רֶפלֶקס- תגובה של הגוף לגירוי של קולטנים, המתבצע על ידי מערכת העצבים.
קשת רפלקס - הנתיב שבו עובר הדחף העצבי במהלך יישום הרפלקס. מורכב מ-5 חלקים:
- קולטן - מבנה רגיש המסוגל להגיב לסוג מסוים של גירוי;
- נוירון רגיש מוליך דחפים למוח;
- הנוירון הבין-קלורי מחבר נוירונים רגישים ומבצעים, ממוקם בחוט השדרה או במוח;
- נוירון מבצעי (מוטורי) מוליך דחפים מהמוח אל האיברים המבצעים;
- סוכנות מבצעת- שריר (חוזים), בלוטה (סודות) וכו'.
רפלקסים לא מותנים
- נוכח מלידה
- במהלך החיים הם לא משתנים ולא נעלמים;
- זהים בכל האורגניזמים מאותו מין;
- להתאים את הגוף לתנאים קבועים.
רפלקסים מותנים
- נרכש במהלך החיים;
- עשוי להשתנות או להיעלם;
- לכל אורגניזם יש משלו;
- להתאים את הגוף לתנאים משתנים.
מבחנים
1) איזה חוליה משמשת כתחילתה של קשת הרפלקס
א) נוירון בין קלוריות
ב) נוירון רגיש
ב) קולטן
ד) נוירון מנהלים
2. באיזה רצף נכללים מרכיבי קשת הרפלקס ביישום הרפלקס
א) איבר מנהל, נוירון מוטורי, נוירון בין קלוריות, נוירון חושי, קולטן
ב) נוירון בין קלוריות, נוירון חושי, נוירון מוטורי, קולטן, איבר מנהל
ג) קולטן, נוירון חושי, נוירון בין קלוריות, נוירון מוטורי, איבר מנהל
ד) נוירון חושי, נוירון בין קלוריות, קולטן, איבר מנהל, נוירון מוטורי
3. השלב הראשוני של קשת הרפלקס ברפלקס הרוק
א) בלוטת רוק
ב) קולטן
ב) נוירון בין קלוריות
ד) נוירון מנהלים
4. פעולת הגירויים גורמת להתרחשות דחף עצבי V
א) נוירונים תחושתיים
ב) נוירונים מוטוריים
ב) קולטנים
ד) נוירונים בין קלוריות
5. עירור מכוון לאורך הנוירון הרגיש
א) למערכת העצבים המרכזית
ב) לגוף המבצע
ב) לקולטנים
ד) לשרירים
6. רצפטורים הם תצורות רגישות ש
א) להעביר דחפים למערכת העצבים המרכזית
ב) להעביר דחפים עצביים מהנוירונים הבין-קלוריים למנהל
ג) לתפוס גירויים ולהמיר את האנרגיה של גירויים לתהליך של עירור עצבי
ד) לקבל דחפים עצביים מנוירונים תחושתיים
7. דחפים עצביים מועברים למוח דרך נוירונים
א) מנוע
ב) intercalary
ב) רגיש
ד) מבצעת
8. קצוות קשת רפלקס
א) מנהל
ב) נוירון רגיש
ב) קולטן
ד) נוירון בין קלוריות
9. דחפים עצביים מקולטנים להתנהלות מערכת העצבים המרכזית
א) נוירונים תחושתיים
ב) נוירונים מוטוריים
ג) נוירונים תחושתיים ומוטוריים
ד) נוירונים בין קלוריות ומוטוריים
10. תצורות עצביםבגוף האדם, קולטים גירויים מהסביבה החיצונית, הם
א) עצבים
ב) קולטנים
ב) המוח
ד) צמתים עצביים
11. הפיכת הגירויים לדחף עצבי מתרחשת ב
א) עצבים מוטוריים
ב) המוח
ב) קולטנים
ד) חוט השדרה
12. דחפים עצביים מועברים מהחושים האנושיים דרך נוירונים
א) רגיש
ב) מנוע
ב) להכניס
ד) מבצעת
13. לתפוס מידע ולהמיר אותו לדחפים עצביים
א) קולטנים
ב) צמתים עצביים
ב) גופים מבצעים
ד) נוירונים בין קלוריות
14. רפלקס מותנה
א) עובר בירושה לצאצאים, אך אינו נמשך לאורך כל החיים
ב) נרכש ואבד בקלות על ידי הגוף במהלך החיים
ג) עובר בירושה לצאצאים מהורים ונמשך עבור
ד) היא תגובה מתמדת של הגוף לגירויים סביבתיים מוגדרים בקפדנות
15. דוגמה לרפלקס מותנה בחתול
א) ריור בזמן האכילה
ב) נסיגה של הכפה בזמן כוויה
ב) תגובה לכינוי
ד) תגובה לצליל חד
16. ריור בבני אדם למראה לימון - רפלקס
א) מותנה
ב) ללא תנאי
ב) מגן
ד) מעיד
17) רפלקסים מותנים בבני אדם נוצרים בתהליך
א) התפתחות אישית
ב) היווצרות מינים
ג) התפתחות היסטורית
ד) היווצרות עוברים
18. דוגמה לרפלקס מותנה היא
א) ריור למראה לימון חתוך
ב) ריור במהלך האכלה
ג) התכווצות האישון בחשיפה לאור חזק
ד) נסיגה של היד עם זריקה פתאומית
19. התגובה של ילד לבקבוק חלב היא רפלקס ש
א) עובר בירושה
ב) נוצר ללא השתתפות הקורטקס ההמיספרות
ב) נרכשים במהלך החיים
ד) נמשכת לאורך כל החיים
20. רפלקס עיטוש מגן
א) לא עובר בירושה
ב) הוא מותנה
ב) נחלש עם הזמן
ד) מאפיין את כל הפרטים של המין
21. תגובות הנרכשות על ידי בני אדם ובעלי חיים במהלך החיים ומתן הסתגלות לתנאי סביבה משתנים נקראות
א) רפלקסים מותנים
ב) אינסטינקטים
ב) רפלקסים בלתי מותנים
ד) רפלקסים שעוברים בתורשה
22. רפלקס לא מותנה
א) עובר בירושה
ב) נרכש במהלך החיים
ג) מופק עבור אותות מסוימים
ד) עומד בבסיס אותות חיצוניים שונים
23. רפלקסים בלתי מותנים של בני אדם ובעלי חיים מספקים
ג) שליטה במיומנויות מוטוריות חדשות על ידי הגוף
24. תגובה אנושית ל צבע ירוקרמזור הוא רפלקס
א) מולד
ב) נרכש
ב) ללא תנאי
ד) בירושה
25. הדגש מיץ קיבהבתגובה לקולטנים לגירוי מזון חלל פה
א) התרגשות
ב) בלימה
ב) רפלקס בלתי מותנה
ד) ויסות עצמי
26. המשמעות הביולוגית של רפלקסים מותנים עבור הגוף היא שהם
א) לשמור על הקביעות של הסביבה הפנימית
ב) ספציפי למין ונמשך לאורך כל החיים
ב) לקדם אינטראקציה מערכות פונקציונליות
ד) לספק התאמה לתנאי הסביבה המשתנים
27. בתהליך החיים, בעלי חיים רוכשים רפלקסים
א) ללא תנאי
ב) בירושה
ג) מאפיין את כל הפרטים של מין נתון
ד) לאפשר להם לשרוד בתנאים משתנים
28. רפלקס מותנה בבני אדם הוא
א) כיווץ אישונים באור בהיר
ב) הפניית הראש לצליל חד
ג) הפרשת רוק כאשר אוכל נכנס לפה
ד) תגובה למשמעות המילה
29. דוגמה לרפלקס מותנה היא
1) היכולת לרכוב על אופניים
2) ריור בעת אכילה
3) רצון לישון כשעייפים
4) התעטשות ושיעול
30. רפלקסים מותנים של בני אדם ובעלי חיים מספקים
א) התאמה של האורגניזם לתנאי סביבה קבועים
ב) הסתגלות הגוף לעולם חיצוני משתנה
ג) התפתחות של אורגניזמים בעלי מיומנויות מוטוריות חדשות
ד) הבחנה בין חיות של פקודות המאלף
31. רפלקסים מולדים
א) הם מאפייני מין של אורגניזם
ב) מאפיין פרטים בודדים מהמין
ג) דורשים תנאים נוספים ליישום
ד) אינם עוברים בירושה
צורה ראשית פעילות עצבניתהוא רפלקס. רפלקס - תגובה סיבתית של הגוף לשינויים בסביבה החיצונית או הפנימית, המתבצעת בהשתתפות חובה של המרכז מערכת עצביםבתגובה לגירוי קולטן. עקב רפלקסים, מתרחשת התרחשות, שינוי או הפסקה של כל פעילות של הגוף.
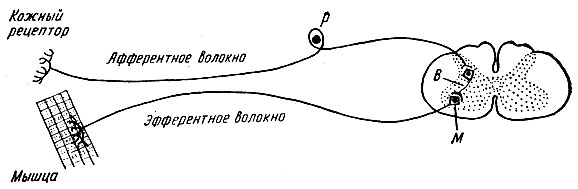
מסלול העצבים שלאורכו מתפשט העירור במהלך יישום הרפלקסים נקרא קשת רפלקס.
קשתות רפלקס מורכבות מחמישה מרכיבים: 1) קולטן; 2) מסלול עצבי אפרנטי; 3) מרכז רפלקס; 4) מסלול עצבי efferent; 5) אפקטור (גוף עובד).
קוֹלֵט- זהו קצה עצב רגיש שתופס גירוי. בקולטנים, האנרגיה של הגירוי מומרת לאנרגיה של דחף עצבי. הבחנה: 1) אקסטרורצפטורים- נרגש בהשפעת גירויים מהסביבה (קולטנים של העור, העיניים, אוזן פנימית, רירית האף והפה); 2) אינטררצפטורים- לתפוס גירויים מהסביבה הפנימית של הגוף (קולטנים איברים פנימיים, כלים); 3) פרופריורצפטורים- להגיב לשינוי במיקום של חלקים בודדים של הגוף במרחב (קולטנים של שרירים, גידים, רצועות, שקיות מפרקים).
מסלול עצבי אפרנטימיוצג על ידי תהליכים של נוירונים קולטן הנושאים עירורים למערכת העצבים המרכזית.
מרכז רפלקסמורכב מקבוצה של נוירונים הממוקמים ברמות שונות של מערכת העצבים המרכזית ומעבירים דחפים עצביים מהנתיב העצבי האפרנטי ל-efferent.
מסלול עצבי efferentמוליך דחפים עצביים ממערכת העצבים המרכזית אל האפקטור.
מַפעִיל- איבר ביצוע, שפעילותו משתנה בהשפעת דחפים עצביים המגיעים אליו דרך תצורות קשת הרפלקס. המשפיעים יכולים להיות שרירים או בלוטות.
קשתות רפלקסיכול להיות פשוט או מורכב. קשת רפלקס פשוטה מורכבת משני נוירונים - תופס ואפקטור, שביניהם יש סינפסה אחת. תרשים של קשת רפלקס דו-נוירונים כזו מוצג באיור. 71.
דוגמה לקשת רפלקס פשוטה היא קשתות רפלקס הגיד, כגון קשת רפלקס הפיקה.
קשתות הרפלקס של רוב הרפלקסים כוללות לא שניים, אלא מספר גדול יותר של נוירונים: קולטן, אחד או יותר intercalary ו-effector. קשתות רפלקס כאלה נקראות מורכבות, רב-עצביות. תרשים של קשת רפלקס מורכבת (שלושה נוירונים) מוצגת באיור. 72.
כעת הוכח שבמהלך התגובה של האפקטור, קצוות עצבים רבים הנמצאים באיבר הפועל נרגשים. דחפים עצביים כעת מהאפקטור נכנסים שוב למערכת העצבים המרכזית ומודיעים לה על התגובה הנכונה של האיבר העובד. לפיכך, קשתות הרפלקס אינן פתוחות, אלא תצורות טבעת.
הרפלקסים מגוונים מאוד. ניתן לסווג אותם לפי מספר קריטריונים: 1) משמעות ביולוגית(אוכל, הגנתי, מיני); 2) בהתאם לסוג הקולטנים המגורים: אקסטרוספטיביים, אינטרוצפטיים ופרופריוצפטיבים; 3) לפי אופי התגובה: מוטורי או מוטורי (איבר ביצוע - שריר), הפרשה (אפקטור - ברזל), vasomotor (התכווצות או התרחבות של כלי דם).
ניתן לחלק את כל הרפלקסים של האורגניזם כולו לשניים קבוצות גדולות: ללא תנאי ומותנה. ההבדלים ביניהם יטופלו בפרק י"ב.
לפעילות מערכת העצבים יש אופי רפלקס. אפילו במאה ה- XVII. הפילוסוף והמתמטיקאי הצרפתי רנה דקארט תיאר את פעולת הרפלקס. הוא ציין את תגובת הגוף לגירוי והציע את קיומו של נתיב שלאורכו עוברת עירור עצבי. המונח "רפלקס" עצמו הועלה מאוחר יותר - במאה ה-18 - על ידי המדען הצ'כי ג'יי פרוצ'אזקה (מהלטינית "רפלקס" - פעולה משתקפת). מאוחר יותר, I.M. Sechenov בעבודתו "רפלקסים של המוח" הוכיח כי התגובות של מערכת העצבים ל סוגים שוניםגירויים ממשיכים לפי מנגנון הרפלקס, כלומר. כל הפעולות המודעות והלא מודעות הן ממקור רפלקס. רפלקס מובן כתגובה ספציפית של הגוף לגירוי של הסביבה הפנימית או המתקבל מבחוץ בהשתתפות חובה של מערכת העצבים המרכזית. רפלקסים נקראים בדרך כלל יחידות פונקציונליות של פעילות עצבית.
מנגנונים של רפלקסים של קשת הרפלקס
ניתן לסווג רפלקסים לפי קריטריונים שונים. אז, בהתאם לרמת סגירת הקשת, כלומר. על פי מיקום מרכז הרפלקס, הרפלקסים מחולקים לשדרה (הרפלקס נסגר בחוט השדרה), בולבר (מרכז רפלקס - לָשָׁד), mesencephalic (מבנה קשת הרפלקס סגור במוח האמצעי), מרכזי רפלקס דיאנצפליים וקורטיקליים נמצאים ב טלנספאלוןוקליפת המוח, בהתאמה. על פי תכונת האפקטור, הם סומטיים, כאשר הנתיב האפרנטי של הרפלקס מספק עיוור מוטורי של שרירי השלד, וצמחיים, כאשר האיברים הפנימיים הם המשפיעים. בהתאם לסוג הקולטנים המגורים, הרפלקסים מחולקים ל-exteroceptive (אם הקולטן קולט מידע מהסביבה החיצונית), פרופריוספטיבי (קשת הרפלקס מתחילה מהקולטנים של מנגנון השריר והשלד) ואינטרוצפטיב (מקולטנים של איברים פנימיים). רפלקסים אינטרוספטיביים, בתורם, מחולקים לקרביים-קרביים (קשת רפלקס מחברת שני איברים פנימיים), קרביים-שריריים (קולטנים ממוקמים על מנגנון השריר-גיד, האפקטור הוא איבר פנימי) וקרביים-עוריים (קולטנים הם מקומי בעור, איברים עובדים - קרניים). לפי פבלוב, הרפלקסים מחולקים למותנים (מפותחים במהלך החיים, ספציפיים לכל פרט) ולא מותנים (מולדים, ספציפיים למין: מזון, מיני, הגנתי-מוטורי, הומאוסטטי וכו').
כיצד פועלת קשת הרפלקס?
ללא קשר לסוג הרפלקס, קשת הרפלקס שלו מכילה קולטן, מסלול אפרנטי, מרכז עצבים, מסלול efferent, איבר עובד ומשוב. היוצא מן הכלל הוא רפלקסי האקסון, שמבנה קשת הרפלקס שלו ממוקם בתוך נוירון אחד: תהליכים תחושתיים יוצרים דחפים צנטריפטליים, העוברים דרך גוף הנוירון, מתפשטים לאורך האקסון למערכת העצבים המרכזית ולאורך הענף. של האקסון, הדחפים מגיעים לאפקטור. רפלקסים כאלה מיוחסים לתפקוד מערכת העצבים המטא-סימפתטית; באמצעותם, למשל, מתבצעים מנגנונים לוויסות טונוס כלי הדם ופעילות בלוטות העור.
הפונקציה של תפיסת גירוי והמרתו לאנרגיית עירור מתבצעת על ידי קולטנים של קשתות רפלקס. לאנרגיית הקולטן של עירור יש אופי של תגובה מקומית, שחשובה בהדרגה של עירור לפי כוח.
בהתבסס על המבנה והמקור של הקולטנים, ניתן לחלק אותם לקצות עצבים תחושתיים ראשוניים, תחושתיים משניים וקצות עצבים חופשיים. בראשון, הנוירון עצמו פועל כקולטן (הוא מתפתח מהנוירופיתל); אין מבנים מתווכים בין הגירוי לנוירון האפרנטי הראשון. התגובה המקומית של הקולטנים התחושתיים הראשוניים - פוטנציאל הקולטן - היא גם פוטנציאל מחולל, כלומר. גרימת פוטנציאל פעולה על פני הממברנה של הסיב האפרנטי. קולטנים תחושתיים ראשוניים כוללים קולטנים חזותיים, חוש הריח, כימו- ובארורצפטורים. של מערכת הלב וכלי הדם.
תאי חישה משניים הם מבנים מיוחדים ממקור לא עצבי המקיימים אינטראקציה עם הדנדריטים של תאי חישה פסאודו-חד-קוטביים בעזרת מגעים של קולטן עצבי סינפטי. פוטנציאל הקולטן הנובע מפעולת גירוי בתאי החישה המשני אינו מחולל ואינו גורם להופעת פוטנציאל פעולה על גבי הממברנה של הסיב האפרנטי. הפוטנציאל הפוסט-סינפטי המעורר נוצר רק באמצעות מנגנון השחרור של המתווך על ידי תא הקולט. הדרגת עוצמת הגירוי מתבצעת באמצעות הפרשה של כמויות שונות של המתווך (ככל שהמתווך משתחרר, כך הגירוי חזק יותר).
תאי חישה משניים כוללים קולטנים שמיעתיים, וסטיבולריים, קרוטידים, מישוש ואחרים. לפעמים, בגלל המוזרויות של התפקוד, קבוצה זו כוללת קולטני פוטו, שמבחינה אנטומית ובשל מוצאם מהנוירו-אפיתל, הם בעלי חישה משנית.
קצות עצבים חופשיים הם דנדריטים מסועפים של תאי חישה פסאודו-חד-קוטביים וממוקמים כמעט בכל הרקמות של גוף האדם.
בהתאם לאופי האנרגטי של הגירוי שאליו מגיב הקולטן, הם מחולקים למכנורצפטורים (מגע, ברוררצפטורים, קולטני נפח, שמיעתי, וסטיבולרי; הם, ככלל, קולטים גירוי מכני בעזרת יציאות תאים), כימורצפטורים ( חוש הריח), קולטנים כימיים של כלי הדם, מערכת העצבים המרכזית, קולטני הפוטו (קולטים גירוי דרך יציאות בצורת מוט וחרוט של התא), תרמורצפטורים (מגיבים לשינוי "חם-קר" - גופי Rufini וצלוחיות קראוזה של ממברנות ריריות) ונוציפטורים (קצוות כאב לא מובלעים).
תצורות פוסט קולטן של קשת הרפלקס
היווצרות הפוסט-קולטן של מבנה קשתות הרפלקס היא מסלול אפרנטי שנוצר על ידי נוירון חושי פסאודו-חד קוטבי, שגופו נמצא ב גנגליון עמוד השדרה, ונוצרים אקסונים שורשים גב עמוד שדרה. תפקידו של המסלול האפרנטי הוא להעביר מידע אל הקישור המרכזי, יתרה מכך, בשלב זה המידע מקודד. למטרות אלו, בגוף של בעלי חוליות, נעשה שימוש בקוד בינארי המורכב מתפרצים (מטחים) של דחפים ומרווחים ביניהם. ישנם שני סוגים עיקריים של קידוד: תדר ומרחב.
הראשון הוא היווצרות של מספר שונה של דחפים בפרץ, מספר שונה של פרצים, משך הזמן שלהם ומשך ההפסקות ביניהם, בהתאם לעוצמת הגירוי המופעל על הקולטן. קידוד מרחבי מבצע את הדרגת הכוח של הגירוי, הכולל מספר שונה של סיבי עצב, שלאורכם מתבצעת עירור בו זמנית.
המסלול האפרנטי מורכב בעיקר מסיבים A-α, A-β ו-A-δ.
לאחר שעבר דרך הסיבים, הדחף העצבי נכנס למרכז הרפלקס, שבמובן האנטומי הוא אוסף של נוירונים הממוקמים ברמה מסוימת של מערכת העצבים המרכזית ולוקחים חלק ביצירת רפלקס זה. תפקידו של מרכז הרפלקס הוא לנתח ולסנתז מידע, כמו גם להעביר מידע מהנתיב האפרנטי לנתיב האפרנטי.
פונקציות קשת רפלקס
בהתאם למחלקה של מערכת העצבים (סומטית ואוטונומית), רפלקסים, שמרכזם ממוקם בחוט השדרה, נבדלים בלוקליזציה של נוירונים בין-קליריים. אז, עבור מערכת העצבים הסומטית, מרכז הרפלקס ממוקם באזור הביניים בין הקרניים הקדמיות והאחוריות של חוט השדרה. מרכז הרפלקס של מערכת העצבים האוטונומית (גוף הנוירונים הבין-קלוריים) נמצא בתוכו קרניים אחוריות. החלקים הסומטיים והאוטונומיים של מערכת העצבים שונים גם בלוקליזציה של נוירונים efferent. גופם של נוירונים מוטוריים של מערכת העצבים הסומטית נמצאים בקרניים הקדמיות של חוט השדרה, גופם של נוירונים פרגנגליוניים מערכת אוטונומית- בגובה הקרניים האמצעיות.
האקסונים של שני סוגי התאים יוצרים את הנתיב האפרנטי של קשת הרפלקס. במערכת העצבים הסומטית, הוא רציף, הוא מורכב מסיבים מסוג A-α. היוצאים מן הכלל היחידים הם סיבי A-γ, המוליכים עירור מתאי חוט השדרה לסיבים תוך-פיוסאליים של צירי השריר. המסלול הפושט של מערכת העצבים האוטונומית מופרע פנימה גנגליון אוטונומיממוקם או תוך-קירי (זוג חלק סימפטי), או ליד חוט השדרה (לבד או בפנים תא מטען סימפטי- חלק סימפטי). הסיב הפרה-גנגליוני שייך לסיבי B, הסיב הפוסט-גנגליוני לקבוצת C.
האיבר הפועל לחלק הסומטי של מערכת העצבים הוא שריר השלד המפוספס, בקשת הווגטטיבית האפקטור הוא בלוטה או שריר (לב חלק או מפוספס). בֵּין דרך שונהוהאיבר העובד הוא סינפסה מיונאורלית או נוירו-הפרשה כימית.
קשת הרפלקס נסגרת לטבעת עקב השפעה הפוכה - זרימת הדחפים מקולטני האפקטור חזרה למרכז הרפלקס. תפקוד משוב – איתות למערכת העצבים המרכזית על הפעולה שבוצעה. אם זה לא מבוצע מספיק, מרכז העצבים נרגש - הרפלקס ממשיך. כמו כן, עקב ההשפעה ההפוכה, מתבצעת השליטה בפעילות ההיקפית של מערכת העצבים המרכזית.
הבחנה בין משוב שלילי לחיובי. הראשון, בעת ביצוע פונקציה מסוימת, משיק מנגנון המעכב פונקציה זו. משוב חיובי מורכב מגירוי נוסף של פונקציה שכבר מבוצעת או עיכוב של פונקציה שכבר מדוכאת. התייחסות הפוכה חיובית היא נדירה, מכיוון שהיא מביאה את המערכת הביולוגית למצב לא יציב.
קשתות רפלקס פשוטות (מונו-סינפטיות) מורכבות משני נוירונים בלבד (אפרנטי ו-efferent) ונבדלות רק ברפלקסים פרופריוצפטיביים. הקשתות הנותרות כוללות את כל הרכיבים לעיל.
קשת רפלקס היא שרשרת של נוירונים מקולטן היקפי דרך מערכת העצבים המרכזית לאפקטור היקפי. האלמנטים של קשת הרפלקס הם קולטן היקפי, מסלול אפרנטי, אינטרנורון אחד או יותר, מסלול efferent ואפקטור.
כל הקולטנים מעורבים ברפלקסים מסוימים, כך שהסיבים האפרנטיים שלהם משמשים כנתיב האפרנטי של קשת הרפלקס המקבילה. מספר האינטרנוירונים תמיד גדול מאחד, למעט רפלקס המתיחה המונוסינפטי. המסלול הפושט מיוצג על ידי אקסונים מוטוריים או סיבים פוסט-גנגליוניים של מערכת העצבים האוטונומית, והמשפיעים הם שרירי השלד והשרירים החלקים, הלב והבלוטות.
הזמן מתחילת הגירוי ועד לתגובת האפקטור נקרא זמן הרפלקס. ברוב המקרים הוא נקבע בעיקר על פי זמן ההולכה במסלולים האפרנטיים והעפרנטיים ובחלק המרכזי של קשת הרפלקס, אליו יש להוסיף את זמן הטרנספורמציה של הגירוי בקולטן לדחף מתפשט, הזמן של העברה דרך סינפסות במערכת העצבים המרכזית (עיכוב סינפטי), זמן ההעברה ממסלול efferent לזמן הפעלת האפקטור והאפקטור.
קשתות רפלקס מחולקות למספר סוגים
1. קשתות רפלקס מונו-סינפטיות – רק סינפסה אחת, הממוקמת במערכת העצבים המרכזית, משתתפת בקשת כזו. רפלקסים כאלה נפוצים מאוד בכל בעלי החולייתנים, הם מעורבים בוויסות טונוס שריריםויציבה (למשל טלטלה בברך). בקשתות אלו נוירונים אינם מגיעים למוח, ופעולות רפלקס מבוצעות ללא השתתפותו, שכן הם סטריאוטיפים ואינם דורשים מחשבה או החלטה מודעת. הם חסכוניים מבחינת מספר הנוירונים המרכזיים המעורבים ומוותרים על התערבות המוח.
2. קשתות רפלקס עמוד השדרה פוליסינפטיות - הן מערבות לפחות שתי סינפסות הממוקמות במערכת העצבים המרכזית, שכן נוירון שלישי כלול בקשת - נוירון בין-קלורית, או נוירון ביניים. כאן יש סינפסות בין הנוירון הסנסורי לאינטרנורון ובין הנוירונים הבין-קלוריים והמוטוריים. קשתות רפלקס כאלה מאפשרות לגוף לבצע תגובות לא רצוניות אוטומטיות הנחוצות להסתגלות לשינויים בסביבה החיצונית (לדוגמה, רפלקס אישוניםאו שמירה על שיווי משקל בזמן תנועה) ולשינויים בגוף עצמו (ויסות קצב הנשימה, לחץ דם וכו').
3. קשתות רפלקס פוליסינפטיות המערבות גם את חוט השדרה וגם את המוח - בסוג זה של קשתות רפלקס ישנה סינפסה בחוט השדרה בין הנוירון הסנסורי לנוירון השולחת דחפים למוח.
ניתן לסווג רפלקסים לפי קריטריונים שונים. אז, בהתאם לרמת סגירת הקשת, כלומר. לפי מיקום מרכז הרפלקס, הרפלקסים מחולקים לשדרה (הרפלקס נסגר בחוט השדרה), בולברי (מרכז הרפלקס הוא medulla oblongata), mesencephalic (קשת הרפלקס סגורה במוח האמצעי), דיאנצפלית ו מרכזי רפלקס קליפת המוח ממוקמים ב-telencephalon ובקורטקס של המיספרות גדולות, בהתאמה.
על פי תכונת האפקטור, הם סומטיים, כאשר הנתיב האפרנטי של הרפלקס מספק עיוור מוטורי של שרירי השלד, וצמחיים, כאשר האיברים הפנימיים הם המשפיעים.
בהתאם לסוג הקולטנים המגורים, הרפלקסים מחולקים ל-exteroceptive (אם הקולטן קולט מידע מהסביבה החיצונית), פרופריוצפטיבי (קשת הרפלקס מתחילה מהקולטנים של מנגנון השריר והשלד) ואינטרוצפטיב (מהקולטנים של איברים פנימיים).
רפלקסים אינטרוספטיביים, בתורם, מחולקים לקרביים-קרביים (קשת רפלקס מחברת שני איברים פנימיים), קרביים-שריריים (קולטנים ממוקמים על מנגנון השריר-גיד, האפקטור הוא איבר פנימי) וקרביים-עוריים (קולטנים הם מקומי בעור, איברים עובדים - קרניים).
לפי פבלוב, הרפלקסים מחולקים למותנים (מפותחים במהלך החיים, ספציפיים לכל פרט) ולא מותנים (מולדים, ספציפיים למין: מזון, מיני, הגנתי-מוטורי, הומאוסטטי וכו').
ללא קשר לסוג הרפלקס, קשת הרפלקס שלו מכילה קולטן, מסלול אפרנטי, מרכז עצבים, מסלול efferent, איבר עובד ומשוב. היוצא מן הכלל הוא רפלקסי האקסון, שקשת הרפלקס שלהם ממוקמת בתוך נוירון אחד: תהליכים תחושתיים יוצרים דחפים צנטריפטליים, העוברים דרך גוף הנוירון, מתפשטים לאורך האקסון למערכת העצבים המרכזית, ולאורך ענף האקסון. , הדחפים מגיעים לאפקטור. רפלקסים כאלה מיוחסים לתפקוד מערכת העצבים המטא-סימפתטית; באמצעותם, למשל, מתבצעים מנגנונים לוויסות טונוס כלי הדם ופעילות בלוטות העור.
הפונקציה של תפיסת גירוי והמרתו לאנרגיית עירור מתבצעת על ידי קולטנים של קשתות רפלקס. לאנרגיית הקולטן של עירור יש אופי של תגובה מקומית, שחשובה בהדרגה של עירור לפי כוח.
בהתבסס על המבנה והמקור של הקולטנים, ניתן לחלק אותם לקצות עצבים תחושתיים ראשוניים, תחושתיים משניים וקצות עצבים חופשיים. בראשון, הנוירון עצמו פועל כקולטן (הוא מתפתח מהנוירופיתל); אין מבנים מתווכים בין הגירוי לנוירון האפרנטי הראשון. התגובה המקומית של הקולטנים התחושתיים הראשוניים - פוטנציאל הקולטן - היא גם פוטנציאל מחולל, כלומר. גרימת פוטנציאל פעולה על פני הממברנה של הסיב האפרנטי. קולטנים תחושתיים ראשוניים כוללים קולטנים חזותיים, חוש הריח, כימו- ובארורצפטורים של מערכת הלב וכלי הדם.
תאי חישה משניים הם מבנים מיוחדים ממקור לא עצבי המקיימים אינטראקציה עם הדנדריטים של תאי חישה פסאודו-חד-קוטביים בעזרת מגעים של קולטן עצבי סינפטי. פוטנציאל הקולטן הנובע מפעולת גירוי בתאים בעלי חישה משנית אינו מחולל ואינו גורם להופעת פוטנציאל פעולה על גבי הממברנה של הסיב האפרנטי. הפוטנציאל הפוסט-סינפטי המעורר נוצר רק באמצעות מנגנון השחרור של המתווך על ידי תא הקולט. הדרגת עוצמת הגירוי מתבצעת באמצעות הפרשה של כמויות שונות של המתווך (ככל שהמתווך משתחרר, כך הגירוי חזק יותר).
תאי חישה משניים כוללים קולטנים שמיעתיים, וסטיבולריים, קרוטידים, מישוש ואחרים. לפעמים, בגלל המוזרויות של התפקוד, קבוצה זו כוללת קולטני פוטו, שמבחינה אנטומית ובשל מוצאם מהנוירו-אפיתל, הם בעלי חישה משנית.
קצות עצבים חופשיים הם דנדריטים מסועפים של תאי חישה פסאודו-חד-קוטביים וממוקמים כמעט בכל הרקמות של גוף האדם.
בהתאם לאופי האנרגיה של הגירוי שאליו מגיב הקולטן, הם מחולקים למכנורצפטורים (מגע, ברוררצפטורים, קולטני נפח, שמיעתי, וסטיבולרי; הם, ככלל, תופסים גירוי מכני בעזרת יציאות תאים), כימורצפטורים (חוש הריח; ), קולטנים כימיים של כלי דם, מערכת העצבים המרכזית, קולטני פוטו (קולטים גירוי דרך יציאות בצורת מוט וחרוט של התא), קולטנים תרמיים (מגיבים לשינוי "חם-קר" - גופי Rufini וצלוחיות קראוזה של ממברנות ריריות) ונוציצפטורים (קצוות כאב שאינם מובלעים).
היווצרות פוסט-קולטן של קשתות רפלקס הוא מסלול אפרנטי שנוצר על ידי נוירון סנסורי פסאודו-חד-קוטבי, שגופו שוכן בגנגליון השדרה, והאקסונים יוצרים את השורשים האחוריים של חוט השדרה. תפקידו של המסלול האפרנטי הוא להעביר מידע אל הקישור המרכזי, יתרה מכך, בשלב זה המידע מקודד. למטרות אלו, בגוף של בעלי חוליות, נעשה שימוש בקוד בינארי המורכב מתפרצים (מטחים) של דחפים ומרווחים ביניהם. ישנם שני סוגים עיקריים של קידוד: תדר ומרחב.
הראשון הוא היווצרות של מספר שונה של דחפים בפרץ, מספר שונה של פרצים, משך הזמן שלהם ומשך ההפסקות ביניהם, בהתאם לעוצמת הגירוי המופעל על הקולטן. קידוד מרחבי מבצע את הדרגת הכוח של הגירוי, הכולל מספר שונה של סיבי עצב, שלאורכם מתבצעת עירור בו זמנית.
הרכב המסלול האפרנטי כולל בעיקר סיבי A-b, A-c ו-A-d.
לאחר שעבר דרך הסיבים, הדחף העצבי נכנס למרכז הרפלקס, שבמובן האנטומי הוא אוסף של נוירונים הממוקמים ברמה מסוימת של מערכת העצבים המרכזית ולוקחים חלק ביצירת רפלקס זה. תפקידו של מרכז הרפלקס הוא לנתח ולסנתז מידע, כמו גם להעביר מידע מהנתיב האפרנטי לנתיב האפרנטי.
בהתאם למחלקה של מערכת העצבים (סומטית ואוטונומית), רפלקסים, שמרכזם ממוקם בחוט השדרה, נבדלים בלוקליזציה של נוירונים בין-קליריים. אז, עבור מערכת העצבים הסומטית, מרכז הרפלקס ממוקם באזור הביניים בין הקרניים הקדמיות והאחוריות של חוט השדרה. מרכז הרפלקס של מערכת העצבים האוטונומית (הגופים של נוירונים בין קלוריות) נמצא בקרניים האחוריות. החלקים הסומטיים והאוטונומיים של מערכת העצבים שונים גם בלוקליזציה של נוירונים efferent. גופי הנוירונים המוטוריים של מערכת העצבים הסומטית שוכנים בקרניים הקדמיות של חוט השדרה, גופם של הנוירונים הפרה-גנגליונים של המערכת האוטונומית שוכבים ברמת הקרניים האמצעיות.
האקסונים של שני סוגי התאים יוצרים את הנתיב האפרנטי של קשת הרפלקס. במערכת העצבים הסומטית היא רציפה, היא מורכבת מסיבים סוג א-ב. היוצאים מן הכלל היחידים הם סיבי A-g, המוליכים עירור מתאי חוט השדרה לסיבים תוך-פיוסאליים של צירי השריר. הנתיב האפרנטי של מערכת העצבים האוטונומית מופרע בגנגליון האוטונומי, הממוקם או תוך-מוורי (חלק פאראסימפתטי) או ליד חוט השדרה (בנפרד או בגזע הסימפתטי - חלק סימפטי). הסיב הפרה-גנגליוני שייך לסיבי B, הסיב הפוסט-גנגליוני שייך לקבוצת C.
האיבר העובד לחלק הסומטי של מערכת העצבים הוא שריר שלד מפוספס, בקשת הווגטטיבית האפקטור הוא בלוטה או שריר (לב חלק או מפוספס). בין המסלול היוצא לאיבר העובד יש סינפסה מיונאורלית או נוירו-הפרשתית כימית.
קשת הרפלקס נסגרת בטבעת עקב השפעה הפוכה - זרימת הדחפים מקולטני האפקטור חזרה למרכז הרפלקס. תפקוד משוב - איתות למערכת העצבים המרכזית על הפעולה שבוצעה. אם זה לא מבוצע מספיק, מרכז העצבים נרגש - הרפלקס ממשיך. כמו כן, עקב ההשפעה ההפוכה, מתבצעת השליטה בפעילות ההיקפית של מערכת העצבים המרכזית.
הבחנה בין משוב שלילי לחיובי. הראשון, בעת ביצוע פונקציה מסוימת, משיק מנגנון המעכב פונקציה זו. משוב חיובי מורכב מגירוי נוסף של פונקציה שכבר מבוצעת או עיכוב של פונקציה שכבר מדוכאת. התייחסות הפוכה חיובית היא נדירה, מכיוון שהיא מביאה את המערכת הביולוגית למצב לא יציב.
קשתות רפלקס פשוטות (מונו-סינפטיות) מורכבות משני נוירונים בלבד (אפרנטי ו-efferent) ונבדלות רק ברפלקסים פרופריוצפטיביים. הקשתות הנותרות כוללות את כל הרכיבים לעיל.
תכונות פיזיולוגיות ומשמעות תפקודית של סיבי עצב
לסיבי עצב יש את העצירות הגבוהה ביותר, את קצב ההולכה הגבוה ביותר של עירור, את תקופת העקשנות הקצרה ביותר ואלישות גבוהה. זה מסופק רמה גבוההתהליכים מטבוליים ופוטנציאל ממברנה נמוך.
תפקיד: הולכת דחפים עצביים מקולטנים למערכת העצבים המרכזית ולהיפך.
תכונות מבניות וסוגים של סיבי עצב
סיב העצב - האקסון - מכוסה בקרום תא.
ישנם 2 סוגים של סיבי עצב:
סיבי עצב ללא מיאלין - שכבה אחת של תאי שוואן, ביניהם - רווחים דמויי חריצים. קרום תאלאורך כל המגע עם סביבה. כאשר מוחל גירוי, מתרחשת עירור באתר הפעולה של הגירוי. לסיבי עצב ללא מיאלין יש תכונות אלקטרוגניות (היכולת ליצור דחפים עצביים) לאורך כל הדרך.
סיבי עצב בעלי מיאלין - מכוסים בשכבות של תאי שוואן, שבמקומות יוצרים צמתים של Ranvier (אזורים ללא מיאלין) כל 1 מ"מ. משך היירוט של Ranvier הוא 1 מיקרומטר. מעטפת המיאלין מבצעת פונקציות טרופיות ומבודדות (התנגדות גבוהה). לאזורים המכוסים במיאלין אין תכונות אלקטרוגניות. יש להם את היירוטים של Ranvier. עירור מתרחש ביירוט של Ranvier הקרוב ביותר לאתר הפעולה של הגירוי. ביירוטים של Ranvier צפיפות גבוההלכן, בכל יירוט של Ranvier, מתרחשת עלייה בדחפים עצביים.
יירוטים של Ranvier פועלים כמחזרים (יוצרים ומעצימים דחפים עצביים).
מנגנון ההולכה של עירור לאורך סיב העצב
1885 - L. German - זרמים מעגליים נוצרים בין החלקים הנרגשים והלא מתרגשים של סיב העצב.
תחת פעולתו של חומר גירוי, יש הבדל פוטנציאלי בין המשטח החיצוני והפנימי של הרקמה (אזורים הנושאים מטענים שונים). בין אזורים אלו נוצר זרם חשמלי (תנועת יוני Na +). בתוך סיב העצב, נוצר זרם מהקוטב החיובי אל הקוטב השלילי, כלומר, הזרם מכוון מהאזור הנרגש אל הלא נרגש. זרם זה יוצא דרך האזור הלא נרגש וגורם לו להיטען מחדש. על פני השטח החיצוניים של סיב העצב, הזרם זורם מהאזור הלא נרגש לאזור הנרגש. זרם זה אינו משנה את מצב האזור הנרגש, מכיוון שהוא במצב של עמידות.
עדות לנוכחות זרמים מעגליים: סיב העצב מונח פנימה תמיסת NaClולרשום את מהירות העירור. לאחר מכן מניחים את סיב העצב בשמן (ההתנגדות עולה) - מהירות ההולכה יורדת ב-30%. לאחר מכן, סיב העצב נשאר באוויר - קצב העירור מופחת ב-50%.
תכונות של הולכת עירור לאורך סיבי עצב עם מיאלין וסיבי עצב ללא מיאלין:
סיבי מיאלין - בעלי נדן בעל עמידות גבוהה, תכונות אלקטרוגניות רק בצמתים של Ranvier. תחת פעולת הגירוי, עירור מתרחש ביירט הקרוב ביותר של Ranvier. יירוט שכן במצב קיטוב. הזרם המתקבל גורם לדה-פולריזציה של המיירט הסמוך. לצמתים של Ranvier יש צפיפות גבוהה של תעלות Na, לכן, בכל צומת הבא, נוצר פוטנציאל פעולה מעט גדול יותר (במשרעת), בשל כך, העירור מתפשט ללא ירידה ויכול לקפוץ על מספר צמתים. זוהי התיאוריה המלוחה של טאסאקי. ההוכחה לתיאוריה היא שלסיב העצב הוזרקו תרופות שחוסמות מספר יירוטים, אך לאחר מכן נרשמה הולכה של עירור. זוהי שיטה אמינה ורווחית ביותר, שכן נזק קל מתבטל, מהירות העירור מוגברת ועלויות האנרגיה מופחתות;
סיבים שאינם מיאלינים - למשטח יש תכונות אלקטרוגניות לכל אורכו. לכן, זרמים מעגליים קטנים מתרחשים במרחק של כמה מיקרומטרים. לעירור יש צורה של גל שנוסע ללא הרף.
שיטה זו פחות משתלמת: עלויות אנרגיה גבוהות (להפעלת משאבת Na-K), קצב עירור נמוך יותר.
סיווג סיבי עצב
סיבי עצב מסווגים לפי:
משך פוטנציאל הפעולה;
מבנה (קוטר) של הסיב;
מהירות עירור.
נבדלות בין הקבוצות הבאות של סיבי עצב:
קבוצה A (אלפא, בטא, גמא, דלתא) - פוטנציאל הפעולה הקצר ביותר, מעטפת המיאלין העבה ביותר, קצב העירור הגבוה ביותר;
קבוצה B - מעטפת המיאלין פחות בולטת;
קבוצה C - ללא מעטפת מיאלין.
הבדלים מורפולוגיים בין דנדריטים ואקסונים
1. לנוירון בודד יש כמה דנדריטים, אקסון הוא תמיד אחד.
2. הדנדריטים תמיד קצרים יותר מהאקסון. אם גודל הדנדריטים אינו עולה על 1.5-2 מ"מ, אז האקסונים יכולים להגיע ל-1 מ' או יותר.
3. דנדריטים מתרחקים בצורה חלקה מגוף התא ובהדרגה יש להם קוטר קבוע על פני מרחק ניכר.
4. דנדריטים מסתעפים בדרך כלל מתחת זוית חדה, והענפים מכוונים הרחק מהתא. אקסונים נותנים בטחונות לרוב בזווית ישרה; הכיוון של בטחונות אינו קשור ישירות למיקום גוף התא.
5. תבנית ההסתעפות הדנדרטית בתאים מאותו סוג קבועה יותר מהסתעפות האקסון של תאים אלו.
6. הדנדריטים של נוירונים בוגרים מכוסים בקוצים דנדריטים, אשר נעדרים על הסומה ועל החלק הראשוני של הגזעים הדנדריטים. לאקסונים אין קוצים.
7. לדנדריטים לעולם אין מעטפת עיסתית. האקסונים מוקפים לרוב במיאלין.
8. לדנדריטים ארגון מרחבי סדיר יותר של מיקרוטובולים, האקסונים נשלטים על ידי נוירופילמנטים והמיקרוטובולים פחות מסודרים
9. בדנדריטים, בעיקר בחלקים הפרוקסימליים שלהם, יש רשת אנדופלזמה וריבוזומים, שאינם נמצאים באקסונים.
10. פני השטח של הדנדריטים נמצאים ברוב המקרים במגע עם פלאקים סינופטיים ויש להם אזורים פעילים עם התמחות פוסט-סינפטית.
מבנה הדנדריטים
אם יש ספרות גדולה יחסית על הגיאומטריה של הדנדריטים, אורך הענפים שלהם, כיוון, אז בערך מבנה פנימי, לגבי המבנה של המרכיבים הבודדים של הציטופלזמה שלהם, יש רק מידע מפוזר נפרד. מידע זה הפך לאפשרי רק עם הכנסתם של מחקרים מיקרוסקופיים אלקטרוניים לנוירוהיסטולוגיה.
המאפיינים העיקריים של הדנדריט, המבדילים אותו בחתכים מיקרוסקופיים אלקטרונים:
1) חוסר נדן מיאלין,
נוכחות המערכת הנכונה של מיקרוטובולים,
3) נוכחותם של אזורים פעילים של סינפסות עליהם עם צפיפות אלקטרונים מבוטאת בבירור של הציטופלזמה של הדנדריט,
4) יציאה מהגזע המשותף של הדנדריט של עמוד השדרה,
5) אזורים מאורגנים במיוחד של צמתים ענפים,
6) הכללת ריבוזומים,
7) נוכחות של רטיקולום אנדופלזמי גרגירי ולא גרגירי באזורים הפרוקסימליים.
התכונה הבולטת ביותר של הציטופלזמה הדנדרטית היא נוכחותם של מיקרוטובולים רבים. הם מזוהים היטב הן בחתכים רוחביים והן בחתכים אורכיים. החל מהקטע הפרוקסימלי של הדנדריט, מיקרוטובולים פועלים במקביל לציר הארוך של הדנדריט לענפים המרוחקים שלו. מיקרוטובולים עוקבים בדנדריט במקביל זה לזה, מבלי להתחבר או להצטלב זה עם זה. בחתכים, ניתן לראות שהמרחקים בין הצינוריות הבודדים קבועים. צינוריות דנדריטיות בודדות משתרעות על פני מרחקים ארוכים למדי, ולעיתים קרובות עוקבות אחרי עקומות שעשויות להיות לאורך מהלך הדנדריטים. מספר הצינוריות קבוע יחסית ליחידת שטח של חתך הדנדריט והוא בערך 100 ל-1 מיקרומטר. מספר זה אופייני לכל דנדריטים שנלקחו ממנו מחלקות שונותמערכת העצבים המרכזית והפריפריאלית, סוגים שוניםבעלי חיים.
תפקידם של המיקרוטובולים הוא הובלה של חומרים לאורך תהליכים של תאי עצב.
כאשר המיקרוטובולים מושמדים, ניתן לשבש את הובלת החומרים בדנדריט, וכך, מקטעי הקצה של התהליכים נמנעת הזרם של חומרים מזינים ו חומרי אנרגיהמגוף התא. דנדריטים, על מנת לשמור על מבנה המגעים הסינפטיים בתנאים קיצוניים ובכך להבטיח את תפקוד האינטראקציה הבין-עצבית, מפצים על הגירעון חומרים מזיניםבשל מבנים הסמוכים אליהם (פלאקים סינפטיים, מעטפת מיאלין רב-שכבתית של סיבים רכים, כמו גם שברי תאי גליה).
אם פעולת הגורם הפתוגני מבוטלת בזמן, הדנדריטים משחזרים את המבנה ואת הארגון המרחבי הנכון של המיקרוטובולים, ובכך משחזרים את מערכת הובלת החומר, הטבועה במוח הרגיל. אם החוזק ומשך הזמן של הגורם הפתוגני משמעותיים, אזי תופעות האנדוציטוזיס, במקום תפקודן ההסתגלותי, עלולות להפוך לקטלניות עבור דנדריטים, שכן לא ניתן לנצל שברים פגוציטים, והצטברות בציטופלזמה של דנדריטים, תוביל לבלתי הפיך שלו. נֵזֶק.
הפרה בארגון של microtubules מובילה לשינוי חד בהתנהגות של בעלי חיים. בבעלי חיים שבהם הושמדו מיקרוטובולים בדנדריטים בניסוי, נצפה חוסר ארגון של צורות התנהגות מורכבות, בעוד רפלקסים מותנים פשוטים נשמרו. בבני אדם, זה יכול להוביל להפרעות חמורות בפעילות עצבית גבוהה יותר.
העובדה שהדנדריטים הם המקום הרגיש ביותר לפעולת גורם פתולוגי ב מחלת נפש, מעידות כמה עבודות של מדענים אמריקאים. התברר שבדמנציה סנילי (דמנציה ציאנוטית) ובמחלת אלצהיימר, תכשירי מוח המעובדים בשיטת גולגי אינם חושפים את התהליכים של תאי עצב. גזעי הדנדריטים נראים שרופים ומפוחמים. אי זיהוי התהליכים הללו על הכנות היסטולוגיות של המוח קשור כנראה גם להפרה של מערכת המיקרוטובולים והנוירופילמנטים בתהליכים אלה.
נמצא בדנדריטים. הם עוקבים במקביל לציר הארוך של הדנדריט, הם יכולים לשכב בנפרד או להיאסף בצרורות, אבל הם לא ממוקמים בקפדנות בציטופלזמה. כנראה, יחד עם microtubules, הם יכולים להיות מקבילה של neurofibrils.
כל הדנדריטים של מערכת העצבים המרכזית מאופיינים בעלייה של פני השטח עקב חלוקה דיכוטומית מרובה. במקרה זה, אתרי הרחבה מיוחדים או צמתים סניפים נוצרים באזורי החלוקה.
ניתוח רגיל מראה שבצומת הענף, שאליו מתקרבים שני ענפים דנדריטים, כל אחד נושא את האות שלו, ניתן לבצע את הפעולות הבאות. דרך צומת הסניף תא מטען משותףובהמשך לגוף הנוירון עובר:
או אות מסניף אחד,
או סתם מאחר
או תוצאה של אינטראקציה של שני אותות,
או שהאותות מבטלים זה את זה.
הציטופלזמה של הצומת הסניף מכילה כמעט את כל המרכיבים האופייניים לגוף תא עצב, והאתרים שונים מאוד במבנה שלהם מהציטופלזמה של הגזע הדנדריטי המשותף והענפים המתקבלים בחלוקה. הצמתים הענפים מכילים מספר מוגבר של מיטוכונדריה, רשת גרעינית וחלקה, נראים אשכולות של ריבוזומים בודדים וריבוזומים המורכבים לשושנת. רכיבים אלה (רשת גרגירית וחלקה, ריבוזומים) מעורבים ישירות בסינתזת חלבון. הצטברות המיטוכונדריה במקומות אלו מעידה על עוצמת תהליכי החמצון.
פונקציות של דנדריטים
ברצוני לציין כי הקשיים העיקריים בהם נתקל חוקר בבחינת תפקוד הדנדריטים הוא היעדר מידע על תכונות קרום הדנדריט (בניגוד לממברנה של גוף הנוירון) עקב חוסר האפשרות להחדיר מיקרואלקטרודה לתוך הדנדריט.
בהערכת הגיאומטריה הכוללת של הדנדריטים, התפלגות הסינפסות והמבנה המיוחד של הציטופלזמה במקומות ההסתעפות הדנדרטית, אפשר לדבר על לוקוסים נוירון מיוחדים עם תפקוד משלהם. הדבר הפשוט ביותר שניתן לייחס לאתרים דנדריטים באתרי הסתעפות הוא פונקציה טרופית.
מהאמור לעיל עולה שהציטופלזמה של הדנדריטים מכילה הרבה אולטרה רכיבים מבנייםמסוגל לספק את הפונקציות החשובות שלהם. ישנם לוקוסים מסוימים בדנדריט, שבהם לעבודתו יש מאפיינים משלה.
המטרה העיקרית של הענפים הדנדריטים הרבים של תא עצב היא לספק חיבור הדדי עם נוירונים אחרים. בקליפת המוח של יונקים נתח גדולחיבורים אקסודנדריים נופלים על מגעים עם יציאות מיוחדות מיוחדות של דנדריטים - קוצים דנדריטים. קוצים דנדריטים הם מבחינה פילוגנטית התצורות הצעירות ביותר במערכת העצבים. באונטוגניה, הם מבשילים מאוחר יותר מאשר מבנים עצביים אחרים ומייצגים את המנגנון הפלסטי ביותר של תא העצב.
ככלל, לעמוד השדרה הדנדריטי יש צורה אופיינית בקליפת המוח של יונקים. (איור 2). מגזע הדנדריטי הראשי יוצא גבעול צר יחסית, המסתיים בהרחבה - הראש. סביר להניח שצורה זו של התוספת הדנדרטית (נוכחות ראש) קשורה, מצד אחד, לעלייה באזור המגע הסינפטי עם סיום האקסון, ומצד שני, היא משמשת ל להכיל אברונים מיוחדים בתוך עמוד השדרה, בפרט, את המנגנון הקוצני, הקיים רק בעמוד השדרה הדנדריטי של קליפת המוח של היונקים. בהקשר זה, אנלוגיה לצורת סוף האקסון הסינפטי, כאשר סיב קדם-טרמינלי דק יוצר שלוחה, נראית מתאימה. התרחבות זו (רובד סינפטי) יוצרת מגע נרחב עם המצע המועצב ומכילה בתוך קבוצה גדולה של רכיבים אולטרה-סטרוקטורליים (שלפוחיות סינפטיות, מיטוכונדריה, נוירופילמנטים, גרגירי גליקוגן).
קיימת השערה (שבמיוחד משותפת ופותחה על ידי חתן פרס נובל פ. קריק) לפיה הגיאומטריה של עמוד השדרה יכולה להשתנות בהתאם למצב התפקודי של המוח. במקרה זה, הצוואר הצר של עמוד השדרה יכול להתרחב, ועמוד השדרה עצמו משתטח, וכתוצאה מכך עלייה ביעילות של מגע האקסו-עמוד השדרה.
אם הצורה והגודל של קוצים דנדריטים בקליפת המוח של יונקים יכולים להשתנות במקצת, אז הקבוע ביותר בהם הוא נוכחות של מנגנון עמוד שדרה ספציפי. זהו קומפלקס של צינורות מחוברים (בור) הממוקמים, ככלל, בראש עמוד השדרה. ככל הנראה, אברון זה קשור לתפקודים חשובים מאוד הגלומים בתצורות המוח הצעירות ביותר מבחינה פילוגנטית, שכן המנגנון הקוצני נמצא בעיקר בקליפת המוח, ורק בבעלי חיים גבוהים יותר.
למרות הכל, עמוד השדרה הוא נגזרת של הדנדריט, הוא חסר נוירופילמנטים וצינוריות דנדריטים, הציטופלזמה שלו מכילה מטריצה גרגירית גסה או עדינה. עוד אחד תכונהעמוד השדרה בקליפת המוח היא הנוכחות החובה של מגעים סינפטיים עם קצוות אקסונים עליהם. לציטופלזמה של עמוד השדרה יש מרכיבים מיוחדים המבדילים בינה לבין גבעולים דנדריטים. אפשר לציין שלשה מוזרה בציטופלזמה של עמוד השדרה: התמחות תת-סינפטית של אזורים פעילים - מנגנון קוצני - מיטוכונדריה. בהתחשב במגוון התפקודים המורכבים והחשובים שמבצעים המיטוכונדריה, ניתן לצפות גם לביטויים תפקודיים מורכבים ב"שלישיות" במהלך העברה סינפטית. ניתן לומר שהציטופלזמה של עמוד השדרה הדנדריטי והמנגנון הקוצני עשויה להיות קשורה ישירות לתפקוד הסינפטי.
קוצים דנדריטים וקצוות דנדריטים גם רגישים מאוד לגורמים קיצוניים. בכל סוג של הרעלה (לדוגמה, מתכות אלכוהוליות, היפוקסיות, כבדות - עופרת, כספית וכו'), מספר הקוצים המצויים על הדנדריטים של תאי קליפת המוח משתנה. ככל הנראה, עמודי השדרה אינם נעלמים, אך המרכיבים הציטופלזמיים שלהם מופרעים, והם ספוגים גרוע יותר במלחים. מתכות כבדות. מאחר שעמוד השדרה הוא אחד המרכיבים המבניים של מגעים בין-עצביים, תקלות בהם מובילות לפגיעה חמורה בתפקוד המוח.
במקרים מסוימים, עם פעולה קצרת טווח של גורם קיצוני, במבט ראשון, עלול להתרחש מצב פרדורסלי, כאשר מספר הקוצים שנמצאים על הדנדריטים של תאי המוח אינו פוחת, אלא עולה. אז זה נצפה במהלך איסכמיה מוחית ניסיוני בתקופה הראשונית שלה. במקביל לעלייה במספר הקוצים המזוהים, ה מצב תפקודימוֹחַ. במקרה זה, היפוקסיה היא גורם התורם להגברת חילוף החומרים ברקמת העצבים, יישום טוב יותר של רזרבות שאינן בשימוש ב סביבה רגילה, והבעירה המהירה של רעלים שהצטברו בגוף. מבחינה אולטרה-סטרוקטורלית, זה מתבטא במחקר אינטנסיבי יותר של הציטופלזמה של עמוד השדרה, צמיחה והגדלה של בורות המים של מנגנון עמוד השדרה. כנראה, תופעה זו של ההשפעה החיובית של היפוקסיה נצפית כאשר אדם, חווה גדול אימון גופניבתנאים של היפוקסיה, כובש פסגות הרים. קשיים אלה מתוגמלים על ידי עבודה פרודוקטיבית יותר, הן של המוח והן של איברים אחרים.
היווצרות דנדריטים
דנדריטים והקשרים הפנימיים שלהם נוצרים במהלך ההתפתחות האונטוגנטית של המוח. יתר על כן, הדנדריטים, במיוחד אלו האפיקיים, אצל אנשים צעירים נשארים חופשיים במשך זמן מה ליצור קשרים חדשים. חלקי הדנדריט הממוקמים קרוב יותר לגוף התא קשורים כנראה לרפלקסים מותנים טבעיים חזקים ופשוטים יותר, והקצוות נותרים ליצירת קשרים ואסוציאציות חדשות.
בבגרות כבר אין אזורים נקיים ממגע פנימי על הדנדריטים, אבל עם ההזדקנות, קצוות הדנדריטים הם שסובלים לראשונה ומבחינת רוויה במגעים.
אצל אנשים זקנים הם דומים לדנדריטים יַלדוּת. הדבר מתרחש הן עקב היחלשות תהליכי סינתזת חלבון התחבורה בתא, והן עקב פגיעה באספקת הדם למוח. אולי כאן טמון הבסיס המורפולוגי לדבר כזה ידוע בנוירולוגיה ובפנים חיי היום - יוםהעובדה שכאשר אנשים זקנים מתקשים ללמוד משהו חדש, הם לעתים קרובות שוכחים אירועים עכשוויים וזוכרים היטב את העבר. אותו הדבר נצפה במקרה של הרעלה.
כפי שכבר צוין, הגידול והסיבוך של העץ הדנדריטי בפילוגניה נחוצים לא רק לתפיסה מספר גדולפולסים נכנסים, אבל גם לעיבוד מקדים.
לדנדריטים של הנוירונים של מערכת העצבים המרכזית יש תפקוד סינפטי לכל אורכו, והקטעים הסופיים אינם נחותים בשום אופן מהאמצעיים בכך. אם אנחנו מדבריםעל החלקים הדיסטאליים (הסופניים) של הדנדריטים האפיקליים נוירונים פירמידלייםקליפת המוח, אז חלקם ביישום של אינטראקציות בין-נוירונאליות משמעותי אפילו יותר מאלה הפרוקסימליות. לשם יותרלוחות סינפטיים סופניים על תא המטען עצמו ועל ענפי הדנדריט האפיקלי מצטרפים למגעים נוספים על קוצים דנדריטים.
במחקר של בעיה זו באמצעות מיקרוסקופ אלקטרונים, החוקרים השתכנעו גם שחלקי הקצה של הדנדריטים מכוסים בצפיפות בפלאקים סינפטיים, ולכן מעורבים ישירות באינטראקציות בין-עצביות. אלקטרון מיקרוסקופיהראה גם שדנדריטים יכולים ליצור קשרים זה עם זה. מגעים אלה יכולים להיות מקבילים, להם רוב המחברים מייחסים תכונות אלקטרוטוניות, או סינפסות א-סימטריות טיפוסיות עם אברונים מוגדרים היטב המספקים העברה כימית. מגעים דנדרו-דנדריטים כאלה רק מתחילים למשוך את תשומת הלב של החוקרים. אז, הדנדריט לכל אורכו מבצע פונקציה סינפטית. כיצד מותאם פני השטח של הדנדריט לספק מגע עם קצוות אקסונים?
קרום פני השטח של הדנדריט מיועד לשמש למקסימום עבור מגע פנימי. הדנדריט כולו מכוסה שקעים, קפלים, כיסים, יש לו אי-סדירות מסוגים של מיקרו-צמיחת, קוצים, תוספות דמוי פטריות וכו'. כל התבליטים הללו של גזעים דנדריטים תואמים את הצורה והגודל של הקצוות הסינפטיים הנכנסים. ובתוך מחלקות שונותמערכת העצבים ובבעלי חיים שונים להקלה על פני השטח הדנדריטיים יש תכונות ספציפיות. כמובן, התוצאה המדהימה ביותר של הממברנה הדנדרטית היא עמוד השדרה הדנדריטי.
דנדריטים רגישים מאוד לפעולתם של גורמים קיצוניים שונים. הפרות בהן מובילות למחלות רבות, כמו הפרעות נפשיות.
קשת הרפלקס מורכבת מ:
- קולטנים - תפיסת גירוי.
- סיב עצב רגיש (צנטריפטלי, אפרנטי) המעביר עירור למרכז
- מרכז העצבים, שבו מתרחש המעבר של עירור מנוירונים תחושתיים לנוירונים מוטוריים
- סיב עצב מוטורי (צנטריפוגלי, efferent) הנושא עירור ממערכת העצבים המרכזית לאיבר הפועל
- אפקטור - איבר עובד המבצע השפעה, תגובה בתגובה לגירוי הקולטן.
קולטנים ושדות קליטה
קוֹלֵט- תא תופס גירוי.
שדה קליטה- זהו האזור האנטומי, כאשר מגורה, הרפלקס הזה נגרם.
שדות הקליטה של קולטני חושים ראשוניים מאורגנים בצורה הפשוטה ביותר. לדוגמה, השדה הקליטה המישוש או הנוציספטיבי של פני העור הוא הסתעפות של סיב חושי בודד.
לקולטנים הממוקמים בחלקים שונים של השדה הקולט יש רגישות שונה לגירוי נאות. אזור רגיש מאוד ממוקם בדרך כלל במרכז השדה הקולט, והרגישות יורדת קרוב יותר לפריפריה של השדה הקולט.
השדות הקליטה של קולטני חישה משניים מאורגנים בצורה דומה. ההבדל הוא שענפי הסיב האפרנטי אינם מסתיימים בחופשיות, אלא יש להם מגע סינפטי עם תאי קולטן רגישים. שדות קולטניים, וסטיבולריים, אקוסטיים מאורגנים בצורה זו.
שדות קליטה חופפים. אותו אזור של המשטח הרגיש (לדוגמה, עור או רשתית) מועצב על ידי מספר רגישים סיבי עצב, אשר, עם הסתעפויותיהם, חופפים את השדות הקליטה של עצבים אפרנטיים בודדים.
בשל החפיפה של שדות קליטה, משטח התחושה הכולל של הגוף גדל.
סיווג רפלקסים.
לפי סוג חינוך:
מותנה (נרכש) - הגיבו לשם, רוק מהכלב אל האור.
ללא תנאי (מולדת) - בליעה ממצמצת, ברך.
לפי מיקום קולטנים:
אקסטרוספטיבי (עור, חזותי, שמיעתי, ריח)
interoceptive (מקולטנים של איברים פנימיים)
פרופריוצפטיבי (מקולטנים בשרירים, גידים, מפרקים)
לאפקטורים:
סומטי, או מוטורי, (רפלקסים שריר השלד);
איברים פנימיים וגטטיביים - עיכול, לב וכלי דם, הפרשה, הפרשה וכו'.
לפי מקור ביולוגי:
הגנתי, או מגן (תגובה לחלוקת כאב מישוש)
מערכת העיכול (קולטנים מגרים בחלל הפה.)
מיני (הורמונים בדם)
משוער (סיבוב הראש, הגוף)
מָנוֹעַ
פוזוטוניק (תומכת תנוחות גוף)
לפי מספר הסינפסות:
מונוסינפטי, שהקשתות שלו מורכבות מנוירונים אפרנטיים ויוצאים (לדוגמה, ברך).
Polysynaptic, שהקשתות שלו מכילות גם 1 או יותר נוירוני ביניים ויש להם 2 או יותר מתגים סינפטיים. (התייחסויות סומטיות וצמחיות).
דיסינפטי (2 סינפסות, 3 נוירונים).
לפי אופי התגובה:
מוטורי \ מוטורי (התכווצויות שרירים)
הפרשה (הפרשת בלוטת הפרשה)
Vasomotor (התרחבות והיצרות של כלי דם)
לב (שינוי. עבודת שריר הלב.)
לפי משך הזמן:
נסיגת יד פאזית (מהירה).
תחזוקת יציבה טוניקית (איטית).
לפי מיקום מרכז העצבים:
עמוד השדרה (נוירונים SM מעורבים) - משיכת היד הרחק מהמקטעים החמים 2-4, טלטלה בברך.
רפלקסים במוח
Bulbar (medulla oblongata) - סגירת העפעפיים בעת מגע. לקרנית.
Mesencephalic (מ' באמצע) - ציון דרך חזון.
Diencephalic (המוח האמצעי) - חוש ריח
קורטיקל (נבוח BP GM) - מותנה. ref.
נכסים מרכזי עצבים.
1. התפשטות חד צדדית של עירור.
עירור מועבר מהנוירון האפרנטי ל-efferent (סיבה: מבנה הסינפסה).
האטת העברת עירור.
מְרַכֵּך שֵׂעָר נוכחותן של סינפסות רבות תלויה גם בעוצמת החומר הגירוי (סיכום) ובמצב הפיזי. CNS (עייפות).
3.סיכוםסיכום השפעות, מתחת לגירויים בסף.
זמני: ר'. מהקודם Imp-sa עדיין לא עבר, אבל עקבות. כבר הגיע.
מרחבי: ערבוב כמה. מים גבאים הם מותנים ינשוף. תמונות. רפ.
מרכז הנחייה וחסימה.
מרכז ההקלה - מתרחש תחת פעולת הגירוי האופטימלי (תגובה מקסימלית) - הופיע. מרכז סיוע.
תחת הפעולה של min irr. (מופחתת אוט"ב רקציה) הייתה חסימה.
הטמעה וטרנספורמציה של קצב ההתרגשות.
טרנספורמציה - שינוי בתדירות של דחף עצבי בעת מעבר דרך מרכז העצבים. ניתן להגדיל או להקטין את התדירות.
הטמעה (ריקוד, שגרת יומיום)
תוֹצָאָה
העיכוב בסיום התגובה לאחר הפסקת הגירוי. קשור לעצב הדם. שֵׁד. סָגוּר מעגלים של נוירונים.
טווח קצר (שברירי שנייה)
ארוך (שניות)
פעילות קצבית של מרכזי עצבים.
עלייה או ירידה בתדירות של דחפים עצביים הקשורים לתכונות הסינפסה ולמשך האינטגרטיבי של נוירונים.
8. פלסטיות של מרכזי עצבים.
היכולת לבנות מחדש את הפונקציונליות של נכס לצורך ויסות יעיל יותר של פונקציות, יישום רפלקסים חדשים שלא היו אופייניים קודם לכן למרכז זה, או שחזור הפונקציות. בלב שכבת הסינפים נמצא שינוי בקניון-th str-ry.
שינויים בריגוש בהשפעת כימיקלים.
רגישות גבוהה להבדלים אמיתיים.
עייפות של מרכזי העצבים.
קשור לעייפות סינפסה גבוהה. הפחת רגשות. קולטנים.
עקרונות כלליים של פעילות הקואורדינציה של מערכת העצבים המרכזית.
בְּלִימָה-נר מיוחד. אָחוּז מתבטא בהפחתה או היעלמות מוחלטת של resp. תגובות.
עקרון ההתכנסות
התכנסות היא התכנסות של דחפים המגיעים דרך מסלולים אפרנטיים שונים בכל נוירון או מרכז עצב מרכזי אחד.
2 . עקרון ההתכנסות קשור קשר הדוק לעיקרון נתיב סופי משותףלפתוח את שרנקטון. גירויים רבים ושונים יכולים לעורר את אותו נוירון מוטורי ואת אותה תגובה מוטורית. עיקרון זה נובע מהמספר הלא שווה של מסלולים אפרנטיים ועפרנטיים.
עקרון ההתבדלות
זהו המגע של נוירון אחד עם רבים אחרים.
הקרנה וריכוז של עירור.
התפשטות תהליך העירור למרכזי עצבים אחרים נקראת הקרנה (אלקטורלי- בכיוון אחד , מוכלל- נרחב).
לאחר זמן מה, ההקרנה מוחלפת בתופעה של ריכוז עירור באותה נקודה ראשונית של מערכת העצבים המרכזית.
תהליך ההקרנה ממלא תפקידים חיוביים (יצירת רפלקסים מותנים חדשים) ושליליים (הפרה של היחסים העדינים שהתפתחו בין תהליכי העירור והעכבה, המובילים להפרעה בפעילות המוטורית).
עקרון ההדדיות (מאטה)
עירור של תאים מסוימים גורם לעיכוב של אחרים דרך הנוירון הבין-קלורי.
עקרון דומיננטי
אוכטומסקי ניסח את עקרון הדומיננטיות כעקרון עבודה של פעילות מרכזי העצבים. טווח דוֹמִינָנטִימציין את המוקד הדומיננטי של עירור של מערכת העצבים המרכזית, אשר קובע את הפעילות הנוכחית של הגוף.
עקרונות המיקוד הדומיננטי :
ריגוש מוגברת של מרכזי עצבים;
התמדה של עירור של עירור לאורך זמן;
היכולת לסכם גירויים זרים;
אינרציה (היכולת לשמור על עוררות במשך זמן רב לאחר סיום פעולת הגירוי); היכולת לגרום לעכבות מצומדות.

