receptoriem
D) uz muskuļiem
6. Receptori ir jutīgi veidojumi, kas
A) pārraida impulsus centrālajai nervu sistēmai
B) pārraida nervu impulsus no starpkalāru neironiem uz izpildvaru
C) uztvert stimulus un pārveidot stimulu enerģiju nervu uzbudinājuma procesā
D) saņem nervu impulsus no sensorajiem neironiem
7. Nervu impulsi tiek pārraidīti uz smadzenēm caur neironiem
A) motors
B) interkalārs
B) jutīgs
D) izpildvaras
8. Refleksa loka gali
A) izpilddirektors
B) jutīgs neirons
B) receptors
D) starpkalāra neirons
9. Nervu impulsi no receptoriem virzās uz centrālo nervu sistēmu
A) sensorie neironi
B) motorie neironi
C) sensorie un motoriskie neironi
D) starpkalārie un motorie neironi
10. Nervu veidojumi cilvēka organismā, uztverot kairinājumus no ārējās vides, ir
A) nervi
B) receptori
B) smadzenes
D) nervu mezgli
11. Stimulu pārvēršanās nervu impulsā notiek gadā
A) motoriskie nervi
B) smadzenes
B) receptori
D) muguras smadzenes
12. Nervu impulsi tiek pārraidīti no cilvēka maņām caur neironiem
A) jutīgs
B) motors
B) ievietot
D) izpildvaras
13. Uztvert informāciju un pārvērst to nervu impulsos
A) receptori
B) nervu mezgli
B) izpildinstitūcijas
D) starpkalārie neironi
14. Nosacīts reflekss
A) manto pēcnācēji, bet nepaliek visu mūžu
B) organisms dzīves laikā viegli iegūst un zaudē
C) manto pēcnācēji no vecākiem un saglabājas līdz
D) ir pastāvīga ķermeņa reakcija uz stingri noteiktiem vides stimuliem
15. Nosacītā refleksa piemērs kaķim
A) siekalošanās ēšanas laikā
B) ķepas izvilkšana apdeguma laikā
B) reakcija uz segvārdu
D) reakcija uz asu skaņu
16. Siekalošanās cilvēkiem ieraugot citronu - reflekss
A) nosacīti
B) beznosacījuma
B) aizsargājošs
D) indikatīvs
17) Nosacīti refleksi cilvēkiem veidojas procesā
A) individuālā attīstība
B) sugu veidošanās
C) vēsturiskā attīstība
D) embriju veidošanās
18. Nosacīta refleksa piemērs ir
A) siekalošanās, ieraugot sagrieztu citronu
B) siekalošanās barošanas laikā
C) zīlītes sašaurināšanās spēcīgas gaismas ietekmē
D) rokas atvilkšana ar pēkšņu injekciju
19. Bērna reakcija uz piena pudeli ir reflekss, kas
A) ir iedzimta
B) veidojas bez garozas līdzdalības puslodes
B) tiek iegūti dzīves laikā
D) saglabājas visu mūžu
20. Aizsargājošs šķaudīšanas reflekss
A) nav iedzimta
B) ir nosacīts
B) laika gaitā vājinās
D) raksturīga visiem sugas indivīdiem
21. Tiek sauktas cilvēku un dzīvnieku dzīves laikā iegūtās reakcijas, kas nodrošina pielāgošanos mainīgajiem vides apstākļiem
A) nosacīti refleksi
B) instinkti
B) beznosacījumu refleksi
D) refleksi, kas ir iedzimti
22.Beznosacījuma reflekss
A) ir iedzimta
B) iegūts dzīves laikā
C) tiek ražots noteiktiem signāliem
D) ir dažādu ārējo signālu pamatā
23. Cilvēku un dzīvnieku beznosacījumu refleksi nodrošina
C) jaunu motoriku apgūšana ar ķermeņa palīdzību
24. Cilvēka reakcija uz zaļa krāsa luksofors ir reflekss
A) iedzimts
B) iegūts
B) beznosacījuma
D) iedzimta
25.Izcelt kuņģa sula reaģējot uz pārtikas stimulēšanas receptoriem mutes dobums
A) uztraukums
B) bremzēšana
B) beznosacījuma reflekss
D) pašregulācija
26. Nosacītu refleksu bioloģiskā nozīme ķermenim ir tāda, ka tie
A) uzturēt iekšējās vides noturību
B) sugai raksturīgs un saglabājas visu mūžu
B) veicināt mijiedarbību funkcionālās sistēmas
D) nodrošināt pielāgošanos mainīgajiem vides apstākļiem
27. Dzīvnieki dzīves procesā iegūst refleksus
A) beznosacījuma
B) iedzimta
C) raksturīga visiem noteiktas sugas indivīdiem
D) ļaujot viņiem izdzīvot mainīgos apstākļos
28. Nosacīts reflekss cilvēkiem ir
A) zīlītes sašaurināšanās spilgtā gaismā
B) pagriežot galvu uz asu skaņu
C) siekalu sekrēcija, kad ēdiens nonāk mutē
D) reakcija uz vārda nozīmi
29. Nosacīta refleksa piemērs ir
1) prasme braukt ar velosipēdu
2) siekalošanās ēdot
3) vēlme gulēt, kad noguris
4) šķaudīšana un klepus
30. Cilvēku un dzīvnieku nosacīti refleksi nodrošina
A) organisma pielāgošanās pastāvīgiem vides apstākļiem
B) ķermeņa pielāgošanās mainīgajai ārējai pasaulei
C) jaunu motorisko prasmju organismu attīstība
D) trenera komandu atšķirība starp dzīvniekiem
31.Iedzimtie refleksi
A) ir organisma sugas īpašības
B) raksturīgs atsevišķiem sugas indivīdiem
C) pieprasīt papildu nosacījumus īstenošanai
D) nav iedzimtas
galvenā forma nervu darbība ir reflekss. Reflekss - ķermeņa cēloņsakarības reakcija uz ārējās vai iekšējās vides izmaiņām, ko veic ar obligātu centrālās puses līdzdalību. nervu sistēma reaģējot uz receptoru stimulāciju. Refleksu ietekmē notiek jebkuras ķermeņa darbības rašanās, maiņa vai pārtraukšana.
Tiek saukts nervu ceļš, pa kuru refleksu īstenošanas laikā izplatās ierosme reflekss loks.
Refleksu loki sastāv no piecām sastāvdaļām: 1) receptoru; 2) aferentā nerva ceļš; 3) refleksu centrs; 4) eferentais nervu ceļš; 5) efektors (darba ķermenis).
Receptors- Šis ir jutīgs nervu gals, kas uztver kairinājumu. Receptoros stimula enerģija tiek pārvērsta nervu impulsa enerģijā. Atšķirt: 1) eksteroreceptori- satraukti apkārtējās vides kairinājuma ietekmē (ādas, acu receptori, iekšējā auss, deguna un mutes gļotāda); 2) interoreceptori- uztvert stimulus no ķermeņa iekšējās vides (receptoriem iekšējie orgāni, kuģi); 3) proprioreceptori- reaģēt uz atsevišķu ķermeņa daļu stāvokļa izmaiņām telpā (muskuļu, cīpslu, saišu, locītavu maisiņu receptori).
Aferentais neironu ceļš ko attēlo receptoru neironu procesi, kas veic ierosinājumus uz centrālo nervu sistēmu.
refleksu centrs sastāv no neironu grupas, kas atrodas dažādos centrālās nervu sistēmas līmeņos un pārraida nervu impulsus no aferenta uz eferento nervu ceļu.
eferents nervu ceļš vada nervu impulsus no centrālās nervu sistēmas uz efektoru.
Efektors- izpildorgāns, kura darbība mainās nervu impulsu ietekmē, kas tam nonāk caur refleksa loka veidojumiem. Efektori var būt muskuļi vai dziedzeri.
refleksu loki var būt vienkārša vai sarežģīta. Vienkāršs reflekss loks sastāv no diviem neironiem - uztverošajiem un efektoriem, starp kuriem ir viena sinapse. Šāda divu neironu refleksu loka diagramma ir parādīta attēlā. 71.
Vienkārša refleksa loka piemērs ir cīpslu refleksu loki, piemēram, ceļa skriemelis.
Vairuma refleksu refleksu loki ietver nevis divus, bet lielāku skaitu neironu: receptoru, vienu vai vairākus starpkalārus un efektorus. Šādus refleksu lokus sauc par sarežģītiem, daudzneironiskiem. Sarežģīta (trīs neironu) refleksa loka diagramma ir parādīta attēlā. 72.
Tagad ir noskaidrots, ka efektora reakcijas laikā tiek uzbudināti daudzi darba orgānā esošie nervu gali. Nervu impulsi tagad no efektora atkal nonāk centrālajā nervu sistēmā un informē to par pareizu darba orgāna reakciju. Tādējādi refleksu loki nav atvērti, bet gan gredzenu veidojumi.
Refleksi ir ļoti dažādi. Tos var klasificēt pēc vairākiem kritērijiem: 1) bioloģiskā nozīme(pārtika, aizsardzība, seksuāls); 2) atkarībā no kairināto receptoru veida: eksteroceptīvie, interoceptīvie un proprioceptīvie; 3) pēc reakcijas rakstura: motors vai motors (izpildorgāns - muskuļi), sekrēcijas (efektors - dzelzs), vazomotors (asinsvadu sašaurināšanās vai paplašināšanās).
Visus visa organisma refleksus var iedalīt divos lielas grupas: beznosacījuma un nosacījuma. To atšķirības tiks aplūkotas XII nodaļā.
Nervu sistēmas darbībai ir reflekss raksturs. Pat XVII gadsimtā. Franču filozofs un matemātiķis Renē Dekarts aprakstīja refleksu aktu. Viņš atzīmēja ķermeņa reakciju uz kairinājumu un ierosināja, ka pastāv ceļš, pa kuru iet nervu uzbudinājums. Pašu terminu "reflekss" vēlāk - 18. gadsimtā - izvirzīja čehu zinātnieks J. Pročazka (no latīņu valodas "reflekss" - atspoguļota darbība). Vēlāk I. M. Sečenovs darbā “Smadzeņu refleksi” pierādīja, ka nervu sistēmas reakcija uz dažāda veida kairinājumi notiek saskaņā ar refleksu mehānismu, t.i. visas apzinātās un neapzinātās darbības ir refleksīvas izcelsmes. Reflekss tiek saprasts kā specifiska ķermeņa reakcija uz iekšējās vides kairinātāju vai saņemta no ārpuses ar obligātu centrālās nervu sistēmas līdzdalību. Refleksus parasti sauc par nervu darbības funkcionālajām vienībām.
Refleksa loka refleksu mehānismi
Refleksus var klasificēt pēc dažādiem kritērijiem. Tātad, atkarībā no loka slēgšanas līmeņa, t.i. atkarībā no refleksu centra atrašanās vietas refleksus iedala mugurkaula (reflekss aizveras muguras smadzenēs), bulbar (refleksa centrs - medulla), mezenencefālisko (refleksa loka struktūra ir slēgta vidussmadzenēs), diencefāliski un kortikālie refleksu centri atrodas telencefalons un smadzeņu garozā, attiecīgi. Pēc efektora pazīmes tie ir somatiski, kad refleksa eferentais ceļš nodrošina skeleta muskuļu motoro inervāciju, un veģetatīvi, kad efektori ir iekšējie orgāni. Atkarībā no kairināto receptoru veida refleksus iedala eksteroceptīvajos (ja receptors uztver informāciju no ārējās vides), proprioceptīvajos (refleksa loks sākas no muskuļu un skeleta aparāta receptoriem) un interoceptīvajos (no iekšējo orgānu receptoriem). Interoceptīvie refleksi savukārt tiek iedalīti viscero-viscerālajos (refleksa loks savieno divus iekšējos orgānus), viscero-muskuļajos (receptori atrodas uz muskuļu-cīpslu aparāta, efektors ir iekšējais orgāns) un viscero-kutānos (receptori lokalizēti ādā, darba orgāni ir iekšpuses). Pēc Pavlova domām, refleksus iedala kondicionētajos (dzīves laikā izstrādātos, katram indivīdam raksturīgos) un beznosacījumu (iedzimtos, sugai raksturīgos: pārtikas, seksuālos, aizsardzības-motoros, homeostatiskie utt.).
Kā darbojas reflekss loks?
Neatkarīgi no refleksa veida tā refleksa loks satur receptoru, aferento ceļu, nervu centru, eferento ceļu, darba orgānu un atgriezenisko saiti. Izņēmums ir aksona refleksi, kuru refleksa loka struktūra atrodas vienā neironā: maņu procesi rada centripetālus impulsus, kas, ejot cauri neirona ķermenim, pa aksonu izplatās uz centrālo nervu sistēmu, un pa aksona atzaru impulsi sasniedz efektoru. Šādi refleksi tiek attiecināti uz metasimpatiskās nervu sistēmas darbību, caur tiem, piemēram, tiek veikti mehānismi asinsvadu tonusa un ādas dziedzeru darbības regulēšanai.
Kairinājuma uztveršanas un pārvēršanas ierosmes enerģijā funkciju veic refleksu loku receptori. Uzbudinājuma receptoru enerģijai ir lokālas reakcijas raksturs, kas ir svarīgs ierosmes gradācijā pēc spēka.
Pamatojoties uz receptoru uzbūvi un izcelsmi, tos var iedalīt primārajos sensorajos, sekundārajos sensorajos un brīvajos nervu galos. Pirmajā neirons pats darbojas kā receptors (tas attīstās no neiroepitēlija); starp stimulu un pirmo aferento neironu nav starpstruktūru. Primāro sensoro receptoru lokālā reakcija – receptoru potenciāls – arī ir ģeneratora potenciāls, t.i. izraisot darbības potenciālu pāri aferentās šķiedras membrānai. Primārie sensorie receptori ir redzes, ožas, ķīmij- un baroreceptori. sirds un asinsvadu sistēmu.
Sekundārās sensorās šūnas ir īpašas nenervu izcelsmes struktūras, kas mijiedarbojas ar pseidounipolāru sensoro šūnu dendritiem ar sinaptisko neiroreceptoru kontaktu palīdzību. Receptoru potenciāls, kas rodas stimula iedarbībā sekundārās sensorās šūnās, nav ģenerators un neizraisa darbības potenciāla parādīšanos uz aferentās šķiedras membrānas. Uzbudinošais postsinaptiskais potenciāls rodas tikai caur receptoru šūnu mediatora atbrīvošanas mehānismu. Stimulēja stipruma gradācija tiek veikta, izvadot dažādus mediatora daudzumus (jo vairāk tiek atbrīvots mediators, jo spēcīgāks ir stimuls).
Sekundārās sensorās šūnas ietver dzirdes, vestibulārās, miega, taustes un citus receptorus. Dažkārt funkcionēšanas īpatnību dēļ šajā grupā ietilpst fotoreceptori, kas no anatomiskā viedokļa un to izcelsmes dēļ no neiroepitēlija ir sekundāri sensējoši.
Brīvie nervu gali ir pseido-unipolāru sensoro šūnu zarojoši dendriti un ir lokalizēti gandrīz visos cilvēka ķermeņa audos.
Atkarībā no stimula enerģētiskā rakstura, uz kuru receptors reaģē, tos iedala mehānoreceptoros (taustes, baroreceptoros, tilpuma receptoros, dzirdes, vestibulārajos; tie parasti uztver mehānisko kairinājumu ar šūnu izaugumu palīdzību), ķīmijreceptoros (ožas), ķīmijreceptoros (centrālo nervu sistēmu uztverošajos, foto-receptoros un centrālo nervu sistēmu uztverošajos, fotoreceptoros). šūnu izaugumi), termoreceptori (reaģē uz izmaiņām "siltums-aukstums" - Rufini ķermeņi un Krause kolbas gļotādas) un nociceptori (nekapsulēti sāpju gali).
Refleksa loka pēcreceptoru veidojumi
Refleksu loku struktūras pēcreceptoru veidošanās ir aferents ceļš, ko veido pseido-unipolārs sensorais neirons, kura ķermenis atrodas mugurkaula ganglijs, un veidojas aksoni muguras saknes muguras smadzenes. Aferentā ceļa funkcija ir novadīt informāciju uz centrālo saiti, turklāt šajā posmā informācija tiek kodēta. Šiem nolūkiem mugurkaulnieku ķermenī tiek izmantots binārs kods, ko veido impulsu pārrāvumi (zalves) un spraugas starp tiem. Ir divi galvenie kodēšanas veidi: frekvence un telpiskā.
Pirmais ir dažāda impulsu skaita veidošanās sprādzienā, atšķirīgs uzliesmojumu skaits, to ilgums un pārtraukumu ilgums starp tiem atkarībā no receptoram pielietotās stimulācijas stipruma. Telpiskā kodēšana veic stimula stipruma gradāciju, iesaistot atšķirīgu nervu šķiedru skaitu, pa kurām vienlaikus tiek veikta ierosme.
Aferentais ceļš galvenokārt sastāv no A-α, A-β un A-δ šķiedrām.
Izejot cauri šķiedrām, nervu impulss nonāk refleksu centrā, kas anatomiskā nozīmē ir neironu kopums, kas atrodas noteiktā centrālās nervu sistēmas līmenī un piedalās šī refleksa veidošanā. Refleksa centra funkcija ir analizēt un sintezēt informāciju, kā arī pārslēgt informāciju no aferenta uz eferento ceļu.
Refleksa loka funkcijas
Atkarībā no nervu sistēmas departamenta (somatiskā un veģetatīvā) refleksi, kuru centrs atrodas muguras smadzenēs, atšķiras ar starpkalāru neironu lokalizāciju. Tātad somatiskajai nervu sistēmai refleksu centrs atrodas starpzonā starp muguras smadzeņu priekšējiem un aizmugurējiem ragiem. Autonomās nervu sistēmas (starpkalāru neironu ķermeņa) refleksu centrs atrodas aizmugurējie ragi. Nervu sistēmas somatiskās un autonomās daļas atšķiras arī ar eferento neironu lokalizāciju. Somatiskās nervu sistēmas motoro neironu ķermeņi atrodas muguras smadzeņu priekšējos ragos, preganglionisko neironu ķermeņi autonoma sistēma- vidējo ragu līmenī.
Abu šūnu tipu aksoni veido refleksa loka eferento ceļu. Somatiskajā nervu sistēmā tas ir nepārtraukts, to veido A-α tipa šķiedras. Vienīgie izņēmumi ir A-γ šķiedras, kas vada ierosmi no muguras smadzeņu šūnām uz muskuļu vārpstu intrafuzālajām šķiedrām. Veģetatīvās nervu sistēmas eferentais ceļš tiek pārtraukts autonomais ganglijs atrodas vai intramurāls (pāris simpātiskā daļa), vai tuvu muguras smadzenēm (atsevišķi vai iekšā simpātisks stumbrs- simpātiskā daļa). Preganglioniskā šķiedra pieder B-šķiedrām, postganglioniskā šķiedra - C grupai.
Nervu sistēmas somatiskās daļas darba orgāns ir šķērssvītrotais skeleta muskulis, veģetatīvā lokā efektors ir dziedzeris vai muskulis (gluda vai svītraina sirds). Starp efektīvā veidā un darba orgāns ir ķīmiska mioneirāla vai neirosekrēcijas sinapse.
Refleksa loks noslēdzas gredzenā, pateicoties apgrieztajai aferentācijai - impulsu plūsmai no efektora receptoriem atpakaļ uz refleksu centru. Atgriezeniskās saites funkcija - signalizācija centrālajai nervu sistēmai par veikto darbību. Ja tas netiek veikts pietiekami, nervu centrs ir satraukts - reflekss turpinās. Arī apgrieztās aferentācijas dēļ tiek veikta centrālās nervu sistēmas perifērās aktivitātes kontrole.
Atšķiriet negatīvās un pozitīvās atsauksmes. Pirmais, veicot noteiktu funkciju, iedarbina mehānismu, kas kavē šo funkciju. Pozitīva atgriezeniskā saite sastāv no jau veiktas funkcijas turpmākas stimulēšanas vai jau nomāktas funkcijas kavēšanas. Pozitīva reversā aferentācija ir reti sastopama, jo tā noved bioloģisko sistēmu nestabilā stāvoklī.
Vienkāršie (monosinaptiskie) refleksu loki sastāv tikai no diviem neironiem (aferentiem un eferentiem) un atšķiras tikai ar proprioceptīviem refleksiem. Pārējie loki ietver visas iepriekš minētās sastāvdaļas.
Reflekss ir neironu ķēde no perifēra receptora caur centrālo nervu sistēmu līdz perifērajam efektoram. Refleksa loka elementi ir perifērais receptors, aferents ceļš, viens vai vairāki interneuroni, eferents ceļš un efektors.
Visi receptori ir iesaistīti noteiktos refleksos, tāpēc to aferentās šķiedras kalpo kā atbilstošā refleksa loka aferentais ceļš. Interneuronu skaits vienmēr ir lielāks par vienu, izņemot monosinaptisko stiepšanās refleksu. Eferento ceļu attēlo vai nu veģetatīvās nervu sistēmas motori aksoni vai postganglioniskās šķiedras, un efektori ir skeleta muskuļi un gludie muskuļi, sirds un dziedzeri.
Laiku no stimula sākuma līdz efektora reakcijai sauc par refleksa laiku. Vairumā gadījumu to galvenokārt nosaka vadīšanas laiks aferentā un eferentā ceļā un refleksa loka centrālajā daļā, kam jāpievieno stimula pārveidošanās laiks receptorā par izplatošo impulsu, pārraides laiks caur sinapsēm centrālajā nervu sistēmā (sinaptiskā aizkave), pārraides laiks no eferentā un aktivizēšanas ceļa vai efekta.
Refleksu loki ir sadalīti vairākos veidos
1. Monosinaptiskie refleksu loki - šādā lokā piedalās tikai viena sinapse, kas atrodas centrālajā nervu sistēmā. Šādi refleksi ir ļoti izplatīti visiem mugurkaulniekiem, tie ir iesaistīti regulācijā muskuļu tonuss un poza (piemēram, ceļgala raustīšanās). Šajos lokos neironi nesasniedz smadzenes, un refleksu darbības tiek veiktas bez tās līdzdalības, jo tās ir stereotipiskas un neprasa domu vai apzinātu lēmumu. Tie ir ekonomiski iesaistīto centrālo neironu skaita ziņā un izvairās no smadzeņu iejaukšanās.
2. Polisinaptiskie mugurkaula refleksu loki - tie ietver vismaz divas sinapses, kas atrodas centrālajā nervu sistēmā, jo lokā ir iekļauts trešais neirons - starpkalārs jeb starpneirons. Šeit ir sinapses starp sensoro neironu un interneuronu un starp starpkalariskajiem un motorajiem neironiem. Šādi refleksu loki ļauj ķermenim veikt automātiskas piespiedu reakcijas, kas nepieciešamas, lai pielāgotos ārējās vides izmaiņām (piemēram, skolēnu reflekss vai līdzsvara saglabāšana kustoties) un izmaiņas pašā organismā (elpošanas ātruma regulēšana, asinsspiediens utt.).
3. Polisinaptiskie refleksu loki, kuros iesaistītas gan muguras smadzenes, gan smadzenes – šāda veida refleksu lokos muguras smadzenēs notiek sinapse starp sensoro neironu un neironu, kas sūta impulsus smadzenēm.
Refleksus var klasificēt pēc dažādiem kritērijiem. Tātad, atkarībā no loka slēgšanas līmeņa, t.i. pēc refleksu centra atrašanās vietas refleksus iedala mugurkaula (reflekss aizveras muguras smadzenēs), bulbar (refleksa centrs ir iegarenās smadzenes), mezenfāliskajos (vidējos smadzenēs ir slēgts refleksu loks), diencefālajos un kortikālajos refleksu centros atrodas attiecīgi telencefalonā un smadzeņu garozā.
Pēc efektora pazīmes tie ir somatiski, kad refleksa eferentais ceļš nodrošina skeleta muskuļu motoro inervāciju, un veģetatīvi, kad efektori ir iekšējie orgāni.
Atkarībā no kairināto receptoru veida refleksus iedala eksteroceptīvajos (ja receptors uztver informāciju no ārējās vides), proprioceptīvajos (refleksa loks sākas no muskuļu un skeleta aparāta receptoriem) un interoceptīvajos (no iekšējo orgānu receptoriem).
Interoceptīvie refleksi savukārt tiek iedalīti viscero-viscerālajos (refleksa loks savieno divus iekšējos orgānus), viscero-muskuļajos (receptori atrodas uz muskuļu-cīpslu aparāta, efektors ir iekšējais orgāns) un viscero-kutānos (receptori lokalizēti ādā, darba orgāni ir iekšpuses).
Pēc Pavlova domām, refleksus iedala kondicionētajos (dzīves laikā izstrādātos, katram indivīdam raksturīgos) un beznosacījumu (iedzimtos, sugai raksturīgos: pārtikas, seksuālos, aizsardzības-motoros, homeostatiskie utt.).
Neatkarīgi no refleksa veida tā refleksa loks satur receptoru, aferento ceļu, nervu centru, eferento ceļu, darba orgānu un atgriezenisko saiti. Izņēmums ir aksona refleksi, kuru refleksu loks atrodas viena neirona ietvaros: maņu procesi rada centripetālus impulsus, kas, ejot cauri neirona ķermenim, pa aksonu izplatās uz centrālo nervu sistēmu, un pa aksona atzaru impulsi sasniedz efektoru. Šādi refleksi tiek attiecināti uz metasimpatiskās nervu sistēmas darbību, caur tiem, piemēram, tiek veikti mehānismi asinsvadu tonusa un ādas dziedzeru darbības regulēšanai.
Kairinājuma uztveršanas un pārvēršanas ierosmes enerģijā funkciju veic refleksu loku receptori. Uzbudinājuma receptoru enerģijai ir lokālas reakcijas raksturs, kas ir svarīgs ierosmes gradācijā pēc spēka.
Pamatojoties uz receptoru uzbūvi un izcelsmi, tos var iedalīt primārajos sensorajos, sekundārajos sensorajos un brīvajos nervu galos. Pirmajā neirons pats darbojas kā receptors (tas attīstās no neiroepitēlija); starp stimulu un pirmo aferento neironu nav starpstruktūru. Primāro sensoro receptoru lokālā reakcija – receptoru potenciāls – arī ir ģeneratora potenciāls, t.i. izraisot darbības potenciālu pāri aferentās šķiedras membrānai. Primārie sensorie receptori ir sirds un asinsvadu sistēmas vizuālie, ožas, ķīmij- un baroreceptori.
Sekundārās sensorās šūnas ir īpašas nenervu izcelsmes struktūras, kas mijiedarbojas ar pseidounipolāru sensoro šūnu dendritiem ar sinaptisko neiroreceptoru kontaktu palīdzību. Receptoru potenciāls, kas rodas stimula iedarbībā sekundārās sensorās šūnās, nav ģenerators un neizraisa darbības potenciāla parādīšanos aferentās šķiedras membrānā. Uzbudinošais postsinaptiskais potenciāls rodas tikai caur receptoru šūnu mediatora atbrīvošanas mehānismu. Stimulēja stipruma gradācija tiek veikta, izvadot dažādus mediatora daudzumus (jo vairāk tiek atbrīvots mediators, jo spēcīgāks ir stimuls).
Sekundārās sensorās šūnas ietver dzirdes, vestibulārās, miega, taustes un citus receptorus. Dažkārt funkcionēšanas īpatnību dēļ šajā grupā ietilpst fotoreceptori, kas no anatomiskā viedokļa un to izcelsmes dēļ no neiroepitēlija ir sekundāri sensējoši.
Brīvie nervu gali ir pseido-unipolāru sensoro šūnu zarojoši dendriti un ir lokalizēti gandrīz visos cilvēka ķermeņa audos.
Atkarībā no stimula enerģētiskā rakstura, uz kuru receptors reaģē, tos iedala mehānoreceptoros (taustes, baroreceptoros, tilpuma receptoros, dzirdes, vestibulārajos; tie parasti uztver mehānisku kairinājumu ar šūnu izaugumu palīdzību), ķīmijreceptoros (ožas), asinsvadu uztverošajos (centrālās nervu sistēmas, foto-receptoros, fotoreceptoros). šūnas izaugumi), termoreceptori (reaģē uz izmaiņām "siltums-aukstums" - Rufini ķermeņi un Krauzes kolbas no gļotādas) un nociceptori (nekapsulēti sāpju gali).
Refleksu loku pēcreceptoru veidošanās ir aferents ceļš, ko veido pseido-unipolārs sensorais neirons, kura ķermenis atrodas mugurkaula ganglijā, un aksoni veido muguras smadzeņu aizmugurējās saknes. Aferentā ceļa funkcija ir novadīt informāciju uz centrālo saiti, turklāt šajā posmā informācija tiek kodēta. Šiem nolūkiem mugurkaulnieku ķermenī tiek izmantots binārs kods, ko veido impulsu pārrāvumi (zalves) un spraugas starp tiem. Ir divi galvenie kodēšanas veidi: frekvence un telpiskā.
Pirmais ir dažāda impulsu skaita veidošanās sprādzienā, atšķirīgs uzliesmojumu skaits, to ilgums un pārtraukumu ilgums starp tiem atkarībā no receptoram pielietotās stimulācijas stipruma. Telpiskā kodēšana veic stimula stipruma gradāciju, iesaistot atšķirīgu nervu šķiedru skaitu, pa kurām vienlaikus tiek veikta ierosme.
Aferentā ceļa sastāvs galvenokārt ietver A-b, A-c un A-e šķiedras.
Izejot cauri šķiedrām, nervu impulss nonāk refleksu centrā, kas anatomiskā nozīmē ir neironu kopums, kas atrodas noteiktā centrālās nervu sistēmas līmenī un piedalās šī refleksa veidošanā. Refleksa centra funkcija ir analizēt un sintezēt informāciju, kā arī pārslēgt informāciju no aferenta uz eferento ceļu.
Atkarībā no nervu sistēmas departamenta (somatiskā un veģetatīvā) refleksi, kuru centrs atrodas muguras smadzenēs, atšķiras ar starpkalāru neironu lokalizāciju. Tātad somatiskajai nervu sistēmai refleksu centrs atrodas starpzonā starp muguras smadzeņu priekšējiem un aizmugurējiem ragiem. Autonomās nervu sistēmas refleksu centrs (starpkalāru neironu ķermeņi) atrodas aizmugurējos ragos. Nervu sistēmas somatiskās un autonomās daļas atšķiras arī ar eferento neironu lokalizāciju. Somatiskās nervu sistēmas motoro neironu ķermeņi atrodas muguras smadzeņu priekšējos ragos, autonomās sistēmas preganglionisko neironu ķermeņi atrodas vidējo ragu līmenī.
Abu šūnu tipu aksoni veido refleksa loka eferento ceļu. Somatiskajā nervu sistēmā tā ir nepārtraukta, sastāv no šķiedrām tips A-b. Vienīgie izņēmumi ir A-g šķiedras, kas vada ierosmi no muguras smadzeņu šūnām uz muskuļu vārpstu intrafuzālajām šķiedrām. Veģetatīvās nervu sistēmas eferents ceļš tiek pārtraukts veģetatīvā ganglijā, kas atrodas vai nu intramurāli (parasimpatiskā daļa), vai muguras smadzeņu tuvumā (atsevišķi vai simpātiskajā stumbrā - simpātiskā daļa). Preganglioniskā šķiedra pieder B-šķiedrām, postganglioniskā šķiedra pieder C grupai.
Nervu sistēmas somatiskās daļas darba orgāns ir šķērssvītrots skeleta muskulis, veģetatīvā lokā efektors ir dziedzeris vai muskulis (gluda vai svītraina sirds). Starp eferento ceļu un darba orgānu atrodas ķīmiska mioneirāla vai neirosekrēcijas sinapse.
Refleksa loks noslēdzas gredzenā, pateicoties reversajai aferentācijai - impulsu plūsmai no efektorreceptoriem atpakaļ uz refleksu centru. Atgriezeniskās saites funkcija - signalizācija centrālajai nervu sistēmai par veikto darbību. Ja tas netiek veikts pietiekami, nervu centrs ir satraukts - reflekss turpinās. Arī apgrieztās aferentācijas dēļ tiek veikta centrālās nervu sistēmas perifērās aktivitātes kontrole.
Atšķiriet negatīvās un pozitīvās atsauksmes. Pirmais, veicot noteiktu funkciju, iedarbina mehānismu, kas kavē šo funkciju. Pozitīva atgriezeniskā saite sastāv no jau veiktas funkcijas turpmākas stimulēšanas vai jau nomāktas funkcijas kavēšanas. Pozitīva reversā aferentācija ir reti sastopama, jo tā noved bioloģisko sistēmu nestabilā stāvoklī.
Vienkāršie (monosinaptiskie) refleksu loki sastāv tikai no diviem neironiem (aferentiem un eferentiem) un atšķiras tikai ar proprioceptīviem refleksiem. Pārējie loki ietver visas iepriekš minētās sastāvdaļas.
Nervu šķiedru fizioloģiskās īpašības un funkcionālā nozīme
Nervu šķiedrām ir visaugstākā uzbudināmība, visaugstākais ierosmes vadīšanas ātrums, īsākais ugunsizturības periods un augsta labilitāte. Tas tiek nodrošināts augsts līmenis vielmaiņas procesi un zems membrānas potenciāls.
Funkcija: nervu impulsu vadīšana no receptoriem uz centrālo nervu sistēmu un otrādi.
Nervu šķiedru struktūras īpatnības un veidi
Nervu šķiedra - aksons - ir pārklāta ar šūnu membrānu.
Ir divu veidu nervu šķiedras:
Nemielinizētas nervu šķiedras – viens Švāna šūnu slānis, starp tām – spraugām līdzīgas atstarpes. šūnu membrānu visā saskarsmē ar vidi. Kad tiek pielietots kairinājums, stimula darbības vietā rodas ierosme. Nemielinizētām nervu šķiedrām ir elektrogēnas īpašības (spēja radīt nervu impulsus).
Mielinētas nervu šķiedras - pārklātas ar Švāna šūnu slāņiem, kas vietām veido Ranvier mezglus (zonas bez mielīna) ik pēc 1 mm. Ranvier pārtveršanas ilgums ir 1 µm. Mielīna apvalks veic trofiskās un izolācijas funkcijas (augsta pretestība). Ar mielīnu pārklātajām vietām nav elektrogēnu īpašību. Viņiem ir Ranvier pārtvertās bumbas. Uzbudinājums notiek Ranvjē pārtveršanā, kas atrodas vistuvāk stimula darbības vietai. Ranvjē pārtvērumos liels blīvums Na-kanāli, tāpēc katrā Ranvier pārtveršanā palielinās nervu impulsi.
Ranvier pārtveršana darbojas kā atkārtotāji (ģenerē un pastiprina nervu impulsus).
Uzbudinājuma vadīšanas mehānisms gar nervu šķiedru
1885. gads - L. Vācu valoda - starp nervu šķiedras ierosinātajām un nesatrauktajām sekcijām rodas apļveida strāvas.
Kairinātāja iedarbībā pastāv potenciāla atšķirība starp audu ārējo un iekšējo virsmu (zonām, kurās ir dažādi lādiņi). Starp šīm zonām rodas elektriskā strāva (Na + jonu kustība). Nervu šķiedras iekšpusē strāva rodas no pozitīvā pola uz negatīvo polu, t.i., strāva tiek virzīta no ierosinātās zonas uz neierosināto. Šī strāva iziet caur neuzbudināto reģionu un izraisa tā uzlādi. Uz nervu šķiedras ārējās virsmas strāva plūst no neuzbudinātās zonas uz ierosināto zonu. Šī strāva nemaina ierosinātās zonas stāvokli, jo tā ir ugunsizturīgā stāvoklī.
Pierādījumi par apļveida strāvu klātbūtni: nervu šķiedra ir ievietota NaCl šķīdums un reģistrē ierosmes ātrumu. Tad nervu šķiedra tiek ievietota eļļā (pretestība palielinās) - vadīšanas ātrums samazinās par 30%. Pēc tam nervu šķiedra tiek atstāta gaisā - ierosmes ātrums tiek samazināts par 50%.
Uzbudinājuma vadīšanas pazīmes pa mielinizētām un nemielinizētām nervu šķiedrām:
mielīna šķiedras - ir apvalks ar augstu pretestību, elektrogēnas īpašības tikai Ranvier mezglos. Stimulēšanas ietekmē uzbudinājums notiek Ranvier tuvākajā krustpunktā. Kaimiņa pārtveršana polarizācijas stāvoklī. Iegūtā strāva izraisa blakus esošā krustpunkta depolarizāciju. Ranvier mezglos ir liels Na kanālu blīvums, tāpēc katrā nākamajā mezglā rodas nedaudz lielāks (amplitūdā) darbības potenciāls, līdz ar to ierosme izplatās bez samazinājuma un var pārlēkt pāri vairākiem mezgliem. Šī ir Tasaki sāļā teorija. Teorijas pierādījums ir tāds, ka nervu šķiedrā tika ievadītas zāles, kas bloķē vairākus pārtveršanas punktus, bet ierosmes vadīšana tika reģistrēta pēc tam. Šī ir ļoti uzticama un izdevīga metode, jo tiek novērsti nelieli bojājumi, tiek palielināts ierosmes ātrums un samazinātas enerģijas izmaksas;
nemielinizētas šķiedras – virsmai visā garumā piemīt elektrogēnas īpašības. Tāpēc nelielas apļveida strāvas rodas dažu mikrometru attālumā. Uzbudinājumam ir pastāvīgi ceļojoša viļņa forma.
Šī metode ir mazāk rentabla: augstas enerģijas izmaksas (Na-K sūkņa darbībai), zemāks ierosmes ātrums.
Nervu šķiedru klasifikācija
Nervu šķiedras klasificē pēc:
darbības potenciāla ilgums;
šķiedras struktūra (diametrs);
ierosmes ātrums.
Izšķir šādas nervu šķiedru grupas:
A grupa (alfa, beta, gamma, delta) - īsākais darbības potenciāls, biezākais mielīna apvalks, visaugstākais ierosmes ātrums;
B grupa - mielīna apvalks ir mazāk izteikts;
C grupa - bez mielīna apvalka.
Morfoloģiskās atšķirības starp dendritiem un aksoniem
1. Atsevišķam neironam ir vairāki dendrīti, aksons vienmēr ir viens.
2. Dendrīti vienmēr ir īsāki par aksonu. Ja dendrītu izmērs nepārsniedz 1,5-2 mm, tad aksoni var sasniegt 1 m vai vairāk.
3. Dendriti vienmērīgi attālinās no šūnas ķermeņa un pakāpeniski iegūst nemainīgu diametru ievērojamā attālumā.
4. Dendriti parasti zarojas zem akūts leņķis, un zari ir vērsti prom no šūnas. Aksoni nodrošinājumus izdala visbiežāk taisnā leņķī, nodrošinājumu orientācija nav tieši saistīta ar šūnas ķermeņa stāvokli.
5. Dendrīta sazarojuma modelis tāda paša tipa šūnās ir konstantāks nekā šo šūnu aksona sazarojums.
6. Nobriedušu neironu dendrīti ir pārklāti ar dendrītiskajiem muguriņiem, kuru nav uz somas un dendrītu stumbru sākuma daļas. Aksoniem nav muguriņu.
7. Dendritiem nekad nav mīkstuma apvalka. Aksonus bieži ieskauj mielīns.
8. Dendritiem ir regulārāka mikrotubulu telpiskā organizācija, aksonos dominē neirofilamenti un mikrotubulās ir mazāk sakārtotas.
9. Dendritos, īpaši to proksimālajās daļās, atrodas endoplazmatiskais tīklojums un ribosomas, kuras nav aksonos.
10. Dendrītu virsma vairumā gadījumu ir saskarē ar sinoptiskām plāksnēm un tajā ir aktīvās zonas ar postsinaptisku specializāciju.
Dendrītu struktūra
Ja ir salīdzinoši liela literatūra par dendrītu ģeometriju, to zaru garumu, orientāciju, tad apm. iekšējā struktūra, par to citoplazmas atsevišķo komponentu struktūru, ir tikai atsevišķa izkaisīta informācija. Šī informācija kļuva iespējama tikai ar elektronu mikroskopisko pētījumu ieviešanu neirohistoloģijā.
Galvenās dendrīta raksturīgās pazīmes, kas to atšķir elektronu mikroskopiskos griezumos:
1) mielīna apvalka trūkums,
pareizas mikrotubulu sistēmas klātbūtne,
3) uz tām ir aktīvas sinapses zonas ar skaidri izteiktu dendrīta citoplazmas elektronu blīvumu,
4) atkāpšanās no mugurkaula dendrīta kopējā stumbra,
5) speciāli organizētas zaru mezglu zonas,
6) ribosomu iekļaušana,
7) granulēta un negranulāra endoplazmatiskā tīkla klātbūtne proksimālajās zonās.
Visievērojamākā dendrītiskās citoplazmas iezīme ir daudzu mikrotubulu klātbūtne. Tie ir labi identificēti gan šķērsgriezumos, gan garengriezumos. Sākot no dendrīta proksimālās daļas, mikrocaurules iet paralēli dendrīta garajai asij līdz tā distālajiem zariem. Mikrocaurules seko dendrītā paralēli viena otrai, savstarpēji nesavienojoties un nekrustojas. Šķērsgriezumos var redzēt, ka attālumi starp atsevišķiem kanāliņiem ir nemainīgi. Atsevišķi dendrītu kanāliņi stiepjas diezgan lielos attālumos, bieži sekojot līknēm, kas var būt gar dendrītu gaitu. Cauruļu skaits ir relatīvi nemainīgs uz dendrīta šķērsgriezuma laukuma vienību un ir aptuveni 100 uz 1 µm. Šis skaitlis ir raksturīgs visiem dendritiem, kas ņemti no dažādas nodaļas centrālā un perifērā nervu sistēma, dažādi veidi dzīvnieki.
Mikrotubulu funkcija ir vielu transportēšana pa nervu šūnu procesiem.
Iznīcinot mikrotubulas, var tikt traucēta vielu transportēšana dendrītā, līdz ar to procesu gala posmos tiek liegta barības vielu pieplūde un enerģētiskās vielas no šūnas ķermeņa. Dendrīti, lai ekstremālos apstākļos saglabātu sinaptisko kontaktu struktūru un tādējādi nodrošinātu starpneironu mijiedarbības funkciju, kompensē deficītu barības vielas tām blakus esošo struktūru dēļ (sinaptiskās plāksnes, mīksto šķiedru daudzslāņu mielīna apvalks, kā arī glia šūnu fragmenti).
Ja patogēnā faktora darbība tiek novērsta savlaicīgi, dendriti atjauno mikrotubulu struktūru un pareizu telpisko organizāciju, tādējādi atjaunojot normālām smadzenēm raksturīgo vielu transportēšanas sistēmu. Ja patogēnā faktora stiprums un ilgums ir nozīmīgs, tad endocitozes parādības, nevis adaptīvā funkcija, var kļūt nāvējošas dendritiem, jo fagocitētie fragmenti nevar tikt izmantoti un, uzkrājoties dendrītu citoplazmā, novedīs pie tā neatgriezeniskiem bojājumiem.
Mikrotubulu organizācijas pārkāpums izraisa asas izmaiņas dzīvnieku uzvedībā. Dzīvniekiem, kuriem eksperimentā tika iznīcināti mikrotubuli dendritos, tika novērota sarežģītu uzvedības formu dezorganizācija, saglabājot vienkāršus kondicionētus refleksus. Cilvēkiem tas var izraisīt nopietnus augstākas nervu darbības traucējumus.
Fakts, ka dendrīti ir visjutīgākais lokuss pret patoloģiskā aģenta darbību garīga slimība, liecina daži amerikāņu zinātnieku darbi. Izrādījās, ka senils demences (cianotiskās demences) un Alcheimera slimības gadījumā smadzeņu preparāti, kas apstrādāti ar Golgi metodi, neatklāj nervu šūnu procesus. Šķiet, ka dendrītu stumbri ir apdeguši un pārogļojušies. Šo procesu nenoteikšana uz smadzeņu histoloģiskajiem preparātiem, iespējams, ir saistīta arī ar mikrotubulu un neirofilamentu sistēmas pārkāpumu šajos procesos.
Atrodas dendritos. Tie seko paralēli dendrīta garajai asij, tie var atrasties atsevišķi vai savākti saišķos, bet tie nav stingri izvietoti citoplazmā. Iespējams, kopā ar mikrotubulām tās var būt līdzvērtīgas neirofibrilām.
Visiem CNS dendritiem ir raksturīgs virsmas pieaugums vairāku dihotomu dalījumu dēļ. Šajā gadījumā sadalīšanas zonās tiek veidotas īpašas izplešanās vietas vai zaru mezgli.
Parastā analīze parāda, ka atzarojuma mezglā, kuram tuvojas divi dendritiskie zari, katrs nesot savu signālu, var veikt šādas darbības. Caur filiāles mezglu kopīgs stumbrs un tālāk uz neirona pārejas ķermeni:
vai signāls no vienas filiāles,
vai vienkārši no cita
vai divu signālu mijiedarbības rezultāts,
vai signāli izslēdz viens otru.
Atzarojuma mezgla citoplazmā ir gandrīz visi organismam raksturīgie komponenti nervu šūna, un vietas pēc savas struktūras krasi atšķiras no kopējā dendrītiskā stumbra citoplazmas un zariem, kas iegūti dalīšanas ceļā. Atzaru mezglos ir palielināts mitohondriju skaits, granulēts un gluds tīklojums, redzamas atsevišķu ribosomu kopas un rozetēs saliktas ribosomas. Šīs sastāvdaļas (granulārais un gludais tīkls, ribosomas) ir tieši iesaistītas olbaltumvielu sintēzē. Mitohondriju uzkrāšanās šajās vietās liecina par oksidatīvo procesu intensitāti.
Dendrītu funkcijas
Vēlos atzīmēt, ka galvenās grūtības, ar kurām pētnieks sastopas, pētot dendrītu funkciju, ir informācijas trūkums par dendrīta membrānas īpašībām (pretstatā neirona ķermeņa membrānai), jo nav iespējams dendrītā ievietot mikroelektrodu.
Vērtējot dendrītu kopējo ģeometriju, sinapšu izplatību un citoplazmas īpašo struktūru dendrītu atzarojuma vietās, var runāt par īpašiem neironu lokusiem ar savu funkciju. Vienkāršākā lieta, ko varētu attiecināt uz dendrītu vietām sazarošanās vietās, ir trofiskā funkcija.
No iepriekš minētā izriet, ka dendrītu citoplazmā ir daudz ultra strukturālās sastāvdaļas spēj nodrošināt savas svarīgās funkcijas. Dendrītā ir noteikti loki, kur tā darbībai ir savas īpatnības.
Nervu šūnas daudzo dendritisko zaru galvenais mērķis ir nodrošināt savstarpēju savienojumu ar citiem neironiem. zīdītāju smadzeņu garozā liela daļa aksodendri savienojumi krīt uz kontaktiem ar īpašiem specializētiem dendrītu izaugumiem - dendrītiem. Dendrītiskie muguriņi ir filoģenētiski jaunākie nervu sistēmas veidojumi. Ontoģenēzē tie nobriest daudz vēlāk nekā citas nervu struktūras un pārstāv nervu šūnas plastiskāko aparātu.
Parasti dendritiskajam mugurkaulam ir raksturīga forma zīdītāju smadzeņu garozā. (2. att.). No galvenā dendrīta stumbra atkāpjas salīdzinoši šaurs kātiņš, kas beidzas ar pagarinājumu – galvu. Visticamāk, ka šī dendrīta piedēkļa forma (galvas klātbūtne), no vienas puses, ir saistīta ar sinaptiskā kontakta laukuma palielināšanos ar aksona galu, un, no otras puses, tā kalpo specializētu organellu ievietošanai mugurkaula iekšpusē, jo īpaši mugurkaula aparātā, kas atrodas tikai zīdītāju smadzeņu smadzeņu mugurkaulā. Šajā sakarā šķiet piemērota līdzība ar sinaptiskā aksona gala formu, kad plāna priekšgala šķiedra veido pagarinājumu. Šī izplešanās (sinaptiskā plāksne) veido plašu kontaktu ar inervēto substrātu un satur lielu ultrastrukturālo komponentu komplektu (sinaptiskās pūslīši, mitohondriji, neirofilamenti, glikogēna granulas).
Pastāv hipotēze (kuru it īpaši dala un izstrādā Nobela prēmijas laureāts F. Kriks), ka mugurkaula ģeometrija var mainīties atkarībā no smadzeņu funkcionālā stāvokļa. Šajā gadījumā šaurais mugurkaula kakls var paplašināties, un pats mugurkauls saplacinās, kā rezultātā palielinās akso-mugurkaula kontakta efektivitāte.
Ja dendritisko muguriņu forma un izmērs zīdītāju smadzeņu garozā var nedaudz atšķirties, tad viskonstantākā tajos ir noteikta mugurkaula aparāta klātbūtne. Tas ir savstarpēji savienotu kanāliņu (cisternu) komplekss, kas, kā likums, atrodas mugurkaula galvā. Iespējams, šī organelle ir saistīta ar ļoti svarīgām funkcijām, kas raksturīgas filoģenētiski jaunākajiem smadzeņu veidojumiem, jo smailais aparāts ir atrodams galvenokārt smadzeņu garozā un tikai augstākiem dzīvniekiem.
Neskatoties uz visu, mugurkauls ir dendrīta atvasinājums, tam trūkst neirofilamentu un dendrītu kanāliņu, tā citoplazmā ir rupji vai smalki granulēta matrica. Vēl vienu funkciju mugurkauls smadzeņu garozā ir obligāta sinaptisko kontaktu klātbūtne ar aksonu galiem uz tiem. Mugurkaula citoplazmā ir īpaši komponenti, kas to atšķir no dendrītiskajiem kātiem. Mugurkaula citoplazmā var atzīmēt savdabīgu triādi: aktīvo zonu subsinaptiskā specializācija - smailais aparāts - mitohondriji. Ņemot vērā mitohondriju veikto sarežģīto un svarīgo funkciju daudzveidību, sinaptiskās transmisijas laikā var sagaidīt arī sarežģītas funkcionālas izpausmes "triādēs". Var teikt, ka dendrītiskā mugurkaula citoplazma un mugurkaula aparāts var būt tieši saistīti ar sinaptisko funkciju.
Dendrītu muguriņas un dendrītu gali ir arī ļoti jutīgi pret ekstremāliem faktoriem. Ar jebkāda veida saindēšanos (piemēram, alkoholisks, hipoksisks, smagie metāli - svins, dzīvsudrabs utt.) mainās uz smadzeņu garozas šūnu dendritiem konstatēto muguriņu skaits. Visticamāk, muguriņas nepazūd, bet to citoplazmas komponenti tiek traucēti, un tie ir sliktāk piesūcināti ar sāļiem. smagie metāli. Tā kā muguriņas ir viena no starpneironu kontaktu strukturālajām sastāvdaļām, to darbības traucējumi izraisa nopietnus smadzeņu darbības traucējumus.
Dažos gadījumos ar īslaicīgu ekstrēma faktora darbību no pirmā acu uzmetiena var rasties paradorsāla situācija, kad smadzeņu šūnu dendritos konstatēto muguriņu skaits nevis samazinās, bet palielinās. Tātad tas tiek novērots eksperimentālās cerebrālās išēmijas laikā tās sākotnējā periodā. Paralēli identificēto muguriņu skaita pieaugumam, funkcionālais stāvoklis smadzenes. Šajā gadījumā hipoksija ir faktors, kas veicina vielmaiņas palielināšanos nervu audos, labāku to rezervju ieviešanu, kuras netiek izmantotas. normālu vidi, un organismā uzkrāto toksīnu ātra sadegšana. Ultrastrukturāli tas izpaužas intensīvākā mugurkaula citoplazmas izpētē, mugurkaula aparāta cisternu augšanā un paplašināšanā. Iespējams, šī hipoksijas pozitīvās ietekmes parādība tiek novērota, ja cilvēks piedzīvo lielu fiziski vingrinājumi hipoksijas apstākļos iekaro kalnu virsotnes. Šīs grūtības pēc tam tiek kompensētas ar intensīvāku produktīvu darbu gan smadzenēs, gan citos orgānos.
Dendrītu veidošanās
Dendrīti un to starpneironālie savienojumi veidojas smadzeņu ontoģenētiskās attīstības laikā. Turklāt dendriti, jo īpaši apikālie, jauniem indivīdiem kādu laiku paliek brīvi, lai izveidotu jaunus kontaktus. Dendrīta daļas, kas atrodas tuvāk šūnas ķermenim, iespējams, ir saistītas ar spēcīgākiem un vienkāršākiem dabiskiem nosacītiem refleksiem, un galus atstāj jaunu savienojumu un asociāciju veidošanai.
Pieaugušā vecumā uz dendritiem vairs nav no starpneironu kontaktiem brīvas vietas, bet novecojot, vispirms cieš dendrītu gali un piesātinājums ar kontaktiem.
veciem indivīdiem tie atgādina dendritus bērnība. Tas notiek gan sakarā ar transporta proteīnu sintēzes procesu pavājināšanos šūnā, gan sakarā ar traucētu asins piegādi smadzenēm. Iespējams, tieši šeit slēpjas morfoloģiskais pamats tik labi zināmam neiroloģijā un citā ikdiena to, ka, kad veciem cilvēkiem ir grūti apgūt ko jaunu, viņi bieži aizmirst aktuālos notikumus un ļoti labi atceras pagātni. Tas pats tiek novērots saindēšanās gadījumā.
Kā jau minēts, dendrītiskā koka palielināšanās un komplikācija filoģenēzē ir nepieciešama ne tikai uztverei liels skaits ienākošajiem impulsiem, bet arī pirmapstrādei.
Centrālās nervu sistēmas neironu dendritiem ir sinaptiska funkcija, un gala sekcijas nekādā ziņā nav zemākas par vidējām. Ja mēs runājam par apikālo dendrītu distālajām (terminālajām) sekcijām piramīdveida neironi smadzeņu garozā, tad to īpatsvars starpneironu mijiedarbības īstenošanā ir vēl nozīmīgāks par proksimālajām. Tur, lai vairāk terminālas sinaptiskās plāksnes uz paša stumbra un apikālā dendrīta zariem tiek savienotas ar vairāk kontaktu uz dendrītiskajiem muguriņiem.
Pētot šo problēmu, izmantojot elektronu mikroskopiju, pētnieki arī pārliecinājās, ka dendrītu gala sekcijas ir blīvi pārklātas ar sinaptiskām plāksnēm un tādējādi ir tieši iesaistītas starpneironu mijiedarbībā. elektronu mikroskopija arī parādīja, ka dendriti var veidot kontaktus viens ar otru. Šie kontakti var būt vai nu paralēli, kam vairums autoru piedēvē elektrotoniskas īpašības, vai tipiskas asimetriskas sinapses ar labi definētām organellām, kas nodrošina ķīmisko transmisiju. Šādi dendrodendrītiskie kontakti tikai sāk piesaistīt pētnieku uzmanību. Tātad dendrīts visā tā garumā veic sinaptisku funkciju. Kā dendrīta virsma ir pielāgota kontaktu nodrošināšanai ar aksonu galiem?
Dendrīta virsmas membrāna ir veidota tā, lai to maksimāli izmantotu starpneironu kontaktiem. Viss dendrīts ir bedrīts ar iedobumiem, krokām, kabatām, ar dažādiem mikroizauguma veida nelīdzenumiem, tapas, sēņveidīgiem piedēkļiem uc Visi šie dendrītu stumbru reljefi atbilst ienākošo sinaptisko galu formai un izmēram. Un iekšā dažādas nodaļas nervu sistēma un dažādiem dzīvniekiem dendrīta virsmas reljefam ir specifiskas iezīmes. Protams, visievērojamākais dendrītiskās membrānas izaugums ir dendrīta mugurkauls.
Dendrīti ir ļoti jutīgi pret dažādu ekstrēmu faktoru iedarbību. Pārkāpumi tajos noved pie daudzām slimībām, piemēram, garīgiem traucējumiem.
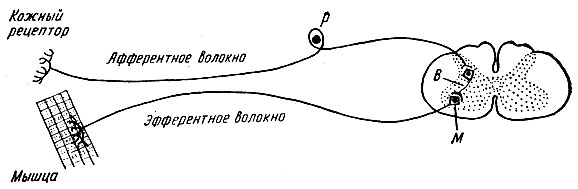
Refleksa loks sastāv no:
- receptori - uztver kairinājumu.
- jutīga (centripetāla, aferenta) nervu šķiedra, kas pārraida ierosmi uz centru
- nervu centrs, kurā notiek ierosmes pārslēgšana no sensorajiem neironiem uz motoriem neironiem
- motora (centrbēdzes, eferenta) nervu šķiedra, kas veic ierosmi no centrālās nervu sistēmas uz darba orgānu
- efektors - darba orgāns, kas veic efektu, reakciju, reaģējot uz receptoru kairinājumu.
Receptori un uztveres lauki
Receptors- šūna, kas uztver kairinājumu.
uztverošs lauks- tas ir anatomiskais apgabals, kairinājuma gadījumā rodas šis reflekss.
Primāro sensoro receptoru uztveres lauki ir sakārtoti visvienkāršākajā veidā. Piemēram, ādas virsmas taustes jeb nociceptīvais uztverošais lauks ir vienas maņu šķiedras atzarojums.
Receptoriem, kas atrodas dažādās uztverošā lauka daļās, ir atšķirīga jutība pret adekvātu stimulāciju. Ļoti jutīga zona parasti atrodas uztverošā lauka centrā, un jutība samazinās tuvāk uztverošā lauka perifērijai.
Sekundāro sensoro receptoru uztveres lauki tiek organizēti līdzīgi. Atšķirība ir tāda, ka aferentās šķiedras zari nebeidzas brīvi, bet tiem ir sinaptiski kontakti ar jutīgām receptoršūnām. Tādā veidā tiek organizēti garšas, vestibulārās, akustiskās uztveres lauki.
pārklājas uztverošie lauki. Vienu un to pašu jutīgās virsmas laukumu (piemēram, ādu vai tīkleni) inervē vairāki jutīgi nervu šķiedras, kas ar saviem atzarojumiem pārklāj atsevišķu aferento nervu uztverošos laukus.
Receptīvo lauku pārklāšanās dēļ palielinās kopējā ķermeņa sensorā virsma.
Refleksu klasifikācija.
Pēc izglītības veida:
Nosacīts (iegūts) - reaģē uz vārdu, siekalas no suņa nonāk gaismā.
Beznosacījuma (iedzimta) - mirgojoša rīšana, ceļgals.
Pēc atrašanās vietas receptori:
Eksteroceptīvs (ādas, redzes, dzirdes, ožas)
Interoceptīvs (no iekšējo orgānu receptoriem)
Proprioceptīvs (no receptoriem muskuļos, cīpslās, locītavās)
Efektoriem:
Somatiski vai motoriski (refleksi skeleta muskulis);
Veģetatīvie iekšējie orgāni – gremošanas, sirds un asinsvadu, izdales, sekrēcijas u.c.
Pēc bioloģiskās izcelsmes:
Aizsardzības vai aizsargājoša (reakcija uz taustes sāpju sadalījumu)
Gremošana (kairinoši receptori mutes dobumā.)
Seksuāls (hormoni asinīs)
Aptuvens (galvas pagrieziens, ķermenis)
Motors
Posotonisks (atbalsta ķermeņa pozas)
Pēc sinapšu skaita:
Monosinaptisks, kura loki sastāv no aferentiem un eferentiem neironiem (piemēram, ceļgala).
Polisinaptisks, kura lokos ir arī 1 vai vairāki starpposma neironi un ir 2 vai vairāki sinaptiskie slēdži. (somatiskās un veģetatīvās atsauces).
Disinaptisks (2 sinapses, 3 neironi).
Pēc atbildes rakstura:
Motors \ motors (muskuļu kontrakcijas)
Sekretārs (sekrēcijas dziedzeru sekrēcija)
Vasomotors (asinsvadu paplašināšanās un sašaurināšanās)
Sirds (mainīt. Sirds muskuļa darbs.)
Pēc ilguma:
fāziska (ātra) rokas atvilkšana
tonizējoša (lēna) stājas uzturēšana
Atkarībā no nervu centra atrašanās vietas:
Mugurkauls (iekļauti SM neironi) - Rokas atvilkšana no karstajiem segmentiem 2-4, ceļgala raustīšanās.
refleksi smadzenēs
Bulbar (medulla oblongata) - plakstiņu aizvēršana pieskaroties. uz radzeni.
Mesencephalic (vidējais m) - redzes orientieris.
Diencephalic (vidussmadzenes) - ožas sajūta
Kortikāls (miza BP GM) - nosacīts. ref.
Īpašības nervu centri.
1. Vienpusēja ierosmes izplatīšanās.
Uzbudinājums tiek pārraidīts no aferenta uz eferento neironu (iemesls: sinapses struktūra).
Uzbudinājuma pārnešanas palēnināšanās.
Kondicionieris Daudzu sinapšu klātbūtne ir atkarīga arī no kairinātāja stipruma (summēšanas) un no fiziskā stāvokļa. CNS (nogurums).
3.Summēšana efektu summēšana, stimuli zem sliekšņa.
Pagaidu: ref. No iepriekšējā Imp-sa vēl nav pagājis, bet pēdas. Jau ieradās.
Telpiskais: sajaucot vairākus. Aizūdens Tās ir pūces kondicionētas. Attēli. Atsauce
Fasilitācijas un oklūzijas centrs.
Parādījās atvieglojuma centrs - rodas optimālā stimula (maksimālā reakcija) ietekmē. Palīdzības centrs.
Zem darbības min irr. (samazināts otv. Rektsiya) bija oklūzija.
Uzbudinājuma ritma asimilācija un transformācija.
Transformācija - nervu impulsa frekvences maiņa, ejot caur nervu centru. Biežumu var palielināt vai samazināt.
Asimilācija (deja, ikdienas rutīna)
Sekas
Reakcijas beigu aizkavēšanās pēc stimula pārtraukšanas. Saistīts ar asinsrites nervu. Imp. Slēgts Neironu ķēdes.
īstermiņa (sekundes daļas)
garš (sekundes)
Nervu centru ritmiskā darbība.
Nervu impulsu biežuma palielināšanās vai samazināšanās, kas saistīta ar sinapses īpašībām un neironu integrācijas ilgumu.
8. Nervu centru plastiskums.
Iespēja pārbūvēt īpašuma funkcionalitāti efektīvākai funkciju regulēšanai, jaunu refleksu ieviešanai, kas iepriekš nebija šim centram raksturīgi vai funkcijas atjaunošanai. Sinps slāņa centrā ir izmaiņas tirdzniecības centrā.
Uzbudināmības izmaiņas ķīmisko vielu ietekmē.
Augsta jutība pret reālām atšķirībām.
Nervu centru nogurums.
Saistīts ar augstu sinapšu nogurumu. Samazināt sajūtas. Receptori.
Centrālās nervu sistēmas koordinācijas darbības vispārīgie principi.
Bremzēšana -īpašs ner. procentiem kas izpaužas kā samazināšanās vai pilnīga izzušana resp. reakcijas.
Konverģences princips
Konverģence ir impulsu konverģence, kas nāk caur dažādiem aferentiem ceļiem jebkurā centrālajā neironā vai nervu centrā.
2 . Konverģences princips ir cieši saistīts ar principu kopīgs gala ceļš atver Sherrinkton. Daudzi dažādi stimuli var ierosināt vienu un to pašu motoro neironu un to pašu motorisko reakciju. Šis princips ir saistīts ar nevienlīdzīgu aferento un eferento ceļu skaitu.
Diverģences princips
Tas ir viena neirona kontakts ar daudziem citiem.
Apstarošana un ierosmes koncentrācija.
Tiek saukta ierosmes procesa izplatīšanās uz citiem nervu centriem apstarošana (vēlēšanu- vienā virzienā , vispārināts- plašs).
Pēc kāda laika apstarošanu aizstāj ar ierosmes koncentrācijas fenomenu tajā pašā CNS sākuma punktā.
Apstarošanas procesam ir pozitīva (jaunu kondicionētu refleksu veidošanās) un negatīva (smalku attiecību, kas izveidojušās starp ierosmes un inhibīcijas procesiem, pārkāpums, kas izraisa motora aktivitātes traucējumus) lomas.
Savstarpīguma princips (palēnina)
Dažu šūnu ierosināšana izraisa citu šūnu inhibīciju caur starpkalāru neironu.
Dominējošais princips
Ukhtomsky formulēja dominēšanas principu kā nervu centru darbības principu. jēdziens dominējošs apzīmē dominējošo centrālās nervu sistēmas ierosmes fokusu, kas nosaka ķermeņa pašreizējo aktivitāti.
Dominējošā fokusa principi :
Paaugstināta nervu centru uzbudināmība;
Uzbudinājuma uzbudinājuma noturība laika gaitā;
Spēja summēt svešus stimulus;
Inerce (spēja saglabāt uzbudinājumu ilgu laiku pēc kairinājuma darbības beigām); spēja izraisīt konjugācijas kavēšanu.

