Произходът на мозъка Савелиев Сергей Вячеславович
§ 37. Асоциативен център на мозъка на влечугите
Имайки предвид общия план на сградата нервна система, трябва да се спрем отделно на новите принципи на организация и работа на мозъка, приложени за първи път при влечугите. Нервната система на архаичните амниоти е станала логическо развитиесгради с успешен амфибиен дизайн. Мозъкът на земноводните обаче практически изпълняваше функцията на сложен рефлексен апарат и интелектуалните му способности оставаха непотърсени. Еволюцията на земноводните се определя от мускули, зъби, линейни размери и мащаби на възпроизвеждане.
От всички бозайници хората са най-развитите и развит мозък. При всички гръбначни обаче мозъкът се състои от 3 основни структурни единици: мозъчни полукълба, мозъчен ствол и малък мозък. Образуването на мрежата е основната област на обмен на информация, която е източник на мозъчна енергия. В тази мрежа нервни влакнавсеки неврон може да бъде свързан с други омрежени формати, които постоянно получават съобщения от сетивните органи, които на свой ред излъчват електрическа активност през мозъчната кора.
Той поддържа съзнание. Ако зашитата лента намали тяхната активност, човекът може да заспи или да страда от халюцинации, да вижда, чува и да има нереалистични усещания. Резултатът от сензорна депривация, когато човек е затворен в тъмна, тиха и празна стая за няколко часа, е привидно настроение, транс или кома.
Имаше елементарно развитие на хранителните ресурси, където нямаше нито място, нито биологична необходимост за развитието на сложно поведение. Със следи от този период на еволюция на гръбначните животни, ние сме изправени пред опити за развитие условни рефлексипри различни представители на съвременните земноводни. Изключително ниската способност за учене и липсата на дългосрочна памет за натрупване на индивидуален опит показват, че древните земноводни никога не са се сблъсквали със сложни поведенчески задачи в своята еволюция.
Характеристиките на развитието на сетивните органи и признаците на сложно поведение на влечугите се основават на особеностите на структурната организация на мозъка. Мозъкът на влечугите се различава от мозъка на земноводните както по количество, така и по качествено. Преди появата на амниотите поведенческите стратегии или реакциите на конкретен стимул бяха избрани според принципа на доминиране (виж Фиг. III-6, f). Този принцип се състои в това, че много първични водни гръбначни животни или земноводни нямат ясно изразен голям асоциативен център на мозъка (виж фиг. III-6, д). Изборът на форма на поведение се извършва въз основа на сравнение на дейностите на приблизително еквивалентни части на мозъка, обслужващи различни сетивни органи. Нивото на възбуждане на мозъчните аналитични центрове на един от анализаторите играе решаваща роля. Представителството на сетивния орган, достигнало най-голямо възбуждане в мозъка, става основната област за вземане на решения. След избор на една от инстинктивните реакции се извършва нейното поведенческо изпълнение. Този процес се контролира от същия просто сравнениедоминации. Ако в хода на реакцията възникне ново дразнене, което променя съотношението на възбужданията на сетивните органи, тогава поведенческата реализация на инстинктивния процес спира. Всяка конкретна ситуация е различна от предишната, но е включен същият набор от сетива. Ако се постигне най-голямо възбуждане в същата сетивна система, тогава поведението се запазва, а ако в друга, то се променя. Тъй като в естествения живот практически никога не се срещат абсолютно идентични условия, поведението дори на най-примитивните анамнии ще бъде безкрайно различно. Следователно поведението на всеки индивид ще бъде индивидуално с доста висока динамична адаптивност.
В мозъка на земноводните се появиха първите признаци на асоциативен център. При тях средният или диенцефалонът може да стане такъв център. Имаше всички основания за това. В диенцефалона има невроендокринни центрове, които контролират сексуалното поведение, миграцията и енергийния баланс на анамниотичния организъм. Чрез активирането на центровете на диенцефалона се стартират инстинктивни поведенчески програми, които контролират работата на други части на мозъка. Изглежда, че диенцефалонът може да се превърне в аналитичен център на поведението на анамнията и след това на амниотите. Но в този случай системата за осъществяване на поведенчески реакции ще се състои не само в работата на нервната система. Всеки път всяко поведенческо събитие би довело до стимулиране на работата на невроендокринните центрове. Хормоналната регулация на поведението се осъществява дълго време, а неврологичната - бързо. При бърза промяна на формите на поведение би възникнал конфликт между инертните хормонални и динамичните невронни програми на поведение. При насекомите този конфликт е решен в полза на неврохормоналните центрове и чисто инстинктивното поведение.
Доста нестабилна ситуация се е развила с първичните водни гръбначни и земноводните. От една страна, ролята на хормонално-инстинктивната регулация на поведението при земноводните е много важна и явно доминира при избора на поведенчески стратегии. От друга страна, невроморфологичният субстрат е достатъчно развит за лека индивидуализация на поведението при изпълнението на тези стратегии. Възникна оригинална система на хормонално-доминиращ избор на форми на поведение от стандартен инстинктивен набор. При земноводните поведенческата стратегия се определя от неврохормоналното състояние на индивида. Когато прилага избраната форма на поведение, тя се адаптира към конкретни условия, като използва сравнението на доминирането, което беше описано по-горе. В такава схема за управление на поведението на анамниите няма място за асоциативен център. Може да е необходимо само когато има нужда от бърза адаптивна индивидуализация на поведението. Такава ситуация може да възникне само когато последователната реализация на инстинктивните форми на поведение ще зависи пряко от постоянно променящата се ситуация.
В такива нестабилни условия заобикаляща средасе оказаха архаични влечуги. Очевидно изискванията за бърза индивидуализация на поведението и паметта са се увеличили и прилагането на хормонално доминиращия принцип на избор от стандартен инстинктивен набор от поведения е станало неефективно. Появи се напълно нов тип вземане на решения, който се е запазил в мозъците на съвременните влечуги, принадлежащи към далечни систематични групи. Всички те са обединени от едно принципно ново качество на мозъка - ясно изразен асоциативен център (виж фиг. III-7).
Основният асоциативен център на влечугите се формира в покрива на средния мозък (виж фиг. III-5, c; III-6; III-7, b). Възникна на базата на няколко сетивни органа, които имаха представителство в тази част на мозъка. Основната част от покрива на средния мозък е заета от представителството зрителна система. зрителни нервислед като преминат през хиазмата, те се пресичат и се издигат до покрива на междинния мозък. Аксоните на ганглиозните клетки на ретината завършват върху невроните на покрива на средния мозък, които са организирани в стратифицирани структури (виж Фиг. III-5, c; III-7, b). Има ясна топографска връзка между определена област на ретината и покрива на междинния мозък. В същото време формата на изображението и взаимно споразумениенеговите елементи. Дълго време предната част на покрива на средния мозък се смяташе изключително за мозъчен център. зрителен анализатор. Функционално-морфологичните изследвания обаче показват, че това далеч не е така.
Заедно с представянето на зрителната система, информацията за соматичната (кожна) чувствителност, двигателния анализатор, вестибуларните и слуховите сигнали (виж фиг. III-6, д) идва на покрива на средния мозък. слухов анализаторпри влечугите значително увеличава представителството си в този център. В резултат на това при много влечуги в задната част на покрива на средния мозък се появяват незабележими сдвоени издатини - задните или долните туберкули. Покривът на средния мозък става не хомогенна анатомична формация, както при анамния, а квадригемината. Той концентрира представянето на почти всички основни дистанционни и контактни анализатори. Дори обонятелната система има своето представителство в покрива на междинния мозък. С изключение на обонятелната система, почти всички сензорни проекции в покрива на междинния мозък на влечугите са топологични по природа. Това означава, че информацията от всяка конкретна област на тялото е представена строго определена областпокрив на междинния мозък. Запазва се принципът на телесната карта, която точково се пренася в мозъка.
По този начин в покрива на средния мозък на влечугите се концентрира разнообразна информация за състоянието на собствения им организъм и околния свят, която се комбинира според топологичния принцип. Помислете какво се случва в покрива на междинния мозък, ако предният десен крайник на влечугото просто стои на необичайна повърхност. При оценка на такава ситуация в покрива на междинния мозък, сравнителен анализсоматична, сензомоторна, слухова и зрителна информация. Това е лесно да се направи, тъй като всички сигнали са концентрирани в един център и често един върху друг, като в слоест пай. Междинният мозък провежда комплексен анализмного фактори на едно явление, което ви позволява да изберете най-подходящия отговор. Стратифицираната структура на покрива на средния мозък е идеална за това.
Изключително опростявайки реалната ситуация, можем да кажем, че в покрива на междинния мозък, представянето на различни сензорни системиразположени на условни "етажи", организирани в хоризонтална равнина. Всеки етаж е зает от своеобразна карта. Това може да бъде моментна информационна карта на рецепторни сигнали от повърхността на тялото, изображение на ретината или акустично поле. Всички тези карти са ориентирани на своите „подове“ по такъв начин, че отразяват приблизително една и съща посока в пространството. Слуховият сигнал от предния десен крайник се намира под неговата визуална карта и над соматичния сигнал от кожата на стъпалото. Специализираните „етажи“ са свързани помежду си с помощта на вертикални връзки, които ви позволяват бързо да оцените конкретна ситуацияи вземете правилното решение. Тази схема на покрива на междинния мозък ни позволява да разберем рефлексната скорост на мозъка на влечугите. Очевидно именно тази скорост е станала основната причина за еволюционния успех на архаичните влечуги.
Появата на перфектен рефлексен център за вземане на решения при влечугите имаше няколко важни последици. От една страна, способността за бърз избор на решения е безсмислена, ако общата скорост на метаболизма остане на същото ниво. Следователно развитието на междинния мозък е придружено от повишаване на метаболизма. От друга страна, увеличаването на размера на покрива на средния мозък създава необходимия клетъчен субстрат за развитие на паметта. Индивидуалният опит на животното става основа за сравняване на събития, разделени във времето. Трудно е да се надцени това събитие. За първи път се появиха реални основи за индивидуализация на поведението въз основа на сравнението на различни събития. Трябва да се отбележи, че тези невробиологични предимства на мозъка на влечугите обикновено дори не се вземат предвид, когато се реконструира ранната еволюция на амниотите (Carroll, 1982).
Архаичните влечуги, следвайки развитието на центъра за рефлексологичен анализ, получиха материален субстрат за запомняне на различни събития. Възпроизвеждането на индивидуалния опит стана достъпно за тях, което служи като основа за обучение. Земноводните вече не могат да се конкурират със самообучаващите се архаични влечуги. Хормонално-инстинктивните принципи на поведение на земноводните, рибите и безгръбначните ги превърнаха в храна за влечуги с развит рефлексно-асоциативен среден мозък.
Всички тези предимства на структурата на мозъка на влечугите не могат да възникнат сами. За такова дълбоко качествено преструктуриране на мозъка трябва да са се развили изключително сурови и необикновени условия. Архаичните влечуги трябваше да попаднат в особена среда с много високи изисквания към аналитичните свойства на мозъка и индивидуалната памет.
От книгата Находки на останки от влечуги от горната юра в шистовата мина Савелиевски автор От книгата Находки на останки от влечуги от горната юра в шистовата мина Савелиевски автор Журавлев Константин ИвановичНаходки от останки от влечуги През 1931 г., в първите месеци на добива на шисти, когато добивът все още се извършваше от открити ями, бяха получени няколко находки от останки от ихтиозавър. Сред тези находки е поредица от 13 опашни прешлена, открити в пласт нефтени шисти в основата на
автор§ 36. Общият план на структурата на нервната система на влечугите С широкомащабното развитие на сушата мозъкът на влечугите се е променил. Морфологичните му пренареждания обаче на пръв поглед не са толкова фундаментални, колкото при земноводните. Повърхностният анатомичен анализ показва, че древните
От книгата Произход на мозъка автор Савелиев Сергей Вячеславович§ 38. Условия за възникване на мозъка на влечугите. Асоциативен мозъчен център не може да възникне случайно. Енергийните разходи за поддържане на асоциативния център на мозъка и разходите за промяна на поведенческите стратегии винаги са много високи (вижте Глава I). Трябва да има причина за това
От книгата Произход на мозъка автор Савелиев Сергей Вячеславович§ 40. Адаптивно излъчване на архаични влечуги Малките хищни и всеядни влечуги, еволюирали в агресивната среда на растителни остатъци, не са най-големите животни от онова време. В края на карбона размерът на тялото им обикновено не надвишава няколко десетки
От книгата Биологична химия автор Лелевич Владимир ВалериановичЕнзимно активно място Областта на ензимна молекула, която специфично взаимодейства със субстрат, се нарича активно място. Активният център е уникална комбинация от аминокиселинни остатъци в ензимна молекула, която осигурява директно
авторКъде е центърът на масата на системата Земя-Луна? Центърът на масата на системата Земя-Луна, така нареченият барицентър, се намира на разстояние 4672 километра от центъра на Земята към Луната, тоест на дълбочина приблизително 1700 километра под повърхността на Земята. Строго погледнато, от
От книгата най-новата книгафакти. Том 1. Астрономия и астрофизика. География и други науки за земята. Биология и медицина автор Кондрашов Анатолий ПавловичКакво е центърът на удоволствието и къде се намира той в тялото? Една от частите на мозъка е хипоталамусът, който е част от диенцефалона и се намира под зрителните туберкули (таламус). Хипоталамусът, в който се намират автономните центрове
автор Кондрашов Анатолий Павлович От книгата Най-новата книга с факти. Том 1 [Астрономия и астрофизика. География и други науки за земята. биология и медицина] автор Кондрашов Анатолий Павлович От книгата Фармацевтична и хранителна мафия от Брауър ЛуисЦентър за фармакологичен надзор "Париж, 4 февруари 1977 г. NCPC, създаден през януари 1974 г., се събра на Общото събрание на 2 февруари, заедно с делегати от Националните харти на лекарите и фармацевтите, към които се присъедини група центрове за контрол на интоксикация и
От книгата В дивата природа на времето автор Чижевски Герман МихайловичКласификация и кратко описание на основните групи земноводни, влечуги, птици и бозайници КЛАС ЗЕМНОВОДНИ ИЛИ АМФИБИИ Подклас Apsidospondylus Надразред Лабиринтодонти Група земноводни, които са изчезнали не по-късно от триаския период и са достигнали своя връх в края на палеозоя
автор Кандел Ерик Ричард От книгата В търсене на паметта [Появата на нова наука за човешката психика] автор Кандел Ерик Ричард От книгата Психопати. Достоверен разказ за хора без жал, без съвест, без угризения от Кийл Кент А.Център за рехабилитация на непълнолетни в Мендота В началото на 90-те години САЩ заля истинска епидемия от тийнейджърско насилие. Броят на престъпленията, извършени от непълнолетни, почти се е удвоил между 1980 и 1993 г. Изглеждаше, че нищо не може да спре това
От книгата Четене между редовете на ДНК [Вторият код на нашия живот, или Книгата, която всеки трябва да прочете] автор Шпорк ПитърОт Берлин до центъра на бъдеща революция Не се обижда, когато го бъркат със студент или докторант. И това се случва постоянно на 32-годишния генетик. Александър Майснер - сиво-сини очи, тъмно руса небрежно сресана коса, тридневна набола - не само млада,
Име на материала:Основи на зоологията и зоогеографията
основни характеристики
Поради появата на шията при влечугите, главата е подвижно съчленена с тялото и може да се движи не само, както при земноводните, във вертикална посока, но и в хоризонтална посока.
Тялото е удължено (само при костенурките е широко), доста гъвкаво; опашката на по-голямата част от видовете е дълга, способна на различни движения; краката са силно развити (при някои видове гущери и при всички змии краката са намалени поради прехода към друг активен начин на движение). Средният размер на тялото е много по-голям от този на земноводните, а някои видове достигат огромни размери.
Корици. Кожата е покрита с рогови силни и в същото време леки люспи от ектодермален произход, които предпазват тялото не само от механични повредино и от изсъхване. Кератинизираният слой на кожата периодично се отделя по време на линеене. При костенурките и крокодилите се развиват осификации в мезенхимния слой на кожата, докато при другите влечуги те липсват. Костенурките са добре защитени от врагове с черупки. Кожата на съвременните влечуги, за разлика от земноводните, е суха, лишена от жлези.
Цветът на външните покривки е разнообразен и при много видове има защитно значение, тъй като благодарение на него животните са трудно забележими на фона на околната среда.
Нервна система. Поради по-мобилен и сложен начин на живот, нервната система на влечугите е по-добре развита от тази на земноводните (фиг. 95).
Ориз. 95. Мозък на гущер, изглед отстрани:
1 - полукълбо преден мозък; 2 - обонятелен лоб; 3 - диенцефалон; 4 - хипофизна жлеза; 5 - среден мозък; 6 - оптична хиазма (хиазма); 7 - малък мозък; 8 - гръбначен мозък
Предният мозък е значително големи размериотколкото последните (особено поради увеличаването на стриатума), се появяват на повърхността на полукълбата нервни клетки- рудиментът на мозъчната кора. Следователно влечугите бързо установяват нови връзки с външната среда от техните предци. Нараства ролята на тази част от мозъка като координатор на жизнената дейност на целия организъм. Структурата на диенцефалона и средния мозък става по-сложна, връзката им с предния мозък и други части на централната нервна система се увеличава. Малкият мозък, поради усложняването на методите на движение и необходимостта от поддържане на баланса на тялото, е много по-добре развит, отколкото при земноводните. Увеличил се е и продълговатият мозък, който образува характерна за влечугите вертикална чупка. Гръбначният мозък е дълъг; нейната структура е станала по-сложна. Това се изразява в по-рязко отделяне на външния слой (бял, т.е. пътища) от вътрешния (сив, т.е. нервни клетки) слой.
От сетивните органи при влечугите (с изключение на змиите и някои други видове) очите са по-добре развити, отколкото при земноводните: те са по-подвижни. Тяхната адаптация към зрението на различни разстояния (доста големи при много видове) се осигурява не само, както при рибите и земноводните, от движението на лещата, но и, както при висшите гръбначни, чрез промяна на нейната кривина. В допълнение към клепачите има мигаща мембрана. Редица видове имат париетален орган, свързан с диенцефалона и способен да възприема светлинни стимули. В органа на слуха са настъпили някои прогресивни промени: долната торбичка е леко увеличена; слухова костицапредава по-добре вибрациите тъпанчевъв вътрешното ухо; в допълнение към овалния прозорец се появи втори прозорец - кръгъл, което увеличава подвижността на екзолимфата, заобикаляща лабиринта; Тимпаничната мембрана не е разположена на повърхността на главата, а в малка депресия. Органите на миризмата са доста добре развити, вътрешната им повърхност се е увеличила. Вкусовите органи при повечето видове са слабо развити. Езикът играе важна роля в контакта.
Скелет. Той е почти напълно осифициран поради замяната на груби влакна костна тъкан, характерен за земноводните, тънък влакнест, здравината му се е увеличила, опорните и защитните функции са се подобрили (фиг. 96).
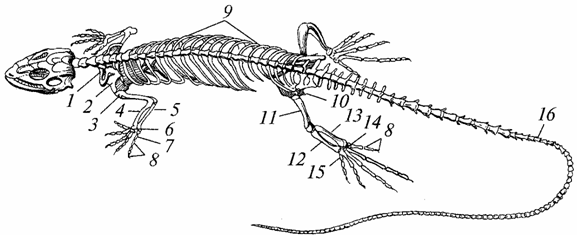
Ориз. 96. Скелет на гущер:
7 - ключица; 2 - лопатка; 3 - рамо; 4 - радиус; 5 - лакътна кост; b - китка; 7 - метакарпус; 8 - фаланги на пръстите; 9 - ребра; 10 - таз; 11 - бедро; 12 - голям пищял; 13 - фибула; 14 - тарзус; 15 - метатарзус; 16 - опашни прешлени
Черепът е увеличен; капацитетът на мозъчния участък стана по-голям от този на земноводните. Странична повърхностчерепът на древните влечуги е твърд, по-късно в него се появяват темпорални ями - една или две, което намалява масата му и увеличава повърхността за закрепване дъвкателни мускули. Долните челюсти, подобно на тези на земноводните, са окачени от мозъчната област на черепа с помощта на квадратна кост и също се състоят от няколко кости (зъбни, ставни и др.).
Тялото на първия прешлен (атлас) е прираснало под формата на зъб към тялото на втория прешлен (епистрофия). Пръстеновидният атлас получи възможност да се върти около епистрофичния зъб, поради което станаха възможни завъртанията на главата и това голямо значениеза получаване на храна (главно други животни) и за защита от врагове. Атласът е свързан с тилната област на черепа с помощта на един кондил, което също допринася за увеличаване на подвижността на главата. Гръбначният стълб е разделен на пет отдела: шиен, гръден, поясен, сакрален и опашен. Прешлени гръдниса свързани с гръдната кост чрез добре развити ребра, в резултат на което се образува гръден кош, който липсва при земноводните. Скъсените ребра се простират от лумбалните прешлени, като не достигат гръдната кост. Има два сакрални прешлена (амфибиите имат един). В опашната област има много прешлени (с изключение на костенурките).
Костите на краката са по-добре развити и по-издръжливи от тези на земноводните. Между костите на китката и тарзуса има подвижни стави. Следователно предмишницата и проксималния ред на китката, както и подбедрицата и проксималния ред на тарзуса, функционират като интегрални образувания. Особено внимание заслужава укрепването на коланите на крайниците: рамото - поради връзките му с гръдната кост и гръдния кош, тазовото - поради факта, че се опира на две сакрални прешлении двете му половини образуват две сраствания - седалищно и срамно.
Мускулна система. Претърпя диференциация, развиха се нови мускулни групи, върху които се въртят главата, движенията на шията, възникнали при влечугите, разширяване и стесняване гръден коши др.. Мускулите на краката се увеличиха значително, развиха се флексорите и екстензорите на пръстите. Масата и силата на цялата система се увеличиха.
Храносмилателната система. Той е по-диференциран, отколкото при земноводните (фиг. 97). Зъбите са съсредоточени главно върху челюстите, броят им е голям, но те са еднотипни и служат главно за улавяне на плячка. Слюнчените жлези са по-добре развити, отколкото при земноводните (при редица видове секретите на тези жлези са отровни). При влечугите започва процесът на образуване на твърдо небце, което е особено развито при крокодилите. Поради образуването на твърдо небце, устната кухина е отделена от назофарингеалната кухина, което улеснява както дишането, така и усвояването на храната. Фаринкса, хранопровода и стомаха са добре развити (особено при хищници, които се хранят с гръбначни). Черният дроб (с жлъчния мехур) и панкреасът са по-съвършени от тези на земноводните. Тънко червозначително удължено, дебелото черво е късо и завършва с клоака. Съвременните влечуги се хранят предимно с животни, броят на тревопасните видове е незначителен.
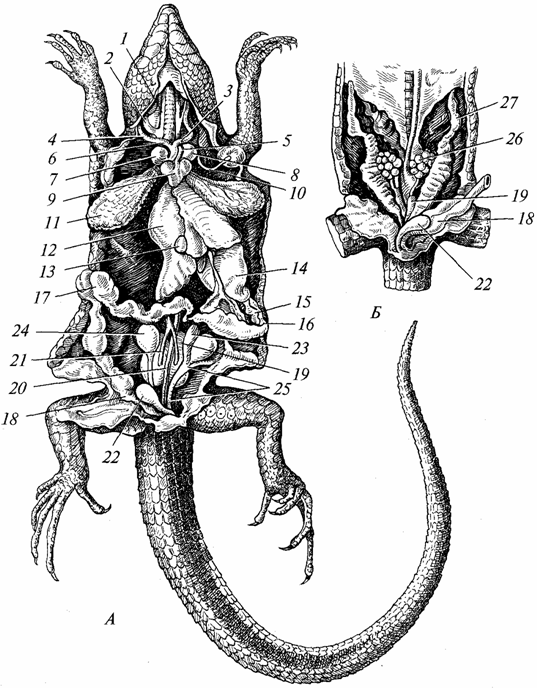
Ориз. 97. Вътрешни органигущери; Мъжки; B - женски:
/ - външни и 2 - вътрешни югуларни вени; 3 - лява и 4 - дясна каротидна артерия; 5 - лява и 6 - дясна аортна дъга; 7 - дясно и 8 - ляво предсърдие; 9 - вентрикул на сърцето; 10 - лява субклавиална артерия 11 - бял дроб; 12 - черен дроб; 13 - жлъчен мехур; 14 - стомаха; /5 - панкреас; 16 - дванадесетопръстника; 17 - дебело черво 18 - ректум; 19 - бъбрек; 20 - дорзална аорта; 21 - еферентни вени на бъбреците; 22 - пикочен мехур; 23 - придатък на тестиса; 24 - тестис; 25 - семенна тръба; 26 - яйчник; 27 - яйцепровод
Дихателната система. Дихателните органи на влечугите са белите дробове (виж фиг. 97), тъй като кожата им е неподходяща за обмен на газ; лигавица устната кухина, който играе важна роля в дишането на земноводните, се използва за изпълнение на тази функция от няколко вида влечуги. Поради тази причина и особено във връзка с повишената активност на влечугите в сравнение с земноводните, структурата на белите дробове и респираторен трактсложно. Белите дробове са по-големи органи и са разположени по-дълбоко в гръдната кухина, отколкото при земноводните. Тяхната вътрешна повърхност се е увеличила значително поради развитието върху нея на много напречни греди с по-плътна мрежа. кръвоносни съдове. Въздухът навлиза в белите дробове, преминавайки през хоаните, фаринкса, ларинкса, дългата трахея, чийто лумен се поддържа от хрущялни пръстени. По време на преминаване през трахеята въздухът се изчиства от прах, температурата му се доближава до телесната. При земноводните трахеята е била в начален стадий. Задният край на трахеята е разделен на два бронха, в стените на които също има хрущялни пръстени. Изпълнението на вдишване и издишване е по-ефективно, отколкото при земноводните, поради развитието на гръдния кош при влечугите, чийто обем се увеличава с вдишване и намалява с издишване. Същият механизъм се е запазил и при висшите гръбначни животни – птици и бозайници.
Кръвоносна система. Сърцето, подобно на земноводните, е трикамерно, но отдолу във вентрикула се появява зародиш на преграда. При крокодилите тя вече е пълна и сърцето им става четирикамерно (две предсърдия и две вентрикули). Следователно при влечугите разделянето на венозния и артериалния кръвен поток е засилено, но пълната изолация на двата потока все още не е пълна дори при крокодилите, както може да се види от следващото описание (фиг. 98).
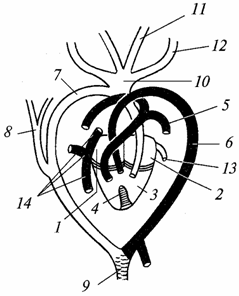
Ориз. 98. Сърцето и основните съдове на гущера (светли - съдове с артериална кръв, засенчване - със смесена кръв; черни - с венозна):
1 - дясно предсърдие; 2 - ляво предсърдие; 3 - вентрикул; 4 - непълна преграда, разделяща вентрикула на лява и дясна половина; 5 - белодробна артерия; 6 - лява аортна дъга; 7 - дясна аортна дъга; 8 - субклавиална артерия; 9 - дорзална аорта; 10 - общ багажниккаротидни артерии; 11 - ляв вътрешен каротидна артерия, 12 - лява външна каротидна артерия; 13 - белодробна вена; 14 - гръб Главна артерияи предна вена кава (югуларна) вени
Артериалната кръв навлиза в лявото предсърдие - идва от белите дробове. Венозната кръв навлиза в дясното предсърдие - тя се събира от всички части на тялото. Освен това кръвта от двете предсърдия навлиза във вентрикула: в дясната му половина - венозна, в лявата - артериална. Поради наличието на преграда, макар и непълна, кръвта във вентрикула се смесва по-малко, отколкото при земноводните. Няма артериален конус и главните артериални стволове се отклоняват директно от сърцето.
от правилната странавентрикулите се отклоняват белодробни артерии(бившата четвърта двойка бранхиални артерии), пренасяща венозна кръв към белите дробове. Бившата трета двойка бранхиални съдове атрофира. Втората двойка съдове се състои от две аортни дъги: дясната дъга започва от лявата страна на вентрикула и носи артериална кръв, лявата дъга започва от дясната страна на вентрикула (но по-близо до лявата му страна) и носи смесена кръв . От дясната дъга на аортата каротидните артерии (бившата първа двойка хрилни съдове) се простират до главата. Те снабдяват мозъка, особено чувствителен към липса на кислород, с артериална кръв. Двете аортни дъги се съединяват, за да образуват аортата, която се разклонява на много артерии. По този начин тялото на влечугото се доставя предимно със смесена кръв, но е наситено с повече кислород от това на земноводните, което допринася за по-интензивен метаболизъм.
отделителна система. Тя се е променила много поради повишената скорост на метаболизма на влечугите, което доведе до увеличаване на количеството на продуктите на дисимилация и пълното им адаптиране към сухоземен начин на живот, в резултат на което стана необходимо по-икономичното използване на водата, която влиза тялото. Следователно влечугите са развили по-сложни отделителни органи, наречени тазови или вторични бъбреци (metayaephros). Всички тубули на тези бъбреци нямат фунии и започват с капсулите на Боуман, заобикалящи малпигиевите тела с увеличен брой капиляри. Тубулите на тазовите бъбреци са много дълги и гъсто обвити с капиляри. В капсулите на Bowman от продуктите на дисимилация, разтворени във вода и някои органична материя(напр. глюкоза) се образува първична урина. Последният, преминавайки през дългите бъбречни тубули, връща в кръвта през капилярите, оплетени стените им, значителна част от водата и други вещества, необходими на тялото. Образуваната след това вторична урина не излиза през каналите на вълците, а през новоразвитите уретери в клоаката и се натрупва в пикочен мехур. Процесите на образуване на първична и вторична урина също се извършват в стволните бъбреци, но с връщане на по-малко количество вода в кръвта. Първичните водни гръбначни животни, особено земноводните, чиято кожа е гола, често страдат от излишък на вода, а не от липса. Трябва също да се отбележи, че при влечугите крайният продукт от разграждането на протеина е пикочна киселина, за чието извеждане от организма с урината се изразходва малко вода. При земноводните и редица други първични водни гръбначни разграждането на протеините води до образуването на урея, отстраняването на която изисква много вода.
Ембрионите на влечугите имат зачатък на главния бъбрек и развит бъбрек на багажника, който функционира при възрастни (при редица видове гущери) до пубертета. Тези факти потвърждават приемствеността на трите вида бъбреци по време на еволюцията на гръбначните животни.
Възпроизвеждане. Мъжката репродуктивна система на влечугите включва два тестиса с кръгла форма, разположени отстрани на лумбалния гръбначен стълб, два придатка на тестисите, образувани от предните части на бъбреците на багажника, два семенни канала (Wolffian канали) и копулаторни органи, развити от стената на клоаката. Дамски репродуктивна системасе състои от два яйчника, които нарастват силно по време на размножителния период, и два яйцепровода (Мюлерови канали), които се вливат в клоаката. Оплождането е само вътрешно. Семенната течност се въвежда от мъжките в клоаката на женската с помощта на копулационни органи. Зрелите яйца, съдържащи много жълтък, попадат във фуниите на яйцепроводите. Свързването на женските и мъжките гамети се извършва в горната част на яйцепроводите, където сперматозоидите проникват след полов акт. Зиготите, движещи се по яйцепроводите, в средната част на последния са заобиколени от протеинова обвивка, а отзад са покрити с черупка.
При някои видове гущери и змии се наблюдава партеногенетично размножаване на женските. При тези видове обикновено няма мъже.
развитие. При влечугите развитието е директно; влечугите нямат ларви. Оплодените яйца обикновено се отлагат от влечуги в почвата, а някои видове - под гниещи растителни остатъци. При редица видове развитието на зиготите се извършва в гениталния тракт на женската и веднага след освобождаването на развитите яйца от тях се излюпват формирани животни, т.е. яйцевидността е характерна за такива видове. При няколко вида се наблюдава истинско живо раждане: съдовете на жълтъчната торбичка са тясно свързани със съдовете на яйцепровода и ембрионът получава значителна част от хранителни веществаот тялото на майката. Яйценосните се наблюдават по-често при видове, разпространени в северните и планинските райони. Очевидно ововивипаритетът е защитата на развиващите се ембриони от охлаждане и други неблагоприятни условия. Няма нито един вид влечуги, при които яйцата да се развиват във вода. Дори и при влечугите провеждане повечетоживотът във водата (всички крокодили, много костенурки, някои змии), яйцеполагането и развитието се извършват само на сушата. Морските змии, които никога не напускат водата, са живородни.
Яйцата и ембрионите на влечугите имат редица адаптации за развитие на сушата. Първо, яйцата се покриват с черупки и други мембрани, които ги предпазват от изсъхване и повреда. Второ, ембрионите развиват специални ембрионални мембрани (серозна, амнионна и алантоисна), които са от голямо значение за осигуряване на ембрионално развитиеживотни на сушата. Серозата и амнионът се развиват от една и съща двойна гънка на тялото на ембриона, произхождаща от ектодермата и мезодермата. Тази гънка се появява от две противоположни странипод формата на рудименти, които растат един към друг и се затварят над ембриона. Външната част на гънката се превръща в сероза (външният й слой е ектодермален, вътрешният е мезодермален), а вътрешната част се превръща в амнион (външният му слой е мезодермален, вътрешният е ектодермален). Кухината, образувана от амниона, е пълна с водниста течност, поради което развитието протича във влажна среда и ембрионът е защитен от изсушаване и въздействието на различни разклащания, особено опасни във въздуха.
Произход. Първите влечуги, възникнали през втората половина на карбона, са живели във влажни земни биотопи и вероятно са се размножавали и развивали във вода. По това време земята вече е обитавана от по-ниски земни растения, различни безгръбначни, започва завладяването на въздуха от насекоми. Влечугите, които можеха да пътуват по-далеч от земноводните от водни тела, имаха изобилна храна и не срещаха сериозни врагове. В края на карбона и многократно през перма климатът става по-сух, което причинява смъртта на много земноводни и, обратно, допринася за прогресивното развитие и по-широкото разпространение на влечугите на сушата. След известно време те преминаха към размножаване и развитие извън водоемите, първо в по-влажни сухоземни биотопи, а след това в по-сухи места. През мезозойската ера те се разпространяват по цялото земно кълбо и много различни групи. Наред с хищниците "имаше много растителноядни видове, влечуги, адаптирани към полет, и влечуги, живеещи в моретата. Размерът на тялото им беше различен, но повечето бяха големи животни, а някои бяха гигантски - до 20 м или повече на дължина. Климат през почти цялата мезозойска ера е била топла, много благоприятна за тези животни, които нямат достатъчно висока постоянна температуратялото и губят своята активност, когато околната среда се охлади.
В края на мезозойската ера, Глобусътпротичат мощни планиностроителни процеси, съпроводени с рязка промяна в условията на съществуване (температура, влажност и др.). Това доведе до масовото изчезване на мезозойските влечуги, които не издържаха на борбата за съществуване в такива трудни за тях абиотични условия на околната среда, усложнени от конкуренцията с бозайници и птици. В същото време през кайнозойската ера възникват и стават многобройни нови групи влечуги - от тях произлизат гущери и змии. От старите групи е оцелял само един много примитивен вид първични гущери, малък брой крокодили и костенурки. Съвременните влечуги са разпространени главно в топлите страни, особено в субтропичните и тропическите страни, те са малко в страните с умерен климат и липсват в далечния север.

