Aivojen alkuperä Saveliev Sergey Vyacheslavovich
§ 37. Matelijoiden aivojen assosiaatiokeskus
Ottaen huomioon rakennuksen yleiskaava hermosto, meidän tulisi tarkastella erikseen aivojen organisoinnin ja toiminnan uusia periaatteita, jotka otettiin käyttöön ensin matelijoissa. Arkaaisten lapsivesien hermosto on muuttunut looginen kehitys onnistuneen amfibiosuunnittelun rakennuksia. Sammakkoeläimen aivot suorittivat kuitenkin käytännössä monimutkaisen refleksilaitteen tehtävän, ja sen älylliset kyvyt jäivät vaatimatta. Sammakkoeläinten kehityksen päättivät lihakset, hampaat, lineaariset mitat ja lisääntymisasteikot.
Kaikista nisäkkäistä ihminen on kehittynein ja kehittyneet aivot. Kuitenkin kaikilla selkärankaisilla aivot koostuvat kolmesta päärakenneyksiköstä: aivopuoliskoista, aivorungosta ja pikkuaivoista. Verkon muodostus on tiedonvaihdon pääalue, joka on aivoenergian lähde. Tässä verkossa hermokuituja jokainen neuroni voidaan liittää muihin ristiinsidottuihin formaatteihin, jotka vastaanottavat jatkuvasti viestejä aistielimistä, jotka puolestaan lähettävät sähköistä aktiivisuutta aivokuoren kautta.
Hän ylläpitää tietoisuutta. Jos ommeltu nauha vähentää hänen aktiivisuuttaan, henkilö voi nukahtaa tai kärsiä hallusinaatioista, nähdä, kuulla ja tuntea epärealistisia tuntemuksia. Sensorisen puutteen seurauksena, kun henkilö on lukittu pimeään, hiljaiseen ja tyhjään huoneeseen useiksi tunteiksi, on näennäinen mieliala, transsi tai kooma.
Elintarvikeresursseja kehitettiin alkeellisella tasolla, jolloin monimutkaisen käyttäytymisen kehittymiselle ei ollut tilaa eikä biologista tarvetta. Tämän selkärankaisten evoluution ajanjakson jälkiä vastaan meidän on yritettävä kehittyä ehdolliset refleksit monissa nykyaikaisten sammakkoeläinten edustajissa. Äärimmäisen alhainen oppimiskyky ja pitkän aikavälin muistin puute yksilöllisen kokemuksen keräämiseksi osoittavat, että muinaiset sammakkoeläimet eivät koskaan kohdanneet monimutkaisia käyttäytymistehtäviä evoluution aikana.
Aistielinten kehityksen piirteet ja matelijoiden monimutkaisen käyttäytymisen merkit perustuvat aivojen rakenteellisen organisaation ominaisuuksiin. Matelijoiden aivot eroavat sammakkoeläinten aivoista sekä määrältään että määrältään laadullisesti. Ennen lapsivesien tuloa käyttäytymisstrategiat tai vasteet tiettyyn ärsykkeeseen valittiin dominanssiperiaatteen mukaisesti (katso kuva III-6, f). Tämä periaate koostuu siitä tosiasiasta, että monilla primaarisilla vedessä elävillä selkärankaisilla tai sammakkoeläimillä ei ole voimakasta suurta aivojen assosiaatiokeskusta (katso kuva III-6, e). Käyttäytymismuodon valinta tapahtuu eri aistielimiä palvelevien aivojen suunnilleen vastaavien osien toiminnan vertailun perusteella. Yhden analysaattorin aivojen analyyttisten keskusten viritystasolla on ratkaiseva rooli. Aivojen suurimman jännityksen saavuttaneen aistielimen esityksestä tulee pääasiallinen päätöksenteko-alue. Kun olet valinnut yhden vaistomaisista reaktioista, sen käyttäytyminen toteutetaan. Tätä prosessia ohjaa sama yksinkertainen vertailu dominanssit. Jos reaktion aikana syntyy uusi ärsytys, joka muuttaa aistielinten viritysten suhdetta, vaistonvaraisen prosessin käyttäytyminen pysähtyy. Jokainen tilanne on erilainen kuin edellinen, mutta samat aistit ovat mukana. Jos suurin heräte saavutetaan samassa aistijärjestelmässä, käyttäytyminen säilyy, ja jos toisessa, se muuttuu. Koska täysin identtisiä olosuhteita ei käytännössä koskaan löydy luonnollisesta elämästä, alkeellisimpienkin anamnioiden käyttäytyminen on äärettömän vaihtelevaa. Näin ollen jokaisen yksilön käyttäytyminen on yksilöllistä ja sen dynaaminen sopeutumiskyky on melko korkea.
Ensimmäiset merkit assosiaatiokeskuksesta ilmestyivät sammakkoeläinten aivoihin. Niissä keskiosasta tai aivokalvosta voisi tulla tällainen keskus. Tähän oli kaikki syyt. Välilihassa on neuroendokriiniset keskukset, jotka säätelevät sukupuolikäyttäytymistä, vaellusta ja anamnioottisen organismin energiatasapainoa. Väliaivokeskusten aktivoitumisen kautta käynnistetään vaistomaisia käyttäytymisohjelmia, jotka ohjaavat muiden aivojen osien toimintaa. Vaikuttaa siltä, että välikalvosta voisi tulla anamnian ja sitten lapsivesien käyttäytymisen analyyttinen keskus. Tässä tapauksessa käyttäytymisreaktioiden toteuttamisjärjestelmä ei kuitenkaan koostu pelkästään hermoston työstä. Joka kerta mikä tahansa käyttäytymistapahtuma johtaisi neuroendokriinisten keskusten toiminnan stimulaatioon. Hormonaalinen käyttäytymisen säätely toteutetaan pitkään ja neurologinen - nopeasti. Käyttäytymismuotojen nopean muutoksen myötä syntyisi konflikti inerttien hormonaalisten ja dynaamisten käyttäytymisohjelmien välillä. Hyönteisissä tämä ristiriita on ratkaistu neurohormonaalisten keskusten ja puhtaasti vaistomaisen käyttäytymisen hyväksi.
Vedessä elävien primääriselkärankaisten ja sammakkoeläinten tilanne on muodostunut melko epävakaaksi. Toisaalta sammakkoeläinten käyttäytymisen hormonaalisesti vaistomaisen säätelyn rooli on erittäin tärkeä ja selvästi hallitseva käyttäytymisstrategioiden valinnassa. Toisaalta neuromorfologinen substraatti on riittävän kehittynyt käyttäytymisen lievää yksilöllistämistä varten näiden strategioiden toteutuksessa. Syntyi alkuperäinen hormonaalisesti hallitseva käyttäytymismuotojen valinta vakiomuotoisesta vaistomaisesta ryhmästä. Sammakkoeläimissä käyttäytymisstrategian määrää yksilön neurohormonaalinen tila. Toteuttaessaan valittua käyttäytymismuotoa se mukautuu tiettyihin olosuhteisiin edellä kuvatun dominanssivertailun avulla. Tällaisessa anamnion käyttäytymisen hallintajärjestelmässä ei ole tilaa assosiatiiviselle keskukselle. Sitä voitaisiin tarvita vain silloin, kun tarvitaan nopeaa mukautuvaa käyttäytymisen yksilöllistämistä. Tällainen tilanne voi syntyä vain, kun vaistomaisten käyttäytymismuotojen johdonmukainen toteutuminen riippuu suoraan jatkuvasti muuttuvasta tilanteesta.
Tällaisissa epävakaissa olosuhteissa ympäristöön osoittautuivat arkaaisiksi matelijoiksi. Ilmeisesti vaatimukset käyttäytymisen ja muistin nopealle yksilöllistymiselle ovat lisääntyneet, ja hormonidominoivan periaatteen toteuttaminen valita tavanomaisesta vaistomaisesta käyttäytymisjoukosta on tullut tehottomaksi. On syntynyt täysin uudenlainen päätöksenteko, joka on säilynyt kaukaisiin systemaattisiin ryhmiin kuuluvien nykyaikaisten matelijoiden aivoissa. Niitä kaikkia yhdistää yksi pohjimmiltaan uusi aivojen laatu - selkeä assosiaatiokeskus (katso kuva III-7).
Matelijoiden tärkein assosiaatiokeskus muodostui keskiaivojen katolle (katso kuva III-5, c; III-6; III-7, b). Se syntyi useiden aistielinten perusteella, joilla oli edustus tässä aivojen osassa. Suurin osa keskiaivojen katosta on edustuksen käytössä visuaalinen järjestelmä. näköhermoja kulkiessaan chiasmatan ne ylittävät ja nousevat keskiaivojen katolle. Verkkokalvon gangliosolujen aksonit päättyvät keskiaivojen katon hermosoluihin, jotka ovat järjestäytyneet kerrostuneiksi rakenteiksi (katso kuva III-5, c; III-7, b). Verkkokalvon tietyn alueen ja väliaivojen katon välillä on selvä topografinen yhteys. Samalla kuvan muoto ja keskinäinen järjestely sen elementtejä. Väliaivojen katon etuosaa pidettiin pitkään yksinomaan aivokeskuksena. visuaalinen analysaattori. Funktionaalis-morfologiset tutkimukset ovat kuitenkin osoittaneet, että näin ei suinkaan ole.
Näköjärjestelmän esityksen mukana väliaivojen katolle tulee tietoa somaattisesta (ihon) herkkyydestä, motorisesta analysaattorista, vestibulaarisista ja kuulosignaaleista (ks. kuva III-6, e). kuuloanalysaattori matelijoilla se lisää merkittävästi edustustaan tässä keskustassa. Seurauksena on, että monissa matelijoissa keskiaivojen katon takaosassa näkyy huomaamattomia parillisia ulkonemia - taka- tai alempia mukuloita. Väliaivojen katosta ei tule homogeenista anatomista muodostumaa, kuten anamniassa, vaan nelijalkainen. Se keskittää lähes kaikkien tärkeimpien kauko- ja kontaktianalysaattoreiden esityksen. Jopa hajujärjestelmällä on edustus keskiaivojen katossa. Hajujärjestelmää lukuun ottamatta lähes kaikki aistinvaraiset projektiot matelijoiden keskiaivojen kattoon ovat luonteeltaan topologisia. Tämä tarkoittaa, että tiedot jokaiselta kehon tietyltä alueelta esitetään tiukasti tiettyä aluetta keskiaivojen katto. Kehokartan periaate säilyy, joka siirtyy pisteittäin aivoihin.
Siten matelijoiden keskiaivojen kattoon keskittyy monenlaista tietoa oman organismin tilasta ja ympäröivästä maailmasta, joka yhdistetään topologisen periaatteen mukaisesti. Mieti, mitä tapahtuu keskiaivojen katolla, jos matelijan oikea eturaaja yksinkertaisesti seisoo epätavallisella pinnalla. Kun arvioidaan tällaista tilannetta keskiaivojen katossa, vertaileva analyysi somaattinen, sensorimotorinen, kuulo- ja visuaalinen informaatio. Tämä on helppo tehdä, koska kaikki signaalit ovat keskittyneet yhteen keskustaan ja usein päällekkäin, kuten kerroksisessa piiraassa. Keskiaivot johtavat monimutkainen analyysi yhden ilmiön monet tekijät, joiden avulla voit valita sopivimman vastauksen. Keskiaivokaton kerrostettu rakenne on ihanteellinen tähän.
Yksinkertaistaen äärimmäisen todellista tilannetta, voimme sanoa, että keskiaivojen katossa on esitetty erilaisia aistijärjestelmät sijaitsevat ehdollisissa "lattioissa", jotka on järjestetty vaakatasoon. Jokainen kerros on eräänlainen kartta. Se voi olla hetkellinen tietokartta kehon pinnalta tulevista reseptorisignaaleista, kuva verkkokalvolla tai akustinen kenttä. Kaikki nämä kartat on suunnattu "lattioilleen" siten, että ne heijastavat suunnilleen samaa suuntaa avaruudessa. Oikean eturaajan kuulosignaali on sen visuaalisen kartan alla ja jalan ihon somaattisen signaalin yläpuolella. Erikoistuneet "lattiat" korreloidaan keskenään pystysuuntaisten linkkien avulla, joiden avulla voit arvioida nopeasti erityinen tilanne ja tehdä oikea päätös. Tämä keskiaivojen katon kaavio antaa meille mahdollisuuden ymmärtää matelijoiden aivojen refleksinopeutta. Ilmeisesti juuri tästä nopeudesta tuli tärkein syy arkaaisten matelijoiden evoluution menestykselle.
Täydellisen refleksin päätöksentekokeskuksen syntymisellä matelijoille oli useita tärkeitä seurauksia. Toisaalta kyky valita nopeasti ratkaisuja on merkityksetöntä, jos kokonaisaineenvaihdunta pysyy samalla tasolla. Tämän seurauksena keskiaivojen kehittymiseen liittyi aineenvaihdunnan lisääntyminen. Toisaalta keskiaivojen katon koon kasvu loi tarvittavan solusubstraatin muistin kehittämiseen. Eläimen yksilöllinen kokemus oli perusta ajallisesti erotettujen tapahtumien vertailulle. Tätä tapahtumaa on vaikea yliarvioida. Ensimmäistä kertaa syntyi todellista perustaa käyttäytymisen yksilöimiselle erilaisten tapahtumien vertailun perusteella. On huomattava, että näitä matelijoiden aivojen neurobiologisia etuja ei yleensä edes oteta huomioon, kun rekonstruoidaan amnioottien varhaista kehitystä (Carroll, 1982).
Arkaaiset matelijat saivat refleksologisen analyysin keskuksen kehittymisen jälkeen materiaalisubstraatin erilaisten tapahtumien muistamiseen. Heidän käyttöönsä tuli yksilöllisen kokemuksen uudelleentuotanto, joka toimii oppimisen perustana. Sammakkoeläimet eivät enää pystyneet kilpailemaan itseoppivien arkaaisten matelijoiden kanssa. Sammakkoeläinten, kalojen ja selkärangattomien hormonaalisesti vaistomaiset käyttäytymisperiaatteet tekivät niistä ruokaa matelijoille, joilla on kehittynyt refleksi-assosiatiivinen keskiaivo.
Kaikki nämä matelijoiden aivojen rakenteen edut eivät voineet syntyä itsestään. Tällaista syvää aivojen laadullista rakennemuutosta varten on täytynyt kehittyä erittäin ankaria ja poikkeuksellisia olosuhteita. Arkaaisten matelijoiden täytyi löytää itsensä erikoisesta ympäristöstä, jossa aivojen analyyttisille ominaisuuksille ja yksilölliselle muistille asetettiin erittäin korkeat vaatimukset.
Kirjasta Yläjurakauden matelijoiden jäännökset Saveljevskin liuskekaivoksessa kirjoittaja Kirjasta Yläjurakauden matelijoiden jäännökset Saveljevskin liuskekaivoksessa kirjoittaja Žuravlev Konstantin IvanovitšMatelijoiden jäännöksiä Vuonna 1931, liuskeen louhinnan ensimmäisinä kuukausina, kun louhintaa tehtiin vielä avolouhoksilla, löydettiin useita ikthyosaurusten jäänteitä. Näiden löytöjen joukossa on 13 hännän nikaman sarja, joka löydettiin öljyliuskekerroksesta
kirjoittaja§ 36. Matelijoiden hermoston rakenteen yleissuunnitelma Matelijoiden laajamittaisen kehityksen myötä matelijoiden aivot ovat muuttuneet. Sen morfologiset uudelleenjärjestelyt eivät kuitenkaan ensi silmäyksellä olleet yhtä tärkeitä kuin sammakkoeläimissä. Pinnallinen anatominen analyysi osoittaa, että muinainen
Kirjasta Origin of the Brain kirjoittaja Saveliev Sergei Vjatšeslavovitš§ 38. Edellytykset matelijoiden aivojen syntymiselle Assosiatiivinen aivokeskus ei voi syntyä sattumalta. Aivojen assosiatiivisen keskuksen ylläpitämisen energiakustannukset ja käyttäytymisstrategioiden muuttaminen ovat aina erittäin korkeat (katso luku I). Siihen täytyy olla syynsä
Kirjasta Origin of the Brain kirjoittaja Saveliev Sergei Vjatšeslavovitš§ 40. Arkaaisten matelijoiden mukautuva säteily Kasvijätteen aggressiivisessa ympäristössä kehittyneet pienet peto- ja kaikkisyöjämatelijat eivät olleet tuon ajan suurimpia eläimiä. Hiilen lopussa heidän ruumiinsa koko ei yleensä ylittänyt useita kymmeniä
Kirjasta Biological Chemistry kirjoittaja Lelevich Vladimir ValeryanovitšEntsyymiaktiivinen kohta Entsyymimolekyylin aluetta, joka on spesifisesti vuorovaikutuksessa substraatin kanssa, kutsutaan aktiiviseksi paikaksi. Aktiivinen kohta on ainutlaatuinen aminohappotähteiden yhdistelmä entsyymimolekyylissä, joka tarjoaa suoran
kirjoittajaMissä on Maa-Kuu -järjestelmän massakeskus? Maa-Kuu -järjestelmän massakeskus, ns. barycenter, sijaitsee 4672 kilometrin etäisyydellä Maan keskustasta Kuuhun, eli noin 1700 kilometrin syvyydessä maan pinnan alapuolella. Tarkkaan ottaen
Kirjasta uusin kirja tosiasiat. Osa 1. Tähtitiede ja astrofysiikka. Maantiede ja muut maantieteet. Biologia ja lääketiede kirjoittaja Kondrashov Anatoli PavlovichMikä on nautintokeskus ja missä se sijaitsee kehossa? Yksi aivojen osista on hypotalamus, joka on osa välilihasta ja sijaitsee visuaalisten tuberkuloiden (talamuksen) alla. Hypotalamus, jossa autonomiset keskukset sijaitsevat
kirjoittaja Kondrashov Anatoli Pavlovich Kirjasta Uusin tosiasioiden kirja. Osa 1 [Astronomia ja astrofysiikka. Maantiede ja muut maantieteet. Biologia ja lääketiede] kirjoittaja Kondrashov Anatoli Pavlovich Kirjasta Pharmaceutical and Food Mafia kirjoittanut Brower LouisFarmakologisen valvonnan keskus "Pariisi, 4. helmikuuta 1977. Tammikuussa 1974 perustettu NCPC kokoontui yleiskokouksessa 2. helmikuuta yhdessä kansallisten lääkärien ja farmaseuttien peruskirjan edustajien kanssa, joihin liittyi joukko valvontakeskuksia päihtymys ja
Kirjasta Ajan erämaissa kirjoittaja Chizhevsky saksalainen MihailovichSammakkoeläinten, matelijoiden, lintujen ja nisäkkäiden pääryhmien luokittelu ja lyhyt kuvaus LUOKKA Sammakkoeläimet TAI sammakkoeläimet Alaluokka Apsidospondylus Superorder Labyrinthodonts Ryhmä sammakkoeläimiä, jotka kuolivat sukupuuttoon viimeistään triaskaudella ja saavuttivat huippunsa paleozokauden lopussa
kirjoittaja Kandel Eric Richard Kirjasta In Search of Memory [The Emergence of a New Science of the Human Psyche] kirjoittaja Kandel Eric Richard Kirjasta Psykopaatit. Luotettava tarina ihmisistä ilman sääliä, ilman omaatuntoa, ilman katumusta Kirjailija: Keel Kent A.Mendota Juvenile Rehabilitation Center 1990-luvun alussa todellinen teiniväkivallan epidemia pyyhkäisi Yhdysvaltoihin. Nuorten tekemien rikosten määrä lähes kaksinkertaistui vuosien 1980 ja 1993 välillä. Tuntui, ettei mikään voinut estää tätä
Kirjasta Reading between the lines of DNA [Elämämme toinen koodi eli kirja, joka jokaisen tulee lukea] kirjailija Shpork PeterBerliinistä tulevan vallankumouksen keskustaan Hän ei loukkaannu, kun hänet luullaan opiskelijaksi tai tohtoriksi. Ja tätä tapahtuu 32-vuotiaalle geneetikolle koko ajan. Alexander Meissner - harmaansiniset silmät, tummanvaaleat huolimattomasti kammatut hiukset, kolmen päivän sänki - ei vain nuori,
Materiaalin nimi: Eläintieteen ja eläingeografian perusteet
Yleiset luonteenpiirteet
Matelijoiden kaulan ulkonäön vuoksi pää on liikkuvasti nivelletty vartalon kanssa ja voi liikkua, kuten sammakkoeläimissä, pystysuunnassa, mutta myös vaakasuunnassa.
Runko on pitkänomainen (vain kilpikonnilla leveä), melko joustava; suurimman osan lajeista häntä on pitkä, kykenee erilaisiin liikkeisiin; jalat ovat voimakkaasti kehittyneet (joissakin liskolajeissa ja kaikissa käärmeissä jalat ovat pienentyneet siirtymisen vuoksi toiseen aktiiviseen liikkumismuotoon). Keskimääräinen ruumiinkoko on paljon suurempi kuin sammakkoeläinten, ja jotkut lajit saavuttavat valtavia kokoja.
Kannet. Iho on peitetty kiimainen vahvoilla ja samalla kevyillä ektodermaalista alkuperää olevilla suomuilla, jotka suojaavat kehoa paitsi mekaanisia vaurioita vaan myös kuivumisesta. Ihon keratinisoitunut kerros irtoaa ajoittain sulamisen aikana. Kilpikonnilla ja krokotiileilla luustumiset kehittyvät ihon mesenkymaalisessa kerroksessa, kun taas muissa matelijoissa niitä ei ole. Kilpikonnat ovat hyvin suojattuja vihollisilta kuorilla. Nykyaikaisten matelijoiden iho, toisin kuin sammakkoeläinten, on kuiva, ilman rauhasia.
Ulkokuorien väri on monipuolinen ja sillä on monilla lajeilla suojaava arvo, koska sen ansiosta eläimet ovat tuskin havaittavissa ympäristön taustalla.
Hermosto. Liikkuvamman ja monimutkaisemman elämäntavan ansiosta matelijoiden hermosto on paremmin kehittynyt kuin sammakkoeläinten (kuva 95).
Riisi. 95. Liskon aivot, sivukuva:
1 - puolipallo etuaivot; 2 - hajulohko; 3 - aivokalvon; 4 - aivolisäke; 5 - keskiaivot; 6 - optinen kiasmi (chiasm); 7 - pikkuaivot; 8 - selkäydin
Etuaivot ovat merkittävästi suuret koot kuin jälkimmäiset (erityisesti striatumin lisääntymisen vuoksi), näkyvät pallonpuoliskojen pinnalla hermosolut- aivokuoren alkuaine. Siksi matelijat muodostavat nopeasti uusia yhteyksiä ulkoiseen ympäristöön kuin esi-isänsä. Tämän aivojen osan rooli koko organismin elintärkeän toiminnan koordinaattorina kasvaa. Väliaivojen ja väliaivojen rakenne monimutkaistuu, niiden yhteys etuaivojen ja keskushermoston muihin osiin lisääntyy. Pikkuaivot ovat liikkumismenetelmien monimutkaisuuden ja kehon tasapainon ylläpitämisen tarpeen vuoksi paljon paremmin kehittyneet kuin sammakkoeläimillä. Myös medulla oblongata on lisääntynyt, mikä muodostaa matelijoille ominaisen pystysuoran mutkan. Selkäydin on pitkä; sen rakenteesta on tullut monimutkaisempi. Tämä ilmeni ulomman kerroksen (valkoinen, ts. reitit) terävämmällä erottelulla sisemmästä (harmaa, ts. hermosolut) kerroksesta.
Matelijoiden aistielimistä (lukuun ottamatta käärmeitä ja joitain muita lajeja) silmät ovat paremmin kehittyneet kuin sammakkoeläimillä: ne ovat liikkuvampia. Niiden sopeutuminen näkemiseen eri etäisyyksillä (melko suuri monilla lajeilla) ei ole varmistettu vain, kuten kaloilla ja sammakkoeläimillä, linssin liikkeellä, vaan myös, kuten korkeammilla selkärankaisilla, muuttamalla sen kaarevuutta. Silmäluomien lisäksi siinä on nittoiva kalvo. Useilla lajeilla on väliseinään yhteydessä oleva parietaalinen elin, joka pystyy havaitsemaan valoärsykkeitä. Joitakin progressiivisia muutoksia on tapahtunut kuuloelimessä: alapussi on hieman kohonnut; kuuloluun luuranko siirtää tärinää paremmin tärykalvo sisäkorvaan; soikean ikkunan lisäksi ilmestyi toinen ikkuna - pyöreä, mikä lisää labyrintia ympäröivän eksolymfin liikkuvuutta; tärykalvo ei sijaitse pään pinnalla, vaan pienessä syvennyksessä. Hajuelimet ovat kehittyneet melko hyvin, niiden sisäpinta on kasvanut. Useimpien lajien makuelimet ovat huonosti kehittyneet. Kielellä on tärkeä rooli kosketuksessa.
Luuranko. Se on lähes kokonaan luutunut karkean kuituaineen korvaamisen vuoksi luukudosta, sammakkoeläimille ominaista, ohut kuitumainen, sen lujuus on lisääntynyt, tuki- ja suojatoiminnot ovat parantuneet (kuva 96).
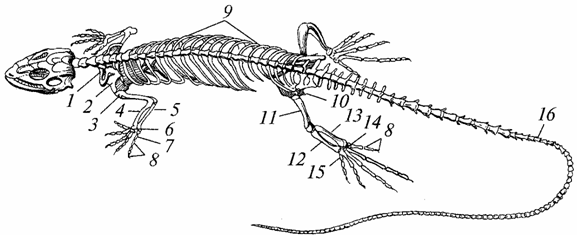
Riisi. 96. Liskon luuranko:
7 - solisluun; 2 - lapaluu; 3 - olkapää; 4 - säde; 5 - kyynärluu; b - ranne; 7 - metacarpus; 8 - sormien falangit; 9 - kylkiluut; 10 - lantio; 11 - reisi; 12 - iso sääriluu; 13 - pohjeluu; 14 - tarsus; 15 - metatarsus; 16 - hännän nikamat
Kallo laajentunut; aivolohkon kapasiteetista tuli suurempi kuin sammakkoeläinten. Sivupinta muinaisten matelijoiden kallo oli kiinteä, siihen ilmestyi myöhemmin ajallisia kuoppia - yksi tai kaksi, mikä vähensi sen massaa ja lisäsi kiinnityspintaa pureskelu lihaksia. Alaleuat, kuten sammakkoeläintenkin, on ripustettu kallon aivoalueelta neliömäisen luun avulla ja koostuvat myös useista luista (hammas-, nivelluista jne.).
Ensimmäisen nikaman runko (atlas) on kasvanut hampaan muodossa toisen nikaman runkoon (epistrofia). Rengasmainen atlas sai mahdollisuuden kiertää epistrofian hampaan ympäri, minkä ansiosta pään käännökset tulivat mahdollisiksi, ja tämä oli hyvin tärkeä ruoan hankkimiseen (pääasiassa muihin eläimiin) ja vihollisilta suojaamiseen. Atlas on yhdistetty kallon takaraivoon yhden kondylen avulla, mikä myös lisää pään liikkuvuutta. Selkäranka on jaettu viiteen osaan: kohdunkaulan, rintakehän, lannerangan, sakraalisen ja kaudaalisen osaan. Selkänikamat rintakehä ovat yhteydessä rintalastaan hyvin kehittyneiden kylkiluiden avulla, minkä seurauksena muodostuu rintakehä, jota sammakkoeläimillä ei ole. Lyhennetyt kylkiluut ulottuvat lannenikamista, eivät ylety rintalastaan. Ristinikamia on kaksi (sammakkoeläimillä yksi). Häntäalueella on monia nikamia (kilpikonnia lukuun ottamatta).
Jalkojen luut ovat paremmin kehittyneitä ja kestävämpiä kuin sammakkoeläinten luut. Ranteen ja olkaluun välillä on liikkuvia niveliä. Siksi kyynärvarsi ja ranteen proksimaalinen rivi sekä sääri ja tarsuksen proksimaalinen rivi toimivat yhtenäisinä muodostelmina. Erityisen huomionarvoista on raajojen vöiden vahvistuminen: olkapää - sen yhteyksien vuoksi rintalastan ja rintarangan kanssa, lantio - koska se nojaa kahteen sakraaliset nikamat ja sen molemmat puolikkaat muodostavat kaksi tartuntaa - ischial ja häpy.
Lihaksisto. Se on erilaistunut, kehittynyt uusia lihasryhmiä, joilla pää kääntyy, matelijoilla syntyneet niskan liikkeet, laajeneminen ja kapeneminen rinnassa Jalkojen lihakset kasvoivat merkittävästi, sormien koukuttajat ja ojentajat kehittyivät. Koko järjestelmän massa ja lujuus ovat kasvaneet.
Ruoansulatuselimistö. Se on erilaistuneempi kuin sammakkoeläimissä (kuva 97). Hampaat keskittyvät pääasiassa leukoihin, niiden lukumäärä on suuri, mutta ne ovat samantyyppisiä ja palvelevat pääasiassa saalista. Sylkirauhaset ovat kehittyneempiä kuin sammakkoeläimillä (useissa lajeissa näiden rauhasten eritteet ovat myrkyllisiä). Matelijoilla alkaa kovan kitalaken muodostumisprosessi, joka on erityisesti kehittynyt krokotiileilla. Kovan kitalaen muodostumisen vuoksi suuontelo on erotettu nenänielun ontelosta, mikä helpottaa sekä hengitystä että ruoan imeytymistä. Nielu, ruokatorvi ja vatsa ovat hyvin kehittyneitä (etenkin selkärankaisia syövillä petoeläimillä). Maksa (sappirakon kanssa) ja haima ovat täydellisempiä kuin sammakkoeläinten. Ohutsuoli huomattavasti pitkänomainen, paksusuoli on lyhyt ja päättyy kloakaan. Nykyaikaiset matelijat ruokkivat pääasiassa eläimiä, kasvinsyöjälajien määrä on merkityksetön.
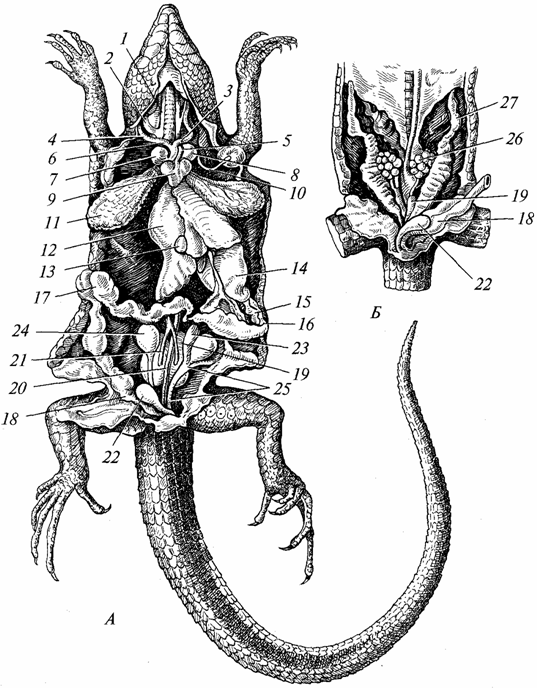
Riisi. 97. Sisäelimet liskoja; Miespuolinen; B - nainen:
/ - ulkoinen ja 2 - sisäinen kaulalaskimot; 3 - vasen ja 4 - oikea kaulavaltimo; 5 - vasen ja 6 - oikea aortan kaari; 7 - oikea ja 8 - vasen atrium; 9 - sydämen kammio; 10 - vasen subclavian valtimo 11 - keuhko; 12 - maksa; 13 - sappirakko; 14 - vatsa; /5 - haima; 16 - pohjukaissuoli; 17 - paksusuoli; 18 - peräsuoli; 19 - munuainen; 20 - dorsaalinen aortta; 21 - munuaisten efferentit suonet; 22 - virtsarakko; 23 - kiveksen lisäosa; 24 - kives; 25 - siemenputki; 26 - munasarja; 27 - munanjohdin
Hengityselimet. Matelijoiden hengityselimet ovat keuhkot (ks. kuva 97), koska niiden iho ei sovellu kaasunvaihtoon; limakalvo suuontelon, jolla on merkittävä rooli sammakkoeläinten hengityksessä, käyttää tätä tehtävää muutamassa matelijalajissa. Tästä syystä ja erityisesti matelijoiden sammakkoeläimiin verrattuna lisääntyneen aktiivisuuden yhteydessä keuhkojen rakenne ja hengitysteitä monimutkainen. Keuhkot ovat suurempia elimiä ja sijaitsevat syvemmällä rintaontelossa kuin sammakkoeläimillä. Niiden sisäpinta on kasvanut merkittävästi, koska siihen on kehitetty monia tiheämmällä verkostolla varustettuja poikkipalkkeja. verisuonet. Ilma pääsee keuhkoihin kulkemalla choanaen, nielun, kurkunpään, pitkän henkitorven läpi, jonka onteloa tukevat rustorenkaat. Henkitorven läpi kulkemisen aikana ilma puhdistuu pölystä, sen lämpötila lähestyy kehon lämpötilaa. Sammakkoeläimillä henkitorvi oli lapsenkengissään. Henkitorven takapää on jaettu kahteen keuhkoputkeen, joiden seinissä on myös rustorenkaita. Sisään- ja uloshengityksen toteuttaminen on tehokkaampaa kuin sammakkoeläimissä, koska matelijoilla on kehittynyt rintakehä, jonka tilavuus kasvaa sisäänhengityksen myötä ja pienenee uloshengityksen myötä. Sama mekanismi on säilynyt korkeammissa selkärankaisissa - linnuissa ja nisäkkäissä.
Verenkiertoelimistö. Sydän, kuten sammakkoeläintenkin, on kolmikammioinen, mutta kammioon ilmestyy alhaalta väliseinän alkio. Krokotiileilla se on jo täynnä, ja heidän sydämensä muuttuu nelikammioiseksi (kaksi eteistä ja kaksi kammiota). Tämän seurauksena matelijoilla laskimo- ja valtimoverivirtojen erottuminen tehostuu, mutta molempien virtojen täydellinen eristäminen ei ole vielä täydellinen edes krokotiileilla, kuten seuraavasta kuvauksesta (kuva 98) voidaan nähdä.
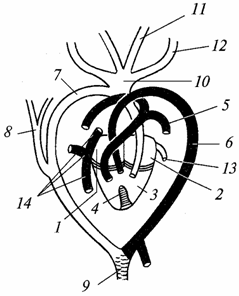
Riisi. 98. Liskon sydän ja pääsuonet (vaaleat suonet valtimoverellä, varjostus - sekaverellä; musta - laskimoveren kanssa):
1 - Oikea eteinen; 2 - vasen atrium; 3 - kammio; 4 - epätäydellinen väliseinä, joka jakaa kammion vasempaan ja oikeaan puoliskoon; 5 - keuhkovaltimo; 6 - vasen aortan kaari; 7 - oikea aortan kaari; 8 - subclavian valtimo; 9 - dorsaalinen aortta; 10 - yhteinen runko kaulavaltimot; 11 - vasen sisäpuoli kaulavaltimo, 12 - vasen ulkoinen kaulavaltimo; 13 - keuhkolaskimo; 14 - takaisin alaonttolaskimon ja anterioriset onttolaskimot (kaulalaskimot).
Valtimoveri tulee vasempaan eteiseen - se tulee keuhkoista. Laskimoveri tulee oikeaan eteiseen - se kerätään kaikista kehon osista. Lisäksi veri molemmista eteisistä tulee kammioon: sen oikeaan puoliskoon - laskimo, vasemmassa - valtimo. Väliseinän, vaikkakin epätäydellisen, vuoksi kammiossa oleva veri sekoittuu vähemmän kuin sammakkoeläimissä. Valtimokartiota ei ole, ja päävaltimorungot lähtevät suoraan sydämestä.
From oikea puoli kammiot lähtevät keuhkovaltimot(entinen neljäs haaravaltimopari), joka kuljettaa laskimoverta keuhkoihin. Entinen kolmas haarasuonipari surkastui. Toinen suonipari koostuu kahdesta aortan kaaresta: oikea kaari alkaa kammion vasemmalta puolelta ja kuljettaa valtimoverta, vasen kaari alkaa kammion oikealta puolelta (mutta lähempänä sen vasenta puolta) ja kuljettaa sekaverta . Aortan oikeasta kaaresta kaulavaltimot (entinen ensimmäinen kidussuonipari) ulottuvat päähän. Ne toimittavat valtimoverta aivoille, jotka ovat erityisen herkkiä hapenpuutteelle. Molemmat aorttakaaret yhdistyvät muodostaen aortan, joka haarautuu useiksi valtimoiksi. Siten matelijan kehossa on pääasiassa sekoittunutta verta, mutta se on kyllästetty enemmän happea kuin sammakkoeläimet, mikä edistää intensiivisempää aineenvaihduntaa.
eritysjärjestelmä. Se on muuttunut paljon matelijoiden lisääntyneen aineenvaihdunnan vuoksi, mikä johti dissimilaatiotuotteiden määrän kasvuun ja niiden täydelliseen sopeutumiseen maanpäälliseen elämäntapaan, minkä seurauksena tuli tarpeelliseksi käyttää sisään tulevaa vettä taloudellisemmin. Vartalo. Siksi matelijat kehittivät monimutkaisempia erityselimiä, joita kutsutaan lantion tai toissijaisiksi munuaisiksi (metayaephros). Kaikissa näiden munuaisten tubuluksissa ei ole suppiloa ja ne alkavat Bowmanin kapseleilla, jotka ympäröivät Malpighian ruumiita lisääntyneillä kapillaareilla. Lantion munuaisten tubulukset ovat erittäin pitkiä ja tiiviisti kietoutuneet kapillaareihin. Bowmanin kapseleissa dissimilaatiotuotteista liuotetaan veteen ja joihinkin eloperäinen aine(esim. glukoosi) muodostuu primäärinen virtsa. Jälkimmäinen, kulkiessaan pitkien munuaistiehyiden läpi, antaa verelle takaisin seiniä punovien kapillaarien kautta merkittävän osan vedestä ja muista elimistölle välttämättömistä aineista. Tämän jälkeen muodostunut sekundaarinen virtsa ei poistu susikanavien kautta, vaan vasta kehittyneiden virtsajohtimien kautta kloakaan ja kerääntyy virtsarakon. Primaarisen ja sekundaarisen virtsan muodostumisprosessit suoritetaan myös rungon munuaisissa, mutta pienemmän määrän vettä palaamalla vereen. Primaariset vedessä elävät selkärankaiset, erityisesti sammakkoeläimet, joiden iho on paljas, kärsivät usein veden ylimäärästä, eivät sen puutteesta. On myös huomattava, että matelijoilla proteiinien hajoamisen lopputuote on Virtsahappo, jonka poistamiseen kehosta virtsan mukana kuluu vähän vettä. Sammakkoeläimillä ja useilla muilla primäärisillä vesiselkärankaisilla proteiinien hajoaminen johtaa urean muodostumiseen, jonka poistaminen vaatii paljon vettä.
Matelijoiden alkioissa on päämunuaisen alkuosa ja kehittynyt runkomunuainen, joka toimii aikuisilla (usilla liskoilla) murrosikään asti. Nämä tosiasiat vahvistavat kolmen tyypin munuaisten jatkuvuuden selkärankaisten evoluution aikana.
Jäljentäminen. Matelijoiden urosten lisääntymisjärjestelmä sisältää kaksi pyöreän muotoista kivestä, jotka makaavat lannerangan sivuilla, kaksi kivesten lisäosaa, jotka muodostuvat rungon munuaisten etuosista, kaksi siemenkanavaa (Wolffin kanavat) ja seinästä kehittyneet paritteluelimet kloaakasta. Naisten lisääntymisjärjestelmä koostuu kahdesta munasarjasta, jotka kasvavat voimakkaasti pesimäkauden aikana, ja kahdesta munasarjasta (Müllerian kanavat), jotka virtaavat kloakaan. Lannoitus on vain sisäistä. Urokset tuovat siemennestettä naaraan kloakaan parituselinten avulla. Kypsät munat, jotka sisältävät paljon keltuaista, putoavat munanjohtimien suppiloihin. Naaras- ja miessukusolujen yhteys tapahtuu munanjohtimien yläosassa, jonne siittiöt tunkeutuvat yhdynnän jälkeen. Tsygootit, jotka liikkuvat munanjohtimia pitkin, jälkimmäisten keskiosassa on proteiinikuoren ympäröimiä, ja takana ne peitetään kuorella.
Joissakin lisko- ja käärmelajeissa havaitaan naaraiden partenogeneettistä lisääntymistä. Näissä lajeissa ei yleensä ole uroksia.
Kehitys. Matelijoilla kehitys on suoraa; matelijoilla ei ole toukkia. Matelijat munivat yleensä hedelmöittyneitä munia maaperään ja jotkut lajit mätänevien kasvijätteiden alle. Monissa lajeissa tsygoottien kehittyminen tapahtuu naaraan sukuelinten alueella, ja heti kehittyneiden munien vapautumisen jälkeen niistä kuoriutuu ulos muodostuneita eläimiä, eli sellaisille lajeille on ominaista ovovivipariteetti. Muutamissa lajeissa havaitaan todellista elävänä syntymää: keltuaispussin verisuonet ovat tiiviisti yhteydessä munanjohtimen suoniin ja alkio saa merkittävän osan ravinteitaäidin ruumiista. Oviparous havaitaan useammin lajeissa, jotka ovat yleisiä pohjoisilla ja vuoristoisilla alueilla. Ilmeisesti ovovivipariteetti on kehittyvien alkioiden suojaaminen jäähtymiseltä ja muilta epäsuotuisilta olosuhteilta. Ei ole olemassa yhtä matelijalajia, jonka munat kehittyvät vedessä. Jopa matelijoissa, jotka johtavat suurin osa elämä vedessä (kaikki krokotiilit, monet kilpikonnat, jotkut käärmeet), munasolut ja kehitys tapahtuvat vain maalla. Merikäärmeet, jotka eivät koskaan poistu vedestä, ovat eläviä.
Matelijoiden munilla ja alkioilla on useita mukautuksia maalla kehittymiseen. Ensin munat peitetään kuorilla ja muilla kalvoilla, jotka suojaavat niitä kuivumiselta ja vaurioilta. Toiseksi alkiot kehittävät erityisiä alkiokalvoja (seroosi, amnion ja allantois), jotka ovat erittäin tärkeitä alkion kehitys eläimet kuivalla maalla. Serosa ja amnion kehittyvät samasta alkion ruumiin kaksoispoimusta, jotka ovat peräisin ektodermista ja mesodermista. Tämä taite näkyy kahdesta vastakkaiset puolet alkukeinojen muodossa, jotka kasvavat toisiaan kohti ja sulkeutuvat alkion yli. Taitteen ulompi osa muuttuu seroosiksi (sen ulompi kerros on ektodermaalinen, sisempi mesodermaalinen) ja sisäosa muuttuu amnioniksi (sen ulompi kerros on mesodermaalinen, sisempi on ektodermaalinen). Amnionin muodostama ontelo on täytetty vesipitoisella nesteellä, jonka vuoksi kehitys tapahtuu kosteassa ympäristössä, ja alkio on suojattu kuivumiselta ja erilaisten ravistelujen vaikutuksilta, jotka ovat erityisen vaarallisia ilmassa.
Alkuperä. Ensimmäiset hiilen toisella puoliskolla syntyneet matelijat elivät kosteissa maabiotoopeissa ja luultavasti lisääntyivät ja kehittyivät vedessä. Tähän mennessä maa oli jo asuttama alemmilla maan kasveilla, erilaisilla selkärangattomilla, hyönteisten valloitus alkoi. Matelijoilla, jotka pystyivät kulkemaan sammakkoeläimiä kauemmaksi vesistöistä, oli runsaasti ravintoa, eivätkä he kohdanneet vakavia vihollisia. Hiilen lopussa ja toistuvasti permin aikana ilmasto muuttui kuivemmaksi, mikä aiheutti monien sammakkoeläinten kuoleman ja päinvastoin edesauttoi matelijoiden asteittaista kehitystä ja laajempaa leviämistä maalla. Jonkin ajan kuluttua ne siirtyivät lisääntymiseen ja kehitykseen vesistöjen ulkopuolella, ensin kosteammissa maabiotoopeissa ja sitten kuivemmissa paikoissa. Mesozoisen aikakauden aikana ne levisivät ympäri maailmaa ja monia erilaisia ryhmiä. Petoeläinten ohella "eläsi monia kasvinsyöjiä, lentoon sopeutuneita matelijoita ja merissä eläviä matelijoita. Niiden ruumiinkoko oli erilainen, mutta useimmat olivat suuria eläimiä, ja jotkut jättimäisiä - jopa 20 metriä pitkiä. Ilmasto lähes koko mesozoic aikakauden oli lämmin, erittäin suotuisa näille eläimille, joilla ei ole tarpeeksi korkea vakio lämpötila ja menettävät aktiivisuutensa, kun ympäristö jäähtyy.
Mesozoisen aikakauden lopussa maapallo tapahtui voimakkaita vuorenrakennusprosesseja, joihin liittyi voimakas muutos olemassaolon olosuhteissa (lämpötila, kosteus jne.). Tämä johti mesotsooisten matelijoiden massasukupuuttoon, jotka eivät kestäneet olemassaolotaistelua heille niin vaikeissa abioottisissa ympäristöolosuhteissa, joita vaikeutti kilpailu nisäkkäiden ja lintujen kanssa. Samaan aikaan syntyi uusia matelijaryhmiä, jotka lisääntyivät Cenozoic aikakaudella - liskoja ja käärmeitä polveutuivat niistä. Vanhoista ryhmistä on säilynyt vain yksi hyvin primitiivinen primitiivilaji, pieni määrä krokotiileja ja kilpikonnia. Nykyaikaiset matelijat ovat levinneet pääasiassa lämpimiin maihin, erityisesti subtrooppisiin ja trooppisiin maihin, niitä on vähän lauhkean ilmaston maissa, ja niitä ei esiinny kaukana pohjoisessa.

